
大草蛉对不同时期芳香植物甜罗勒及其主要挥发物质的嗅觉行为反应
2023-10-19 13:10杨亚洁岳艳丽顾俊杰
环境昆虫学报 2023年4期
杨亚洁,李 姝,方 艳,王 甦,岳艳丽,顾俊杰*
(1. 四川农业大学农学院,成都 611130;2. 北京市农林科学院植物保护研究所,北京 100097)
捕食性天敌昆虫在控制农林重要害虫、保证农产品的质量与产量等方面起着不可或缺的作用(陈学新等, 2010; 张帆等, 2015; 段雪莹等, 2021)。作为我国广泛分布的重要捕食性天敌,大草蛉Chrysopapallens具有捕食量大、适应性强、年发生世代数多、成虫寿命长且产卵量大等特点(杨集昆, 1974; 杨星科, 1998; Booetal., 1998)。近年来,越来越多的学者聚焦在大草蛉生物学研究,尤其是通过研究感觉器官、滞育、人工饲料、控害能力等挖掘大草蛉的生防价值(李姝等, 2019)。但由于大草蛉成虫搜寻扩散能力强而定殖性差,难以使其种群维持发挥持续性控害作用(武鸿鹄等, 2014),这极大限制了大草蛉的田间应用。
新兴的保护型生物防治策略是通过功能植物创造有利于天敌昆虫的生存环境,提高其适合度以增强害虫防治效果(戈峰, 2020)。其中,芳香植物是一类目前广泛应用的功能植物(宋备舟等, 2010; 张正群等, 2016)。很多研究表明芳香植物释放的挥发性气味对植食性昆虫具有杀虫、驱避和拒食作用(沈君辉等, 2007; Contietal., 2010; Dardourietal., 2018)。同时,田间实践也发现,通过间作芳香植物能调控害虫和天敌的种群数量,如在果园内种植芳香植物对天敌种群具有吸引作用,显著增加了主栽作物区的天敌昆虫多样性,提高了对目标害虫的生物防治效果(Songetal., 2017; Lietal., 2019; Hattetal., 2019)。然而,许多芳香植物对天敌昆虫作用的化学生态学机制并不明确。植物与昆虫的化学通讯系统中,植物释放的特定挥发性物质能够诱导植食性昆虫定位寄主、选择适宜的产卵场所或对植物产生逃避行为,也可为天敌定位寄主栖境提供线索(王冰等, 2021)。因此,天敌对芳香植物的嗅觉行为反应是揭示其化学生态学机制的重要基础。
甜罗勒Ocimumbasilicum为唇形科罗勒属喜温的一年生草本芳香植物,原产亚热带地区,主产于法国、美国南部、匈牙利、印尼、西班牙等地(黄士诚等, 1989),作为食品、化妆品等经济价值高(Mouraetal., 2020)。目前已有研究证明罗勒可直接影响害虫种群增长,例如茶园中间作丁香罗勒Ocimumgratissimum可以降低主要害虫茶尺蠖Ectropisobliqua种群(张正群等, 2016);蚕豆田Viciafaba间作丁香罗勒或者香薄荷Saturejahortensis可以降低黑豆蚜Aphisfabae种群数量(Basedowetal., 2006);圣罗勒Ocimumsanctum能降低绿豆象Callosobruchuschinensis的产卵量(Kiradoo &Srivastava, 2010)。本研究团队前期调查芳香植物上节肢动物群落发现,甜罗勒上大草蛉种群数量显著高于其他植物,那为什么大草蛉更喜趋向甜罗勒植株?是否与甜罗勒植株的挥发性化学物质有关?
为明确芳香植物甜罗勒对大草蛉的吸引作用机制,本研究(1)使用Y型嗅觉仪测定了大草蛉成虫对不同时期甜罗勒植株的嗅觉行为反应;(2)采用顶空固相微萃取方法收集并用气相色谱质谱分析了甜罗勒植株营养期的叶、开花期的叶和花的挥发性物质成分;(3)测定了大草蛉成虫对不同浓度下主要挥发物质标准品的嗅觉行为反应。本研究结果为探究大草蛉对芳香植物识别机理和反应机制提供基础行为学证据,亦可为进一步开发甜罗勒作为天敌的吸引定殖功能植物提供支撑数据。
1 材料与方法
1.1 材料
供试虫源:大草蛉来自北京市农林科学院植物保护研究所室内繁育种群。在盆栽蚕豆苗上扩繁豌豆修尾蚜种群,待扩繁到适宜的密度时将大草蛉放入养虫笼(50 cm×45 cm×45 cm,尼龙网120目)中进行试验种群扩繁。所有种群在恒定条件下(温度25℃±1℃,相对湿度70%±5%,光周期L∶ D=16 h∶8 h)饲养。
供试植物:甜罗勒种子购自于北京花儿朵朵仙子农业有限公司。待种子萌发长至3~4片真叶时,移栽到塑料花盆中(高为20 cm,直径为13 cm),盆栽所用基质成分为蛭石、珍珠岩、泥炭土按1∶1∶4比例混合,每盆移栽一株幼苗,底部吸水至土壤微湿,培养环境条件同上。待植株长至30 cm时开始试验。
供试试剂:芳樟醇、罗勒烯、萜品油烯、丁香酚标准品购于Sigma-Aldrich公司,樟脑、β-榄香烯购于上海源叶生物有限公司,纯度均在95%以上。以上6种标准品分别以石蜡油稀释至0.1 μg/L、0.01 μg/L,充分混匀后密封,放置于4℃冰箱内储存备用。
供试仪器:顶空固相微萃取-气相色谱-质谱联用仪(HS-SPME-GC-MS)(Agilent 6890N-5975)。“Y”型嗅觉仪的主臂内径为3 cm,长为15 cm,两个侧臂长为15 cm,夹角为60°,用Teflon管分别连接芳香植物植株和对照(洁净空气或石蜡油)后再连接LS-2800型气泵(北京劳动仪器研究所)。气泵推动空气进入系统,气流经活性炭、蒸馏水净化加湿、气味源后通过玻璃转子流量计进入Y形管两侧。
1.2 大草蛉对不同时期甜罗勒植株的嗅觉行为反应
Y型嗅觉仪放置在100 cm×100 cm×60 cm的试验箱中(25℃±1℃,RH 60%±5%)。用照度计测试两臂光照强度,并调整位置,保证“Y”型嗅觉仪各部位受光一致。接通抽气泵电源开始抽气,调节流速计使两边的气流速度相同,控制气体流速在400 mL/min左右。试验前先行通气10 min,使气味充满管道,以保证测试结果。试验前将5~7日龄的大草蛉成虫饥饿12 h。试验时,将饥饿后的大草蛉成虫逐头引入嗅觉仪的主臂内,观察大草蛉在5 min内的行为反应,大草蛉爬行超过某气味源试管臂的1/2处记为选择,在5 min内仍未到达某气味源试管臂1/2处,记为无选择。每测试5头大草蛉结束后,用95%乙醇擦洗管的内、外壁,烘干后调换Y型管的气味源与对照源的方向,以消除误差。
用一次性无味透明塑料袋分别套住健康的甜罗勒植株,下部进气,上部出气,作为气味源,另一位置使用相同空气塑料袋为空白对照。每头试虫仅测试一次,每测试5头更换气味源。大草蛉雌、雄成虫对营养期和开花期的甜罗勒植株分别测试50头,共计200头。
1.3 不同发育阶段甜罗勒的挥发物质收集与鉴定
采用顶空固相微萃取方法对甜罗勒营养期的叶、开花期的叶和花的挥发物质进行收集测定分析。将不同时期健康的甜罗勒营养期的叶、开花期的叶和花放入玻璃瓶(45 mL,Cleman),利用固相微萃取针SPME纤维中(DVB/CAR/PDMS, 50/30 μm)萃取挥发物质30 min。
采用气相色谱-质谱联用仪(GC-MS)进行挥发物质成分检测。质谱柱为HP-5和DBWAX(30 m×250 μm×0.25 μm),氦气用作载气流速为1.7 mL/min,GC进样口设置为不分流模式。柱初始温度60℃,保持2 min,以10℃/min程序升温至250℃,保持10 min,再以20℃/min升温至320℃,保持3 min;氦气流速为3.0 mL/min;离子源EI为70eV、350V;扫描质量为30~600 amu;进样口温度250℃,接口温度280℃。每个试验重复3次。
1.4 大草蛉对不同浓度下标准品的嗅觉行为反应
测试大草蛉对不同浓度下主要挥发物质标准品嗅觉行为的方法同1.2.1。将10μL的标准品滴在一张3 cm×0.5 cm的滤纸条上放进气味瓶内作为处理气味源,以石蜡油作为对照气味源。分别测试大草蛉雌成虫对0.01 μg/ L和0.1 μg/L浓度的6种标准品试验,每个重复90头,共计1 080头大草蛉雌成虫成功选择测试。
1.5 数据分析
试验所得数据Excel 2019进行整理,然后使用SPSS Statistics 22.0软件进行统计分析。采用χ2检验分析大草蛉雌/雄成虫对处理组与对照组选择差异显著性。选择率(%)=选择处理组虫数/总头数×100。若值为正,为诱集率,若值为负,则为驱避率;采用NIST05 标准谱库对挥发物质进行定性,用挥发物质的峰面积进行相对含量定量比较及分析。
2 结果与分析
2.1 大草蛉对不同时期甜罗勒植株嗅觉行为选择
甜罗勒营养期和开花期的植株对大草蛉雌、雄成虫对吸引作用不同(图1)。开花期的甜罗勒植株对大草蛉雌成虫具有极显著吸引作用,对雄虫也有一定的吸引作用,但与对照相比无显著差异(雌开花期:χ2=9.680,P<0.05;雄开花期:χ2=0.720,P=0.396)。营养期的甜罗勒植株对大草蛉雌、雄成虫均无吸引作用,且显著性差异相差较小(雌营养期:χ2=0.720,P=0.396;雄营养期:χ2=0.320,P=0.572)。

图1 大草蛉对不同时期甜罗勒植株的嗅觉反应Fig.1 Olfactory response of Chrysopa pallens to Ocimum basilicum plants at different stages
2.2 甜罗勒不同发育阶段叶片和花挥发物质成分
不同时期甜罗勒叶片和花挥发物质成分大致相同,相对含量在0.5%以上的共鉴定出28种挥发化合物(表1),甜罗勒营养期的叶有19种、开花期的叶和花分别有21、22种化合物。其中共有的化合物组分包含13种,主要为芳樟醇、丁香酚、α-香柑油烯、桉油素、β-榄香烯等。这些挥发物化学成分属性为醇类、萜烯类、酯类等化合物。两个不同时期甜罗勒叶片挥发物化学组分中,含量依次为芳樟醇(22.64%~30.64%)、α-香柑油烯(9.05%~15.1%)、丁香酚(3.41%~13.23%)、β-榄香烯,仅这四种物质相对含量就占总含量37.92%~63.28%。叶片中芳樟醇、丁香酚从营养期到开花期的相对含量逐渐下降;萜品油烯、β-月桂烯、罗勒烯相对含量逐渐上升。其中,有些挥发性组分仅在某一阶段出现,如樟脑、γ-衣兰油烯只在花中被检测到;β-水芹烯、乙酸龙脑酯仅在花期叶片中被检测到;桧烯只在营养期叶片中出现。
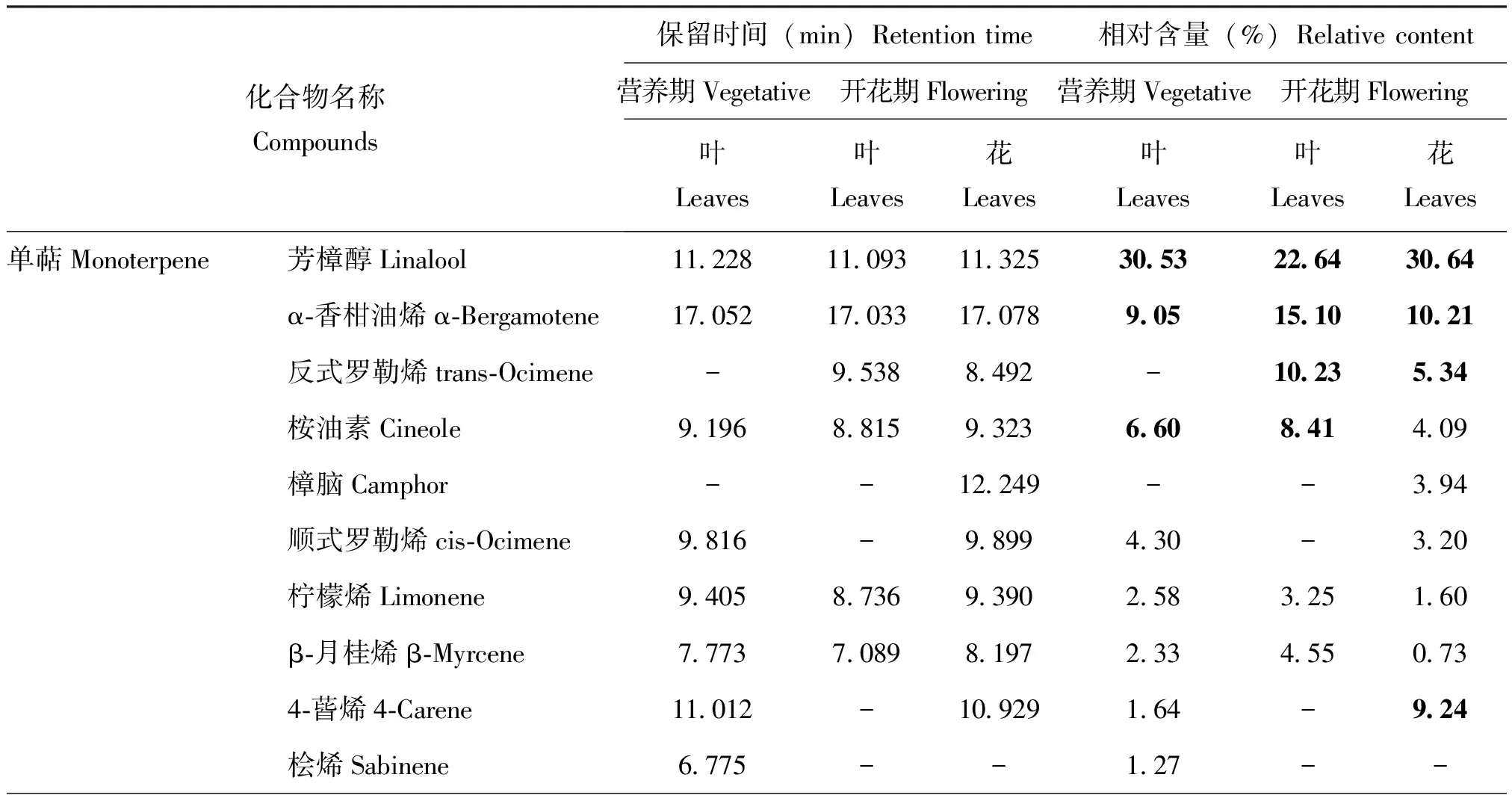
表1 不同时期甜罗勒叶和花的化学成分
甜罗勒在营养期和花期释放的挥发性有机成分及相对含量动态变化发现,花期的甜罗勒含量较高的挥发物为烯萜类,其中单萜和倍半萜相对含量之和达95.5%。其中芳樟醇含量最高,相对百分比含量为30.64%。随着甜罗勒植株的生长发育,单萜类物质的在营养期到开花期的叶片中相对含量逐渐升高,在花中达到峰值74.86%。倍半萜类物质含量的相对含量仅升高了4.71%,其中愈创烯从营养期到开花期相对含量就升高了4.56%。丁香酚在叶中从营养期到开花期的相对含量升高4%左右,但在花中含量仅是叶中的25.8%。
2.3 大草蛉对不同浓度标准品的嗅觉行为反应
在甜罗勒主要挥发物质中选取6种芳樟醇、罗勒烯、萜品油烯、丁香酚、樟脑、β-榄香烯标准品,测定了不同浓度时对大草蛉雌虫的嗅觉行为选择。结果表明,0.1 μg/L浓度的芳樟醇对大草蛉具有显著的吸引作用(P<0.05);0.01 μg/L浓度的β-榄香烯对大草蛉具有极显著地吸引作用(P<0.01)。大草蛉对0.1 μg/L、0.01 μg/L浓度的萜品油烯和0.01 μg/L浓度的樟脑、丁香酚、芳樟醇、罗勒烯等挥发物不显著的趋向选择(图2)。
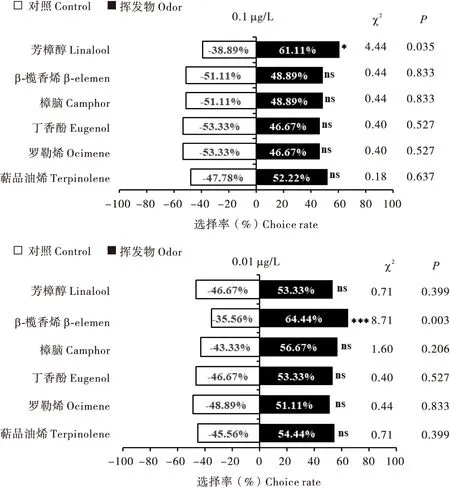
图2 大草蛉对不同浓度主要挥发物质标准品的嗅觉行为选择Fig.2 Selection of olfactory behavior of Chrysopa pallens to different concentrations of volatile odors
3 结论与讨论
植物在不同生长发育时期会产生多种挥发物质,其组成和丰度的差异与植物生长发育代谢有关(付国需等, 2008; Desurmontetal., 2015),本研究发现了芳香植物甜罗勒不同发育阶段叶和花的挥发性物质相对含量动态变化情况。植物挥发气味物质可能不会影响植株本身的生长发育,但却是植物与昆虫之间联系的信号物质(苏建伟等, 2020)。本研究关注了不同时期甜罗勒植株不同组织的挥发物质对天敌昆虫大草蛉的嗅觉行为影响。开花期的甜罗勒植株可显著引诱大草蛉雌虫,而对雄虫也有引诱作用但不显著。这可能与大草蛉雌雄触角上气味结合蛋白,ORs基因特异性表达不同有关(王娟, 2016)。天敌昆虫利用植物挥发物进行搜寻、产卵、逃避、取食、聚集等行为,依靠灵敏的嗅觉识别机制可简化复杂的气味环境并做出反应(盛子耀等, 2019; 王冰等, 2021)。由此推测,大草蛉对甜罗勒偏好性可能与不同时期挥发物质不同有关,这可能有助于大草蛉取食、遮蔽或产卵等行为。
芳香植物挥发物的类型主要包括烃类、醇、醛、酮、酯、有机酸和萜烯类,其中萜烯类化合物中种类相比其他化合物种类丰富(Songetal., 2017; Hattetal., 2019)。萜烯类化合物在植物-植食性昆虫-天敌昆虫三级营养级关系中起着重要作用,一方面在天敌搜寻害虫过程中常起信号定位作用(James, 2003; 李小龙等, 2014; 陈晓亚等, 2015)。另一方面在天敌栖境选择、产卵等发挥作用(Zhuetal., 2005)。本研究发现甜罗勒叶在营养期和开花期中含有大量单萜化合物(60.37%~76.55%)。这与Kpoviessietal.(2014)报道的丁香罗勒在营养期和开花期的单萜类挥发物组分相似(56.72%、55.87%)。其中单萜化合物被报道是天敌搜寻害虫过程中重要的定位信号物质(Fontanaetal., 2011)。本研究发现甜罗勒所有时期叶片和花相对含量最多的单萜化合物-芳樟醇,尤其是在花中高达30.64%。许多研究报道了芳樟醇对多种天敌及传粉昆虫具有吸引作用,例如烟草Nicotianatabacum被取食后释放的芳樟醇吸引天敌攻击虫卵从而降低害虫为害(Kessleretal., 2001);国槐Styphnolobiumjaponicum释放的芳樟醇对瓢虫有显著的引诱作用(薛皎亮等, 2008);茶树Camelliasinensis中释放的芳樟醇对中华蜜蜂Apiscerana有显著引诱作用(吴国火等, 2020)。本研究中发现芳樟醇对大草蛉的引诱效果有浓度差异性,在0.1 μg/L浓度比0.01 μg/L浓度时对大草蛉有显著的引诱作用。
本研究结果表明0.1 μg/L浓度时萜品油烯、樟脑、β-榄香烯、丁香酚和罗勒烯这5种挥发物对大草蛉具有一定趋避趋势,但在0.01 μg/L浓度却表现出吸引趋势。昆虫对同一种化合物的反应浓度不同,主要是受自身相关蛋白表达量和敏感性影响(狄贵秋等, 2019)。这也表明捕食性天敌对化合物的选择是一个复杂的过程。浓度差异可区分不同植物或者植物生理状态,而昆虫定位的与化合物浓度有关,在一定化合物浓度范围中昆虫才会产生反应,同一种植物挥发物可能具有多种功能,会因浓度不同从而对害虫及其天敌产生吸引或驱避作用(Altuzaretal., 2010; 蒋娜娜等, 2018)。
此外,芳香植物具有强烈的挥发物质直接影响天敌种群,而且花粉或花蜜也可以为天敌昆虫提供营养、延长其寿命和提高繁殖力(Wäckers &Rijn., 2012; Parolinetal., 2012)、种群增长(Venzonetal., 2016; Hattetal., 2019)、提供避难所(Venzonetal., 2016)和产卵场所(Laubertieetal., 2012; Andréetal., 2017)等功能,间接降低害虫的数量,达到控害的效果。比如当缺乏猎物时,开花期甜罗勒植株可显著吸引草蛉Ceraeochrysacubana雌成虫,其花粉或花蜜作为补充食物用于草蛉成虫和幼虫的生长繁殖(Batistaetal., 2017)。甜罗勒作为储蓄植物时,对捕食性天敌温室中矮小长脊盲蝽Macrolophuspygmaeus的种群增长虽没有显著影响,但是却能直接趋避番茄Lycopersiconesculentum上温室白粉虱Trialeurodesvaporariorum种群数量,以达到较好的控害效果(Parolinetal., 2015),这充分证明了芳香植物在生物防治中的应用中的潜力。本研究发现不同时期甜罗勒对大草蛉的诱集作用差异,也为田间应用甜罗勒使用时期提供了支撑数据。尽管本研究中仅测定了大草蛉对甜罗勒6种主要挥发物质的嗅觉行为,未来随着化学分析技术的提升,必将推进对大草蛉在化学生态学方面的研究。此外,近年来使用引诱剂来增加作物生态系统中天敌昆虫的补充和定殖是增强保护性生物防治的新尝试。例如水杨酸甲酯不仅可以能增加农业生态系统中草蛉种群数量,而且对瓢虫、食蚜蝇、寄生蜂的种群也有增益效果(Thmingetal., 2021)。因此,未来继续深入探究具有诱集作用的化合物按不同比例混配成复合引诱剂,可以最大发挥对大草蛉的引诱作用。这些有助于提升功能植物增效天敌生物防治效率,优化可持续控害的生态调控策略。
猜你喜欢
农业与技术(2023年1期)2023-01-31
广州化工(2021年19期)2021-10-25
孩子(2019年2期)2019-02-20
天然产物研究与开发(2018年1期)2018-02-02
中国调味品(2017年2期)2017-03-20
中国调味品(2017年2期)2017-03-20
广西林业科学(2016年2期)2016-03-20
中国房地产业(2016年18期)2016-02-18
经济林研究(2015年3期)2015-12-21
天然产物研究与开发(2014年6期)2014-04-27
