
C源与NP添加对Cd胁迫下林地土壤呼吸作用的影响
2024-01-23 05:35孙劲伟王圣燕范弟武朱咏莉
南京林业大学学报(自然科学版) 2024年1期
孙劲伟,王圣燕,范弟武,朱咏莉,2,3*
(1. 南京林业大学生态与环境学院,江苏 南京 210037;2. 南京林业大学 南方现代林业协同创新中心,江苏 南京 210037;3. 江苏洪泽湖湿地生态系统国家定位观测研究站,江苏 洪泽 223100)
毒物兴奋效应(Hormesis)指生物体暴露在低浓度胁迫时会出现刺激性响应而高浓度时受到抑制的现象[1-3]。Hormesis是对传统环境毒理学线性与阈值模型的挑战,对环境基准与质量标准制定与污染修复理念等产生深远影响。近年来,为进一步探索Hormesis在生态系统尺度的表达及其潜在的污染修复价值,Hormesis研究从传统的生物对象转向更为复杂的生物-非生物复合体[4-6]。
土壤是目前已知的最复杂的生物-非生物复合体。Han等[7]以土壤本身为试验模型(experimental model),土壤碱性磷酸酶(ALP)活性为测试终点(endpoint),研究并揭示Cd胁迫下的土壤Hormesis效应。Wang等[8]通过评估土壤中微生物数量、组成以及群落多样性来研究土壤微生物群落对外源Cd的刺激反应,并进一步探究群落水平的Hormesis效应及其在生态风险评估中的应用。Fan 等[9]研究不同土地利用类型土壤酶活性的Hormesis效应,发现自然覆被下土壤ALP对外源Cd胁迫具有更高的可塑性;但人为干扰下土壤ALP对外源Cd胁迫的抵抗能力更强,Hormesis效应参数因而可用来预测土地利用变化对土壤质量与功能的影响。土壤呼吸是土壤中的植物根系、食碎屑动物、真菌和细菌等进行新陈代谢活动,消耗有机物产生CO2的过程,是当前土壤Hormesis效应研究中除土壤酶和土壤微生物之外最具潜力的测试终点。以往有关Cd对土壤呼吸影响的研究中,Chen等[10]发现1.5~6.0 mg/kg的Cd均抑制了土壤呼吸作用;Zalaghi等[11]研究表明,添加Cd(50、100 mg/kg)显著降低土壤呼吸作用;Yao 等[12]认为N沉降加剧了Cd对土壤呼吸的抑制效应。显然,以往研究结果大多显示为Cd对土壤呼吸的抑制效应,有关土壤呼吸作用潜在的Hormesis效应还鲜见报道。这可能是大多实验设计中对低剂量的设置过少导致Hormesis效应很难被捕捉到,以及不同研究中使用的土壤类型与性状等差异大,刺激效应或抑制效应对应的剂量大相径庭。
土壤C、N、P等生源要素是影响Hormesis效应表达的重要因素。理论上,通过外源添加的方式能够更好地揭示相关因子的影响及机制,但由于土壤本身的形成与组分十分复杂,特别是本底C、N、P含量差异大,往往掩盖了所添加组分的真实影响。以往研究大多通过相关分析或路径分析等来评估这些因子的影响[7, 9, 13]。近年来,模式土壤(artificial standard soil)在环境毒理学研究中得到越来越广泛的应用[14-17]。模式土壤的组分与含量固定,且不具备生物学特征,替代真实土壤可以显著提高研究结果的可比性与可重复性,是揭示Hormesis效应相关机制的理想基质。为此,本研究以模式土壤为对象,通过接种林地土壤微生物,研究外源添加C源与矿质营养N、P时,Cd胁迫下土壤呼吸作用潜在的Hormesis效应,以期为揭示土壤毒物兴奋效应机制和探索相应调控措施提供理论依据。
1 材料与方法
1.1 供试材料
采用文献[18]中提到的模式土壤(69.5%石英砂、20%高岭土、10%泥炭土、0.5%碳酸钙等组成)为土壤基质。添加的C源为葡萄糖,矿质营养N、P为硫酸铵[(NH4)2SO4]和磷酸二氢钾(KH2PO4),试剂均为分析纯。
土壤微生物接种液的制备:土壤样品采自尚未受到重金属污染的新疆伊犁杨树林0~20 cm土层,风干,用木锤碾碎过2 mm筛。称取10 g风干土于150 mL锥形瓶,添加50 mL去离子水。25 ℃下150 r/min震荡24 h。此后,将此土壤溶液在4 ℃下2 450 r/min离心5 min。上清液即为微生物接种液。对接种液的微生物群落结构与多样性进行测定,细菌群落主要为变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidota),其相对丰度分别为52.2%~71.7%、148%~31.3%和5.9%~19.6%。
1.2 实验设计
称取20.0 g灭菌(121 ℃下高温灭菌20 min)的模式土壤于250 mL无菌培养瓶中,加入4 mL上述土壤接种液,用无菌封口膜封住瓶口,置于25 ℃相对湿度为30%的恒温培养箱48 h。此后,设置单独添加C源(GC)、单独添加矿质营养(NP),同时添加C源和矿质营养NP(GC+NP)以及二者均无添加的对照(CK),共计4个处理。其中:GC处理添加葡萄糖溶液的含量为12 mg/kg(干土);NP处理为添加 (NH4)2SO4和KH2PO4混合溶液,其中N和P的含量分别为1.0和0.5 mg/kg;(GC+NP)处理的C、N、P含量为GC和NP之和;CK处理加入等体积超纯水。4个处理加入溶液的体积均为4 mL。
胁迫梯度的设置参照中国(GB 15618—2018)以及欧美国家、东亚国家或地区土壤Cd污染环境质量标准值[19-22],在添加C源与NP后预培养6 h,再分别加入CdCl2溶液,形成0、0.01、0.02、0.04、0.10、0.20、0.30、0.40、0.60、1.00、2.50、5.00、10.00、13.00、20.00和30.00 mg/kg等16个Cd胁迫梯度,并分别于处理后的0、6、12、24、48 h时测定土壤呼吸速率。
1.3 气体样品的采集与分析
采集气样前,去除培养管透气膜,用橡胶塞塞住无菌培养瓶,分别在密闭0和20 min后,用注射器采集顶空气体10 mL。使用安捷伦7890 A气相色谱分析仪(Agilent 7890A Series GC)测定气样中的CO2浓度,采用热导检测器(TCD),载气为高纯N2与He和95%的Ar与5%的CH4混合气。标准气由中国计量科学研究院提供(595669#),高纯N2作为反吹气。通过标准气体和待测气体的峰面积比值计算样品中的CO2浓度。土壤呼吸速率(R)的计算如下:
(1)
式中:R为土壤呼吸速率,mg/(kg·h);C1为富集20 min后CO2的质量浓度,mg/m3;C0为富集开始时的CO2的质量浓度,mg/m3;V为培养瓶的体积,mL;m为培养瓶中土样的质量,g。
1.4 刺激或抑制效应计算
Cd胁迫下土壤呼吸速率的增加和降低分别定义为“刺激”或“抑制”。用刺激幅度(M)反映刺激或抑制效应的大小。
(2)
式中:M指刺激率或者抑制率,%;An为Cd胁迫下的土壤呼吸速率,mg/(kg·h);A0为无Cd添加(超纯水)时的土壤呼吸速率,mg/(kg·h)。
1.5 葡萄糖与NP添加后土壤呼吸作用下交互作用的测算
采用土壤呼吸速率净变化量(ΔR)判断葡萄糖与NP添加的交互作用:
ΔR=(RGC+NP-RCK)-(RGC-RCK)+(RNP-RCK)。
(3)
式中:RGC+NP、RCK、RGC和RNP为同时添加葡萄糖与NP、空白对照,以及单独加入葡萄糖和单独加入NP处理的土壤呼吸速率,mg/(kg·h)。当ΔR=0时,表示葡萄糖与NP添加对土壤呼吸作用影响的交互作用为加合作用;ΔR>0时,表示二者为协同作用;ΔR<0时,则为拮抗作用。
1.6 数据处理
采用SPSS 19.0对数据进行统计分析,以单因素方差分析法比较不同处理下与不同Cd剂量间土壤呼吸速率(均以平均值±标准差表示)的差异显著性(P<0.05)。
2 结果与分析
2.1 不同梯度Cd胁迫下土壤呼吸速率的变化特征
不同梯度Cd胁迫下,不同培养时间土壤呼吸速率变化见图1。由图1a、1b可见,CK处理下,0 h时,土壤平均呼吸速率在Cd剂量为0.01~0.60 mg/kg区间从1.6增至最高5.7 mg/(kg·h),此后相对稳定在4.0~5.3 mg/(kg·h)范围内。添加Cd后土壤呼吸速率均表现出明显的增加,平均刺激幅度(M)为77.9%~522.6%。土壤呼吸速率均低于对照(Cd剂量为0.0 mg/kg)时,6 h内变化在2.2~9.5 mg/(kg·h),相应的M值为-81.1%~-19.2%。12 h且当Cd剂量为0.01~0.60 mg/kg时,土壤呼吸速率明显高出对照,M值在140.9%~1 306.6%范围变化。此后,土壤呼吸速率快速下降。24 h和48 h时,所有Cd胁迫下土壤呼吸速率均高于对照,分别在1.6~4.8 mg/(kg·h)和1.3~4.9 mg/(kg·h)变化,对应的M在59.8%~199.1%和51.7%~288.7%变化。不同培养时间下,土壤呼吸的刺激或抑制响应不同,呈现非连续性变化特征。

图1 不同处理(CK、GC、NP和GC+NP)下土壤呼吸速率在Cd胁迫时的变化特征Fig. 1 Changes of soil respiration under Cd stress in CK, GC, NP and GC+NP treatments
GC处理下,不同培养时间土壤呼吸速率的变化均相对稳定,0、6、12、24和48 h时平均为(8.9±2.9)、(16.8±2.8)、(23.2±3.9)、(21.4±3.8)和(11.1±2.9) mg/(kg·h)。M值分别为-59.0%~-5.3%、-34.2%~16.5%、-46.8%~7.3%、-43.4%~-9.7%和-63.2%~0.4%(图1c、1d)。
NP处理下,0 h时的土壤呼吸速率在Cd剂量为0.1和0.4 mg/kg时有两个明显的刺激峰,表现出显著的Hormesis效应,对应的M值分别为262.6%和219.2%。12 h时,Cd剂量为0.02 mg/kg条件下的土壤呼吸速率显著高于对照,也表现出显著的Hormesis效应,M值为4.4%。相比较而言,6、24和48 h时,土壤呼吸速率相对稳定,分别在1.9~3.0、1.9~2.7和1.2~2.9 mg/(kg·h)范围变化(图1e、1f)。
GC+NP处理下,0 h且当Cd剂量为0.02、0.40、2.50、13.00 mg/kg时,土壤呼吸速率显著升高,表现出显著的Hormesis效应,M值分别为106.0%、83.5%、40.9%和23.5%。6 h时,土壤呼吸速率相对稳定在13.4~19.3 mg/(kg·h)。相比较而言,12和48 h且Cd剂量为0.01 mg/kg时,土壤呼吸速率表现为短暂但显著的刺激效应,也表现出显著的Hormesis效应,M值分别为19.7%和19.1%。24 h时,所有Cd剂量下土壤呼吸速率均低于0 mg/kg(对照),在16.3~29.3 mg/(kg·h)范围变化,M值为-53.8%~-17.1%(图 1g、1h)。
2.2 C源与NP添加对Cd胁迫下土壤呼吸速率的影响及交互效应
分别扣除GC、NP和(GC+NP)3个处理的CK背景值,在全培养期(0~48 h)分析Cd胁迫条件下C源与NP添加对土壤呼吸速率的影响及二者的交互效应,结果见图2。添加NP后,不同Cd剂量下土壤呼吸速率在-2.7~0.2 mg/(kg·h)范围变化,C源(GC-CK)、C源与NP同时添加[(GC+NP)-CK]下,土壤呼吸速率分别在8.0~18.5和6.5~16.9 mg/(kg·h)范围变化。可见,添加NP使土壤呼吸速率显著降低。此外,无Cd添加时,C源添加与NP添加对土壤呼吸的影响表现为拮抗;当Cd剂量为0.01~0.20 mg/kg时,C源添加与NP添加对土壤呼吸的影响表现为连续的协同效应(表1)。此后,二者之间表现为协同与拮抗效应的交替出现。
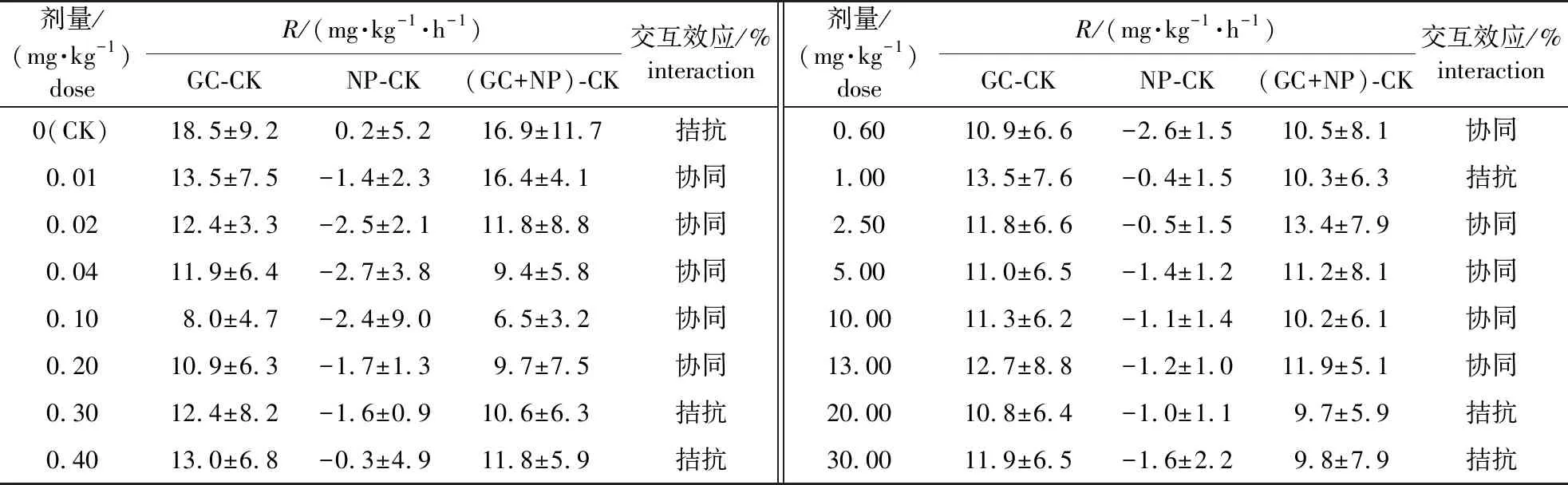
表1 各剂量不同处理单独对土壤呼吸速率影响的差异

图2 C源与NP添加对Cd胁迫下土壤呼吸速率的影响Fig. 2 Effects of C and N and P addition on soil respiration rate under Cd stress
3 讨 论
以往有关Hormesis效应研究的试验模型主要包括植物、动物和微生物等[23-25]。近年来,随着土壤污染研究的逐步深入,试验模型开始从生物个体向生物-非生物复合体延伸。多位研究者以土壤本身为试验模型,以土壤碱性磷酸酶活性和土壤微生物相对丰度为测试终点开展了重要探索[7-8, 13, 26]。这些研究表明,污染胁迫下,Hormesis现象以土壤酶或微生物群体响应的方式表达在群落、甚至生态系统尺度。除土壤酶和土壤微生物外,土壤呼吸作用是土壤微生物群体效应主要的表达方式之一。以往有关研究表明,Cd对土壤呼吸作用的刺激现象较少报道,抑制效应的阈值浓度差异很大。如,周涵君等[27]发现土壤呼吸速率随Cd浓度的升高而降低;Alfaro 等[28]发现,Cd含量为40 mg/kg时土壤基质中微生物呼吸降低73.6%~87.6%;Stefanowicz 等[29]发现,只有当Cd含量超过400 mg/kg时才会对土壤微生物生物量、酶活性和丛枝菌根真菌有显著的负面影响。本研究以模式土壤为基质,通过设置103剂量跨度的Cd水平(0.01~30.00 mg/kg)探索C、NP添加对土壤呼吸Hormesis效应的影响,发现NP处理和GC+NP处理下Cd诱导土壤呼吸作用表现出明显的Hormesis效应。特别是NP处理下Cd剂量为0.10和0.40 mg/kg时,以及GC+NP处理下Cd剂量为0.02、0.40、2.50、13.00 mg/kg时,均出现“多重”Hormesis现象。Erofeeva[3]在102~105范围的污染物剂量区间观测麦苗生长生理指标等的变化,发现Hormesis现象的多次出现。产生这一现象的原因被认为是麦苗在不同胁迫压力下采取了不同的适应机制(adaptive mechanism)。对土壤而言,“多重”Hormesis现象的出现可能是对Cd产生响应的微生物群落结构与多样性在不同剂量下发生了显著变化。以往有关重金属类型对微生物的选择效应研究较多:如Chen 等[30]基于Cu、Zn和Cd的选择压力探索微生物的功能作用,发现属水平的抗性、敏感型和重金属富集微生物。Zhao等[31]发现稻田土壤中的As和Cd通过对细菌丰度的影响主导着抗生素抗性基因的分布。有关剂量约束下土壤微生物的响应研究目前还很不充分。低剂量下土壤微生物的类群、丰度与功能可能与高剂量下的存在显著差异,土壤微生物对Cd剂量的选择效应可能导致较宽的剂量区间多次出现Hormesis现象。此外,从产生刺激效应的最大Cd剂量看来,最为接近的是德国、日本、捷克、中国台湾和法国等目前采用Cd的土壤环境质量相关标准。从出现Hormesis现象的暴露时间看来,0 h的响应最为明显。尽管12和48 h也有响应,但响应的幅度与剂量宽度明显降低。因此,可以认为,Cd胁迫下土壤呼吸作用的Hormesis效应主要发生在早期,具有“即时”响应特征。与以往报道的土壤酶与土壤微生物Hormesis效应的时间特征相比[7-8],土壤呼吸作用的Hormesis效应对时间的响应更为灵敏。
添加N、P后土壤净呼吸速率在-2.7~0.2 mg/(kg·h)之间变化,这可能是由于N的添加所致。Wang 等[32]认为土壤呼吸受到来自施N肥后各种细菌群落的影响,这些细菌群落包括酸杆菌门、绿屈菌门、变形杆菌门和拟杆菌门,最终有利于增加土壤有机碳的储存。另一方面,NH4+-N的添加不但增加了土壤微生物生物量,还通过降低土壤pH使土壤酶活性下降,最终导致土壤呼吸速率下降[33]。此外,添加N对土壤碳氮比(C/N)的改变也是土壤呼吸速率下降的重要原因,如Tao 等[34]发现N添加会抑制土壤有机物的分解。当面临逐渐加剧的Cd胁迫时,添加C源与N、P对土壤呼吸速率交互作用的影响依次表现为拮抗(无Cd添加)、协同效应(0.01~0.20 mg/kg)和协同与拮抗效应的交替出现(>0.20 mg/kg)。这表明土壤呼吸在面临逐渐增加的Cd胁迫时,对C源与NP添加的交互效应会发生转变。即对葡萄糖和NP添加产生响应的微生物随Cd剂量的增加从低剂量时的“竞争”转向高剂量下的“合作”。本研究结果与目前植物与动物面临资源限制时的胁迫梯度假说(the stress-gradient hypothesis)[35]相似。该假说认为,随着非生物因子胁迫的增加,物种间的关系将从相互竞争转为相互合作。如Campos等[36]发现两种昆虫在食源(落叶)质量高的条件下相互竞争,而当落叶营养质量降低时相互合作。在C源限制条件下,植物对N转移和C在丛枝菌根的分配表现出互惠共生的支持功能[37]。随着水分和养分胁迫强度的增加,形成土壤结皮的地衣在空间的排列从普遍的种间分离转变为聚集[38]。然而,有所不同的是,胁迫梯度假说主要指的是生源要素的限制。本研究中土壤呼吸作用的这一现象是在污染物(重金属Cd)胁迫下发生的,这可能是对胁迫梯度假说的重要拓展与延伸。相关效应的机理研究可能需要基于污染物的剂量与暴露时间深入分析相关过程微生物的差异。
总之,本研究中外源添加NP条件下,Cd诱导土壤呼吸表现出显著的Hormesis效应。Hormesis效应主要出现在胁迫初期,并且在0.02~13.00 mg/kg Cd剂量范围内,土壤呼吸速率表现出“多重”Hormesis交替出现的现象。当面临逐渐增加的Cd胁迫时,添加C源与NP对土壤呼吸速率交互作用的影响依次表现为拮抗效应(无Cd添加)、协同效应(0.01~0.20 mg/kg)和协同与拮抗效应的交替出现(>0.20 mg/kg)。
猜你喜欢
核科学与工程(2021年4期)2022-01-12
今日农业(2020年19期)2020-12-14
学生天地(2020年14期)2020-08-25
中学生数理化·高一版(2020年6期)2020-07-25
特别文摘(2018年3期)2018-08-08
中学生数理化·八年级物理人教版(2017年3期)2017-11-09
中学物理·高中(2016年12期)2017-04-22
诗选刊(2015年6期)2015-10-26
上海金属(2014年5期)2014-12-20
食品工业科技(2014年23期)2014-03-11
