
不同添加剂对越冬后许氏平鲉生长、抗氧化及免疫性能的影响
2024-01-26 08:19王晓艳王成强孙晓宇李培玉郝甜甜黄利娜李宝山黄炳山
广东海洋大学学报 2024年1期
王晓艳,王成强,孙晓宇,李培玉,郝甜甜,黄利娜,李宝山,黄炳山
(1.山东省海洋资源与环境研究院/山东省海水渔用饲料工程技术研究中心/水生动物营养与饲料研发创新示范平台/山东省海洋生态修复重点试验室/烟台市海珍品质量安全控制与精深加工重点实验室,山东 烟台 264006;2.济宁市任城区行政审批服务局,山东 济宁 272000)
许氏平鲉(Sebastes schlegelii)隶属于鲉形目(Scorpaeniformes)鲉科(Sebastidae)平鲉属(Sebastes),主要分布于我国渤海、黄海、东海,以及朝鲜半岛、日本、鄂霍次克海,是一种近岸冷水肉食性鱼类,具有耐低温、抗病强的特点,适宜人工鱼礁资源增殖和网箱养殖[1]。近年来,随着深水网箱养殖技术的快速发展,许氏平鲉网箱养殖的规模和产量也在逐年增加。我国北方海域冬季严寒,越冬期间不投喂,在长期低温和饥饿的双重压力下,鱼类免疫系统会发生重大调整以适应环境变化[2]。长达4个月的低温和饥饿应激会对鱼体健康产生明显损伤,而在越冬后通过普通饲料投喂很难在短时间内帮助机体恢复健康[3],因此迫切需要寻找能够快速恢复鱼体机能的方法。目前已有鱼类越冬期间的生理变化及死亡率[4-5]的研究,但未见关于海水网箱养殖鱼类越冬后营养调控的研究报道。因此,本研究针对越冬后许氏平鲉生理特点,分别通过复合菌制剂、抗菌肽及植物多酚3 种不同的营养策略对越冬后的许氏平鲉进行营养干预,以期寻找快速恢复鱼体机能的方式,提高网箱养殖效率,同时为网箱养殖鱼类越冬后功能饲料的开发提供理论依据。
1 材料与方法
1.1 实验饲料
以鱼粉、酪蛋白和发酵花生粕为主要蛋白源,以鱼油为主要脂肪源,设计粗蛋白质量分数约为52%,粗脂肪质量分数约为11%的基础饲料(对照),在对照组饲料表面使用复合益生菌[丁酸梭菌(Clostridium butyricum)10 mL/kg、复合乳酸菌15 mL/kg、芽孢杆菌(Bacillus)15 mL/kg,与等体积水混匀后,均匀裹在饲料表面]为复合益生菌组,在基础配方中添加质量分数分别为0.5%抗菌肽(山东深海生物股份有限公司提供)和0.05%植物多酚(山东礼恩生物科技有限公司提供),为抗菌肽组和植物多酚组。基础饲料组分质量分数见表1,饲料常规成分组成见表2。
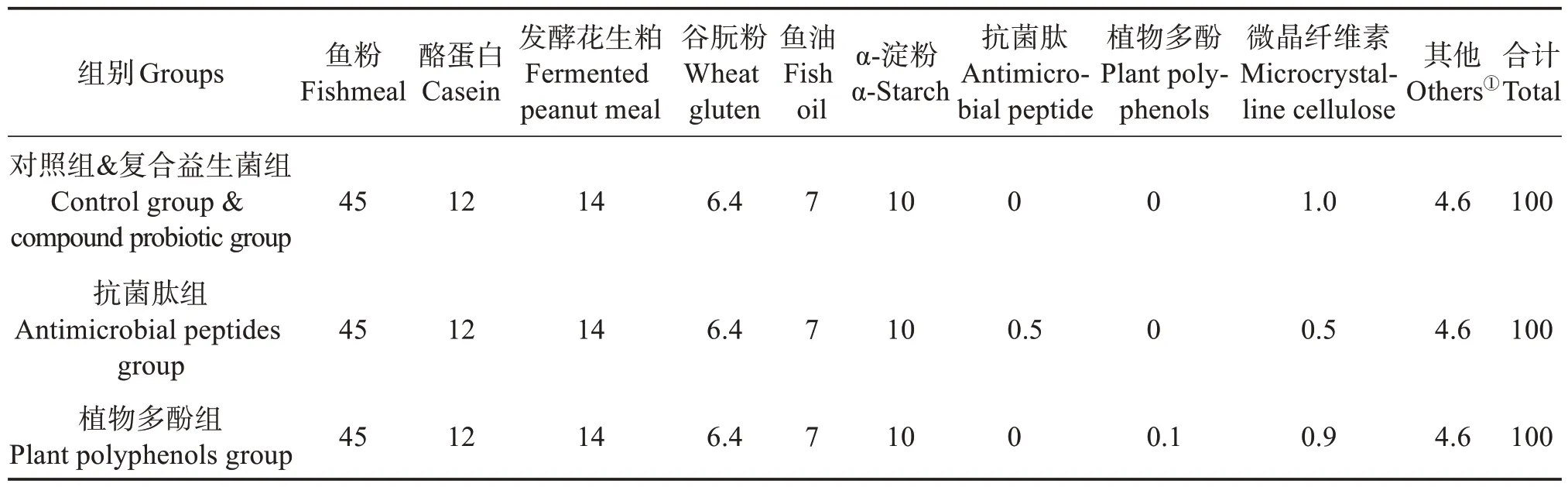
表1 基础饲料组分质量分数Table1 Mass fraction of basic diet components %

表2 实验饲料常规成分质量分数及能量(干基)Table 2 Mass fraction of nutrients and energy of experimental diets(dry mass basis)
1.2 实验对象及饲养管理
养殖实验在山东省海洋资源与环境研究院循环水养殖系统内进行,选用同一批次在自然海域网箱中越冬约4 个月(12 月初至次年4 月初)的许氏平鲉作为实验对象,用对照组饲料驯养2 周以适应养殖系统及饲料。正式实验开始前挑选出144尾初体质量(Initial body mass,IBM)为(249.13±0.58)g、大小均匀、体质健壮的许氏平鲉,随机分为4组,每组3个重复,每个重复12 尾鱼。养殖实验进行21 d,每天定时定量投喂两次(8:00 和16:00),初始投喂量约为体质量的1%,根据摄食情况调整投喂量至表观饱食,投喂30 min 后排残饵。驯养及实验期间控制水温(14±1)℃,溶氧>6 mg/L,pH 7.6~8.2,氨氮和亚硝酸氮质量浓度均小于0.1 mg/L。
1.3 样品采集与实验方法
养殖结束后,禁饲24 h,MS-222 (100 mg/L)麻醉后,每桶鱼计数并称末体质量(Final body mass,FBM),以计算增重率(Weight gain rate,WGR)和存活率(Survival rate,SR)。每桶随机取6 尾鱼,尾静脉取血,分离肠道用于炎症相关基因相对表达量的测定。所有操作均在无菌冰盘内完成,肠道样品置于无菌冻存管中经液氮速冻后保存于-80 ℃冰箱待测,血液于4 ℃静置4 h 后离心(4 000 r/min,10 min),取上清液保存于-20 ℃冰箱待测。
血清总抗氧化能力(T-AOC)、超氧化物歧化酶(SOD)、丙二醛(MDA)、溶菌酶(LZM)、酸性磷酸酶(ACP)、免疫球蛋白M(IgM)、补体3(C3)、总蛋白(TP)、白蛋白(ALB)、谷丙转氨酶(ALT)及谷草转氨酶(AST)活性均使用南京建成生物工程研究所的试剂盒测定。
1.4 qPCR测定
根据本实验室转录组测序结果及许氏平鲉基因组(CNP0000222)数据,查找白介素(Interleukin,IL)IL-1β、IL-15、IL-12b,Toll 样受体8(Toll-like receptors,TLR8)基因序列,在NCBI Nucleotide BLAST 进行同源性比对,确定同源性较高的基因序列,使用Primer Premier 5.0软件设计引物(序列见表3),以核糖体蛋白L17 (RPL17) 为内参基因。使用SPARKeasy Improved Tissue/Cell RNA Kit 进行肠道样本总RNA 提取,使用Nanodrop®2000 (Thermo Fisher Scientific,USA)检测RNA 浓度及纯度,琼脂糖凝胶电泳检测RNA完整性及基因组污染情况,使用SPARKscript II RT Plus Kit(with gDNA Eraser)去除基因组DNA 并反转录成cDNA。使用2X SYBR Green qPCR Mix(with ROX)进行qPCR(LightCycler®480II)分析。按照2-ΔΔCt计算目的基因相对表达量。
表3 许氏平鲉肠道相关基因引物序列Table 3 Sequence of primers for gut related genes of Sebastes schlegelii
1.5 数据统计
所有数据采用SPSS 18.0 进行单因素方差分析(one-way ANOVA),用Duncan’s 检验进行多重比较分析,P <0.05 时认为差异显著。统计数据以平均值±标准差表示。
2 结果
2.1 不同添加剂对越冬后许氏平鲉生长性能的影响
由表4 可见,抗菌肽组和植物多酚组末体质量和增重率显著高于对照组和复合益生菌组(P <0.05),复合益生菌组与对照组差异不显著(P >0.05);各组间存活率差异不显著(P >0.05)。

表4 不同添加剂对越冬后许氏平鲉生长性能的影响Table 4 Effects of different additives on growth performance of Sebastes schlegelii after overwintering
2.2 不同添加剂对越冬后许氏平鲉抗氧化性能的影响
由表5 可见,不同添加剂对越冬后许氏平鲉的血清抗氧化性能具有显著影响。抗菌肽组T-AOC显著高于对照组、复合益生菌组(P <0.05),植物多酚组与其他各组差异不显著(P >0.05);植物多酚组SOD 活性显著高于其他各组(P <0.05),复合益生菌组、抗菌肽组与对照组差异不显著(P >0.05);各添加组MDA 含量显著低于对照组(P <0.05),以植物多酚组含量最低。
2.3 不同添加剂对越冬后许氏平鲉非特异性免疫性能的影响
由表6 可见,植物多酚组血清LZM 活性显著高于对照组、复合益生菌组(P <0.05),复合益生菌组、抗菌肽组与对照组差异不显著(P >0.05),但抗菌肽组显著高于复合益生菌组(P <0.05);各组间ACP 活性差异不显著(P >0.05);各处理组IgM 与对照组差异不显著(P >0.05),但抗菌肽组、植物多酚组显著高于复合益生菌组(P <0.05);植物多酚组C3显著高于其他各组(P <0.05),复合益生菌组、抗菌肽组与对照组差异不显著(P >0.05)。
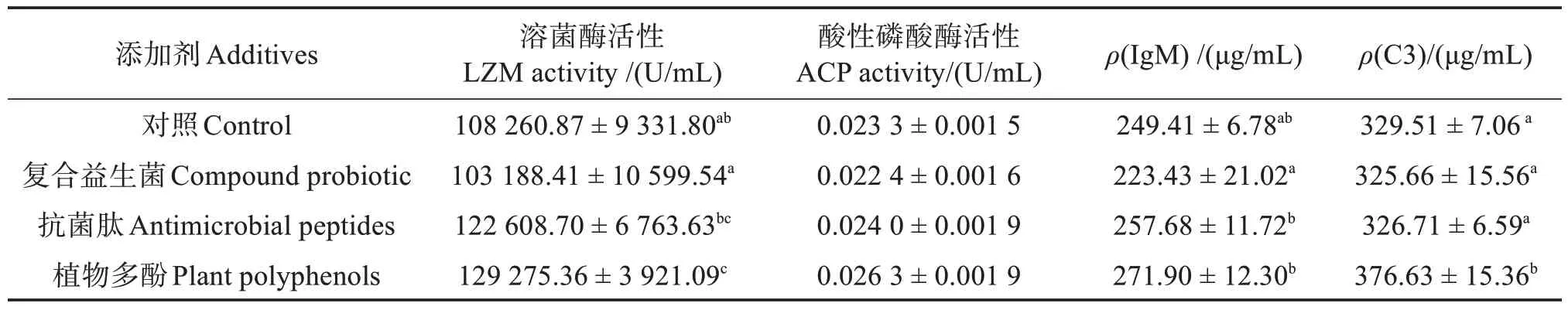
表6 不同添加剂对越冬后许氏平鲉血清非特异性免疫的影响Table 6 Effects of different additives on serum non-specific immunity of Sebastes schlegelii after overwintering
2.4 不同添加剂对越冬后许氏平鲉血清生化指标的影响
由表7 可见血清总蛋白、白蛋白受不同饲料添加剂影响显著(P <0.05),植物多酚组总蛋白、白蛋白显著高于其他各组(P <0.05),复合益生菌组、抗菌肽组与对照组差异不显著(P >0.05);各组间谷丙转氨酶和谷草转氨酶活性差异均不显著(P >0.05)。

表7 不同添加剂对越冬后许氏平鲉血清生化指标的影响Table 7 Effects of different additives on serum biochemical indexes of Sebastes schlegelii after overwintering
2.5 不同添加剂对越冬许氏平鲉肠道炎症因子和抗炎因子基因表达的影响
由图1可见,不同饲料配方对许氏平鲉肠道炎症因子IL-1β基因相对表达量影响不显著(P >0.05);复合益生菌组和抗菌肽组IL-15表达量有下调趋势,但与对照组差异不显著(P >0.05),植物多酚组显著低于其他各组(P <0.05);各处理组TLR8均下调,抗菌肽组和植物多酚组显著低于对照组(P <0.05);抗菌肽组IL-12b表达量显著高于对照组、复合益生菌组(P <0.05),植物多酚组与各组无显著差异(P >0.05)。
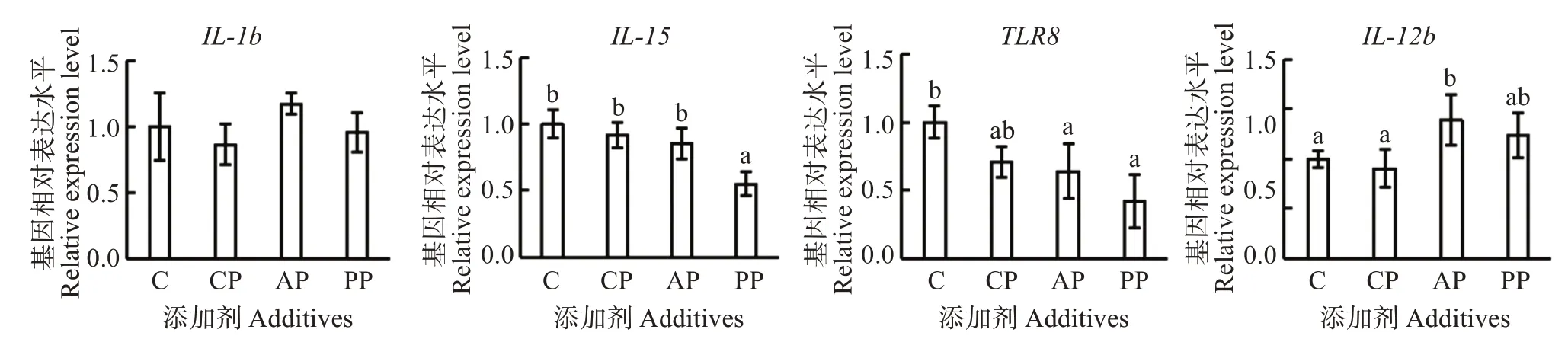
图1 不同添加剂对越冬后许氏平鲉肠道炎症因子和抗炎因子基因相对表达量的影响Fig.1 Effects of different additives on the relative expression levels of intestinal inflammatory factor and anti-inflammatory factor genes of Sebastes schlegelii after overwintering
3 讨论
3.1 生长性能
在我国北方网箱养殖过程中,冬季鱼类需要同时面对低温和饥饿双重挑战,鱼类处于免疫低下状态,生长停滞。本实验探究复合益生菌、抗菌肽、植物多酚3种添加剂对越冬后许氏平鲉生长性能的影响,结果显示,在越冬后恢复投喂阶段,相比较普通饲料投喂,在基础配方中添加抗菌肽和植物多酚显著提高了越冬后许氏平鲉的生长性能,且二者的促生长作用均优于复合益生菌。本研究添加的抗菌肽是从微生物中提取的小分子多肽,在消化道内易被蛋白酶降解成氨基酸,是环保、高效、无毒的新型抗生素替代品[7]。湘云鲫(Carassius auratus)[8]和大口黑鲈(Micropterus salmoides)[9]饲料中添加抗菌肽均显著提高了生长性能,与本研究结果一致。本研究所使用的植物多酚中含有鼠尾草酸、白藜芦醇、单宁、槲皮素、地榆素等植物多酚,这几种植物多酚在鱼类上研究较少,对珍珠龙胆石斑鱼(Epinephelus fuscoguttatus♀×Epinephelus lanceolatus♂)[10]研究发现,质量分数为0.1%的水解单宁能显著提高实验鱼生长性能,茶多酚等其他多酚可以提高马口鱼(Opsariichthys bidens)[11]和团头鲂(Megalobrama amblycephala)[12]等鱼的生长性能。据报道,益生菌的添加能提高中华鳖(Pelodiscus sinensis)和草鱼(Ctenopharyngodon idella)的生长性能[13],但本研究中,复合益生菌组实验鱼的生长性能并未与对照组产生显著差异,可能是因为复合菌中的丁酸梭菌具有酸臭味道,影响鱼类摄食积极性,进而影响实验鱼的生长性能。
3.2 抗氧化性能
机体正常代谢中会保持抗氧化系统的动态平衡,一旦平衡被打破,过多自由基会对机体造成氧化损伤。鱼类在低温和饥饿条件下会增加脂肪酸过氧化[14],而脂质过氧化不仅会降低细胞膜的流动性,还会产生大量MDA,损害机体健康。在鱼类越冬后恢复投喂阶段若不及时清除自由基、恢复机体抗氧化功能,会影响鱼类生产效率。本研究相比较对照组,复合益生菌组虽然T-AOC 和SOD 活性并未有明显升高,但MDA 含量显著降低,其原因一方面可能是益生菌的代谢产物如丁酸等短链脂肪酸抑制了氧化应激,另一方面可能是益生菌通过Nrf2-Keap1-ARE途径上调涉及活性氧簇(ROS)解毒的基因,保护机体免受氧化应激,从而抑制了氧化产物MDA 的产生[15]。本研究抗菌肽和植物多酚均显著提高了鱼体抗氧化能力、降低了MDA含量。抗菌肽的抗氧化功能主要由其特定结构域决定,其肽键或肽链的构象和氨基酸组成密切相关[16]。湘云鲫饲料中添加天蚕素抗菌肽,血清SOD 活性显著提高,MDA 含量显著降低[8];采用抗菌肽浸泡(84 U/mL)及饲料添加(50 mg/kg)两种处理方式处理虹鳟(Oncorhynchus mykiss)初孵仔鱼均显著提高SOD 活性[7]。植物多酚的抗氧化功能可归因于其含有酚羟基结构,其邻位酚羟基易被氧化成醌类结构,具有很强捕捉自由基能力[17]。1~20 mg/mL 白藜芦醇强化卤虫12 h后投喂斑马鱼(Barchydanio rerio),显著提高了肠道抗氧化能力[18];松浦镜鲤(Cyprinus carpio)饲料添加160 mg/kg 白藜芦醇,肝脏SOD 活性显著升高,MDA 含量显著降低[19];500 mg/kg 茶多酚可缓解四氯化碳诱导的团头鲂急性肝应激损伤,显著提高肝肠组织SOD 水平,降低MDA 含量[20]。以上研究均证明了植物多酚在抗氧化方面的有效性。
3.3 非特异性免疫
血清LZM、ACP、IgM 及C3 是鱼类重要的非特异性免疫指标。有研究证明,益生菌可通过调节肠道菌群、黏膜屏障功能及炎症反应来提高机体非特异性免疫,大口黑鲈(M.salmoides)饲料中添加质量分数为1%的枯草芽孢杆菌(Bacillus subtilis)制剂,血清ACP 和LZM 活性显著升高[21]。而本研究复合益生菌并未表现出提高机体免疫力的作用,这与复合益生菌组较低的生长性能相呼应。对斜带石斑鱼(Epinephelus coioides)[22]、杂交鲟(Acipenser gueldenstaedti♀×Acipenser schrenckiBrandt♂)[23]等鱼的研究证明,抗菌肽可增强鱼类免疫器官活力,提高LZM活性,增强机体非特异性免疫[24],从枯草芽孢杆菌中提取的抗菌肽(质量分数0.2%)添加在饲料中,团头鲂血清补体C3显著提高[25],本研究中抗菌肽的添加显著提高了血清IgM 水平,对LZM 活性也有一定程度的提高,提示了抗菌肽对提高鱼类非特异性免疫的积极作用。植物多酚对机体免疫力的作用在许氏平鲉[26]、马口鱼[11]、草鱼[27]、奥尼罗非鱼(Oreochromis aureus×O.niloticus)[28]、松浦镜鲤[19]等鱼类上均得到证实:茶多酚的添加显著提高了许氏平鲉[26]、马口鱼[11]和奥尼罗非鱼[28]血清或血浆LZM 活性;20、40 mg/kg的α-苯基苯并吡喃(植物多酚)提高了草鱼血清LZM 活性[27]。本研究中,添加植物多酚组血清LZM、IgM、C3 得到显著提高,提示植物多酚对越冬后许氏平鲉的抗病力具有明显的促进作用。
3.4 血清生化指标
血清TP是血清各种蛋白质的复合物,具有维持血管内正常胶体渗透压和酸碱度的功能,ALB 是血液中的主要蛋白成分,具有结合和运输内源性与外源性物质、维持血液胶体渗透压、清除自由基等功能,二者水平高低可体现机体蛋白质代谢水平的高低[29]。本研究中,复合益生菌和抗菌肽的添加对血清TP 和ALB 水平均无显著影响,但植物多酚组TP、ALB均显著高于其他各组。复合益生菌的添加对中华鳖血清TP 和ALB 也没有显著影响[13],与本研究结果一致。将枯草芽孢杆菌中提取的抗菌肽(质量分数0.2%)添加在饲料中,团头鲂血清TP 显著提高;在斜带石斑鱼上也证明了抗菌肽能提高血清TP 和ALB 水平[22],本研究得到不一致的结果,可能是因为抗菌肽种类不同。对仔猪(Sus scrofadomesticus)[30]、肉鸡(Gallusdomesticus)[31]的研究均证明植物多酚能有效提高血清TP、ALB 水平,提示植物多酚在促进肝脏合成代谢方面的有效作用。
AST是活性最高的转氨酶,参与肝细胞内的氨基酸代谢,ALT是催化转氨酶的特异性酶,正常情况下二者在血清中含量较低,在肝细胞损伤时会出现血清中含量异常升高[32]。对中华鳖[13]、鲤(C.carpio)[32]和许氏平鲉[26]的研究分别证明了饲料中益生菌、抗菌肽和植物多酚的添加不影响实验鱼血清AST、ALT 活性,本研究获得了一致结果,提示本研究中复合益生菌、抗菌肽及植物多酚的添加未引起肝细胞损伤。
3.5 肠道炎症因子和抗炎因子基因相对表达量
炎症过程特点是免疫细胞从血管向炎症部位迁移,同时大量释放促炎介质,包括细胞因子、趋化因子和氧化剂(如ROS)等[33]。据报道,益生菌可通过调节肠道免疫相关基因的表达来调节免疫功能:复合益生菌[枯草芽孢杆菌、嗜酸乳杆菌(Lactobacillus acidophilus)及红法夫酵母(Phaffia rhodozyma)]可有效抑制嗜水气单胞菌(Aeromonas hydrophila)引起的鲤鱼肠道炎症基因IL-1β的过表达,并上调抗炎因子IL-10的表达量[34];枯草芽孢杆菌显著抑制了2,4,6-三硝基苯磺酸(TNBS)诱导的大口黑鲈肠道IL-1β、IL-15、IL-8等促炎因子的表达,上调抗炎因子1L-10的表达[21]。本研究中,虽然复合益生菌的添加对IL-1β、IL-15及TLR8的表达量有一定下调趋势,但与对照组差异不显著,且IL-12b的表达也未上调,提示本实验中复合益生菌在炎症调节方面无明显优势。
目前普遍认为抗菌肽一方面能通过膜渗透作用直接杀灭微生物,另一方面能通过免疫调节作用发挥其抗菌功能,即通过刺激免疫信号通路和相关受体蛋白的表达,调节多种免疫相关基因和细胞因子,中和巨噬细胞和单核细胞等释放的促炎细胞因子,从而减轻炎症反应[35-36];本研究中,抗菌肽的添加显著下调了TLR8的表达量,上调了IL-12b的表达量,证实了以上观点。
研究证明,NF-κB信号通路能促进黏附分子、细胞因子和其他促炎介质的表达,而NF-κB是多种植物多酚的首选分子靶点之一,植物多酚主要通过抑制NF-κB信号通路发挥抗炎作用[33]。研究发现,绿茶多酚可抑制促炎细胞因子TLR4 信号通路而减轻肠道炎症[37];单宁酸等植物多酚能够减轻葡聚糖-硫酸钠(DSS)诱导的小鼠溃疡性结肠炎,炎症细胞因子水平降低[38];槲皮素能通过调节鸡胚胎自噬和程序性细胞死亡来改善脂多糖诱导的十二指肠炎症[39]。本研究得到的结果与上述结果类似。本研究中,相比较对照组,抗菌肽和植物多酚在抗炎功能上具有明显优势。
4 结论
本实验条件下,相比较对照组饲料,植物多酚和抗菌肽的添加能显著提高越冬后许氏平鲉生长、抗氧化及非特异性免疫性能,促进肝脏合成代谢,调节炎症因子和抗炎因子基因表达,复合益生菌则效果不明显。
猜你喜欢
今日农业(2021年21期)2021-11-26
现代畜牧科技(2021年9期)2021-10-13
水产养殖(2021年5期)2021-05-21
保健与生活(2018年17期)2018-01-27
现代园艺(2017年11期)2017-06-28
广东饲料(2016年5期)2016-12-01
国外医药(抗生素分册)(2016年5期)2016-07-12
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
特产研究(2014年4期)2014-04-10
