
水稻不育系包颈特性及穗颈节间形态解剖结构的研究
2010-06-08 04:07张桂莲张分云杨烨雷东阳陈立云
湖南农业大学学报(自然科学版) 2010年1期
张桂莲,张分云,杨烨,雷东阳,陈立云
(湖南农业大学 水稻科学研究所,湖南 长沙410128)
目前生产上利用的籼型杂交水稻不育系都存在着不同程度包颈(穗不能完全从剑叶叶鞘中抽出)的遗传障碍,极大地制约了不育系异交结实率的提高,降低了制种产量[1-3].虽然生产上在不育系始穗期应用赤霉素可部分解除不育系的包颈问题,但适合使用赤霉素的时期较短,易受阴雨天气的影响,而且赤霉素的大量使用不仅会增加制种的成本,加重田间穗上发芽及稻粒黑粉病的发生,影响种子的质量,还会污染环境[4-6],因此,必须从根本上解除不育系包颈的遗传障碍.1981年,Rutger等[7]发现了第一个隐性长穗颈基因. 随后中国的水稻研究者也相继报道由一对隐性高秆基因控制的突变体[8-10].笔者以包颈程度较轻的水稻品系H7408和包颈程度较重的水稻不育系培矮64S为材料,研究不育系包颈特性及穗颈节的形态解剖结构,旨在探明水稻不育系的包颈机理,为生产上选育不包颈的水稻不育系提供理论依据.
1 材料与方法
1.1 材 料
水稻品系 H7408,为湖南农业大学水稻科学研究所选育;培矮64S,由国家杂交水稻工程中心提供.
1.2 方 法
试验于 2008年在湖南农业大学水稻试验田进行.5月25日播种,6月18日移栽,常规肥水管理. 成熟期测量植株的秆高、倒一节间长、剑叶鞘长和穗颈节伸出度,取 30株的平均值作为各性状指标.
开花盛期每个材料随机取样5株,每株取主穗当天开放的3朵颖花混合捣碎,碘─碘化钾溶液染色后置100倍光学显微镜下镜检3个视野,计算不育度,取其平均值作为该株不育度观察值.
将定长的H7408和培矮64S的穗颈节间切成若干段,节间两端每段长0.7 cm,节间中部每段长1.5 cm,用 FAA液固定,经酒精脱水,二甲苯透明及石蜡包埋后进行连续纵向切片,切片经1.0%的番红和苯胺蓝染色. 在显微镜下对各小段切片中靠髓腔一侧的第一列薄壁细胞(内层薄壁细胞)和紧靠厚壁纤维细胞层的最外一列薄壁细胞(外层薄壁细胞)进行计数,并用显微测微尺计量细胞的长度,然后用OLYMPUSBH-2型显微摄像机,选择有代表性的部位进行拍照. 根据整个穗颈节间细胞的长度,将细胞长于50 μm的区段称为中部区段,由中部区段至倒一节的区段称为基部区段,由中部区段至穗颈节的区段称为顶部区段,并分别对不同区段的细胞数目和细胞长度进行统计分析.
2 结果与分析
2.1 培矮64S和H7408包颈特性的比较
培矮64S的包颈率为100%,包颈长度为11.12 cm ,花粉可染率(育性)为1.68%,而H7408包颈率为 14.86%,包颈长度为 0.86 cm ,花粉可染率为98.5%,两者差异显著.相关分析结果表明,包颈率、包颈长度与花粉可染率分别呈负相关关系(R=-0.468 1;R=-0.564 4).
2.2 培矮64S和H7408茎秆各节间长度的比较
从表1可看出,培矮64S的茎秆高度比H7408短23.42 cm,倒一节间(即穗颈节间)长比H7408短11.88 cm,倒二节间、倒三节间、倒四节间也比H7408短.穗颈伸出度由倒一节间长度和剑叶叶鞘长度决定,培矮 64S剑叶叶鞘与 H7408相比增长1.69 cm,而培矮64S剑叶叶鞘比穗颈节长11.58 cm,H7408穗颈伸出度为1.99 cm,因此培矮64S有相当一部分的颖花被包在叶鞘中.
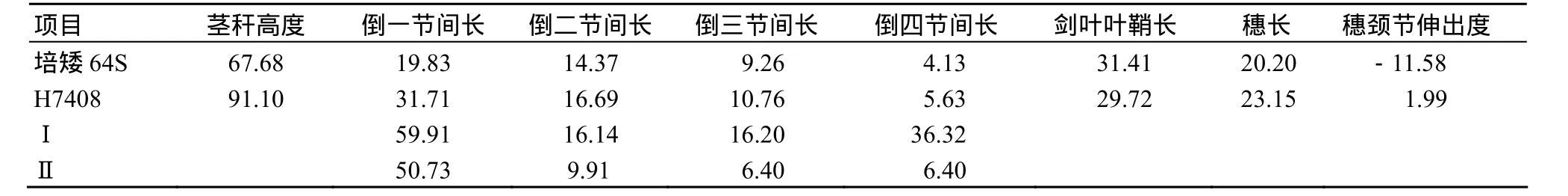
表1 培矮64S和H7408茎秆各节间长度及穗颈伸出性状Table 1 Comparison of length ang character of panicle exserted between Peiai64S and H7408 cm
2.3 穗颈节间细胞数目和细胞平均长度的比较
从表2可知,培矮64S穗颈节间外层薄壁细胞和内层薄壁细胞比H7408分别少1 016个和470个,说明穗颈节间薄壁细胞数量的多少与穗颈节间长度有密切关系. 培矮64S穗颈节间薄壁细胞的平均长度也比H7408短,其中以内层薄壁细胞平均长度差值较大,为17.8 µm,说明穗颈节间细胞长度增加是促进节间伸长的另一重要原因.

表2 穗颈节间细胞数目和平均长度Table 2 Comparison of the number and length of uppermost internode cell
按增加的细胞数与平均细胞长度的乘积来表示细胞数量增加对穗颈节间伸长的贡献率,H7408外层薄壁细胞数量增加使穗颈节伸长的长度为5.923 cm(表 3),占穗颈节总增加长度的51.19%;由外层薄壁细胞伸长使穗颈节伸长的长度为 5.957 cm,贡献率为 50.14%,说明外层薄壁细胞数量的增加与细胞长度增加对穗颈节伸长的贡献率基本相当. H7408内层薄壁细胞数量增加较少,对穗颈节伸长的贡献为 3.267 cm,仅占总增加长度的27.50%,内层薄壁细胞伸长的贡献则占72.50%,说明内层薄壁细胞主要靠细胞伸长促进穗颈节伸长.
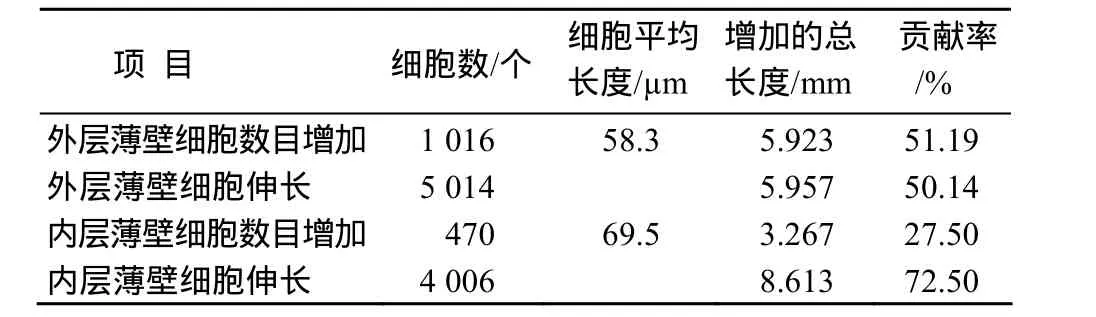
表3 细胞数和长度增加对节间长度伸长的贡献率Table 3 Comparison of uppermost internode elongation between cell number increasing and cell elongating
2.4 穗颈节间细胞长度的变化
从表4可知,2个材料的穗颈节间细胞(内、外层薄壁细胞)长度的变化规律是节间两端的细胞长度较小,中部区段细胞最长.培矮64S穗颈节间基部区段、顶部区段的细胞长度与H7408的相差不大(图 1-1,2和图 1-5,6),但中部区段相差较大(图1-3,4),内层薄壁细胞与外层薄壁细胞长度在中部区段培矮64S比H7408分别短25.2、37.8 µm.说明除节间两端外,培矮64S中下部区段薄壁细胞长度都比H7408短,始穗前后培矮64S穗颈节间伸长的长度过小,致使部分穗被包在叶鞘中.

表4 穗颈节间不同区段的细胞数和长度Table 4 Comparison of parenchyma cell number and length in different sections

图1 培矮64S和H7408穗颈节间不同区段细胞的比较(×100)Fig.1 Comparison of different sections of uppermost internode between Peiai64S and H7408(×100)
3 小结与讨论
有研究[11]表明,杂交水稻包颈现象不仅与不育细胞质有关,而且与细胞核也有密切关系.如果保持系最上节间足够长,就有可能减轻或消除不育系的包颈.本研究结果表明,包颈率、包颈长度与花粉可染率呈显著负相关关系.
植物细胞的生长或扩张的驱动力为膨压,它向四周的强度是均等的,而细胞不对称伸长原因在于细胞壁的作用,细胞壁中纤维素的排列方式直接影响着细胞的伸长.当纤维素微纤丝与细胞壁某一平面平行排列时,它就与伸展素、膨胀素等多种蛋白质交织,形成复杂的网络结构,从而限制了细胞这一方向的伸长[12-16].钟卫华等[17]对隐性长穗颈温敏核不育水稻长选 3S穗颈节间细胞学研究发现,其eui基因对不同位置细胞的作用不同,促进穗颈节间内层薄壁细胞伸长的作用明显大于促进其分裂的作用,紧接髓腔的薄壁细胞起运输营养物质作用. 本研究结果表明,穗颈节间伸长是由细胞数目增多和细胞长度增加双重作用所致,其中后者作用更为显著.对穗颈节各区段的细胞长度变化的比较研究表明,穗颈节间基部区段、顶部区段的细胞长度,培矮64S与 H7408的相差不大. 但在中部区段细胞长度差异显著,这说明穗颈节间中部区段细胞伸长是促进穗颈节间伸长的主要部位.季兰等[18]研究发现,赤霉素(GA)、生长素(IAA)、脱落酸(ABA)和乙烯影响水稻茎(或节间)的伸长,其中赤霉素与水稻茎伸长生长的关系最密切,因此关于穗颈节间伸长的机制还有待于从激素水平、分子机理方面进一步研究.
[1]袁隆平.杂交水稻学[M].北京:中国农业出版社,2002:247-255.
[2]雷东阳,陈立云,肖层林,等.超级杂交早稻陆两优996制种“九二O”施用技术[J].湖南农业大学学报:自然科学版,2007,33(1):1-4.
[3]杨远柱,唐平徕,杨文才,等.水稻低温敏核不育系陆18 的选育及育性表现[J].湖南农业大学学报:自然科学版,2000,26(3):155-159.
[4]杨孚初.“九二O”对杂交稻种子质量的影响[J].杂交水稻,1998,14(1):20-21.
[5]Lu Z M,Hong D L.Advances in hybrid rice seed production techniques[C]//Amarjit S,Basra. Heterosis and hybrid seed production in agronomic crops.New York:Food Products Press- an Imprint of the Haworth Press,1999:65-79.
[6]Oruno K,Kawai T.Variations of internode length and other characters in induced long-culm mutants of rice[J].Jpn Breed,1978,28:242-250.
[7]Rutager J N,Carnahan H L.A fourth genetic element to faciliate hybrid cereal production-a recessive tall in rice[J].Crop Sci,1981,21:273-276.
[8]LI An-xiang,LI Ci-hou,DING Ke-xin,et al.Synthetical treatment for kernel smut in hybrid rice seed production[J].Jiangsu Agricultural Sciences,1995(4):34-36.
[9]何祖华,申宗坦.水稻伸出度的遗传和不育系改良[J].中国水稻科学,1991,5(1):1-6.
[10]梁满中,钟卫华,徐孟亮,等.长穗颈温敏核不育水稻双低培 eS遗传特性的研究[J].作物学报,2004,4(4):290-294.
[11]杨仁崔,张书标,黄荣华,等.高秆隐性杂交水稻(e-杂交水稻)育种技术[J].中国农业科学,2002,35(3):233-237.
[12]郑家团,胡万星,谢华安,等.籼型水稻光温敏不育系包颈长度与育性的相关性研究[J].浙江农业大学学报,1998,24(6):627-630.
[13]Sauter M,Kende H.Levels of β-glucan and lignin in elongating internodes of deep wate rice[J].Plant Cell Physiol,1992,33(8):1089-1097.
[14]Cho H T,Kende H.Expansins and internodal growth of deep wate rrice[J].Plant Physiol,1997c,113:1145-1151.
[15]陈金桂,杨军,周燮.植物激素对微管和纤维素微纤丝排向的调节[J].生命科学,2001,13(3):139-142.
[16]郑湘如,王希善.植物解剖结构显微图谱[M].北京:农业出版社,1983:168-171.
[17]钟卫华,肖辉海,梁满中,等.隐性长穗颈温敏核不育水稻穗颈节间的细胞学观察[J].作物学报,2005,31(5):592-595.
[18]季兰,杨仁崔.水稻茎伸长生长与植物激素[J].植物学通报,2002,19(1):109-115.
猜你喜欢
汽车工程(2021年12期)2021-03-08
模具制造(2019年7期)2019-09-25
学苑创造·B版(2019年2期)2019-02-19
农机化研究(2018年2期)2018-06-05
课程教育研究(2017年28期)2017-08-29
植物保护(2016年1期)2016-09-14
学语文(2016年3期)2016-07-19
凿岩机械气动工具(2015年3期)2015-11-11
湖南农业(2014年9期)2014-04-02
汽车与新动力(2014年4期)2014-02-27
