
低温下不同抗寒性冬小麦叶片组织结构比较
2010-08-09 06:45周子珊牟永潮张永侠
东北农业大学学报 2010年4期
于 晶,周子珊,牟永潮,张永侠,苍 晶
(东北农业大学生命科学学院,哈尔滨 150030)
温度是影响植物生长及分布的重要环境因子之一。植物在对低温寒冷环境的长期适应中通过自身的遗传变异和自然选择,形成了抵御寒冷的能力即抗寒性。研究植物抗寒性的方法很多,主要包括:田间自然鉴定法、室内恢复生长法、电导法、镜检法、生理生化研究法、马林契克法、低温放热分析法及分子生物学研究法等。其中,采用镜检法(即借助于显微镜直接观察低温后细胞内部组织结构的动态变化)研究植物的抗寒性的报道很多,如苹果[1-2]、番荔枝[3]、唐古特红景天[4]、桃叶珊瑚[5]、圆柏[6]、小麦[7]、梨[8]、柑桔[9]、桃[10]、云杉[11]、松树[12]等,但上述报道多来自低温较敏感或木本植物,由于抗寒性不同,所以它们细胞的结构变化存在着一定差异。本试验对强抗寒冬小麦品种东农冬麦1号和弱抗寒冬小麦品种济麦22低温胁迫下的叶片进行解剖结构的观察,探讨了冬小麦叶片结构特征与其抗寒性的相关性,旨在为冬小麦生产及抗寒种质的选育提供相应依据。
1 材料与方法
1.1 材料
冬小麦(Triticum aestivum L.)品种东农冬麦1号和济麦22由东北农业大学小麦室提供。东农冬麦1号耐受的最低极限温度可达-30~-35℃,在黑龙江省可以越冬,返青率大于85%;济麦22耐受的最低极限温度为-10~-15℃,在黑龙江省不能越冬,返青率小于1%。
1.2 方法
试验材料大田生长季种植(9月),于最低温度稳定7 d通过5℃和-15℃时,每个材料随机选取10株,在靠近主脉处切取叶片,3次重复。
1.2.1 显微结构观察
制作石蜡切片:叶片(0.5 cm×0.5 cm)用FAA固定24 h,70%、85%、95%、100%梯度酒精渐次脱水(每一级间隔2 h),经1/2二甲苯+1/2酒精过渡到纯二甲苯透明,浸蜡包埋,切片(厚度约10 μm),番红-固绿双染色,中性树胶封片。显微镜观察并拍照。
1.2.2 超微结构观察
透射电镜样品制作:叶片(1 mm×3 mm小条)用2.5%、pH 6.7戊二醛溶液固定,经0.1 mol·L-1磷酸缓冲液冲洗3次,用1%锇酸固定,磷酸缓冲液冲洗3次后乙醇梯度脱水,Epon812包埋剂包埋,LKB-V型超薄切片机切片,醋酸双氧铀、柠檬酸铅双重染色,JEM-1200EX透射电镜观察、测定并拍照。
2 结果与分析
2.1 叶片显微结构比较
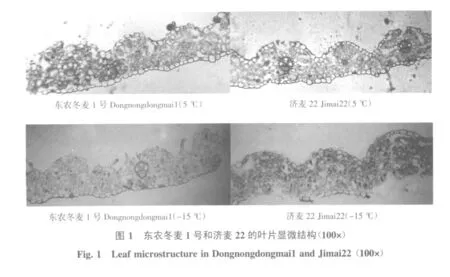
叶片距主脉中心1 cm处的横切面结构见图1。东农冬麦1号和济麦22在5℃和-15℃时均可清晰观察到表皮、叶肉细胞及维管束。上下表皮细胞狭长形,排列整齐;叶肉细胞无栅栏组织和海绵组织之分,全部由海绵状薄壁细胞组成,细胞间隙很大。在-15℃时,济麦22叶肉细胞排列较东农冬麦1号混乱。
2.2 叶片超微结构比较
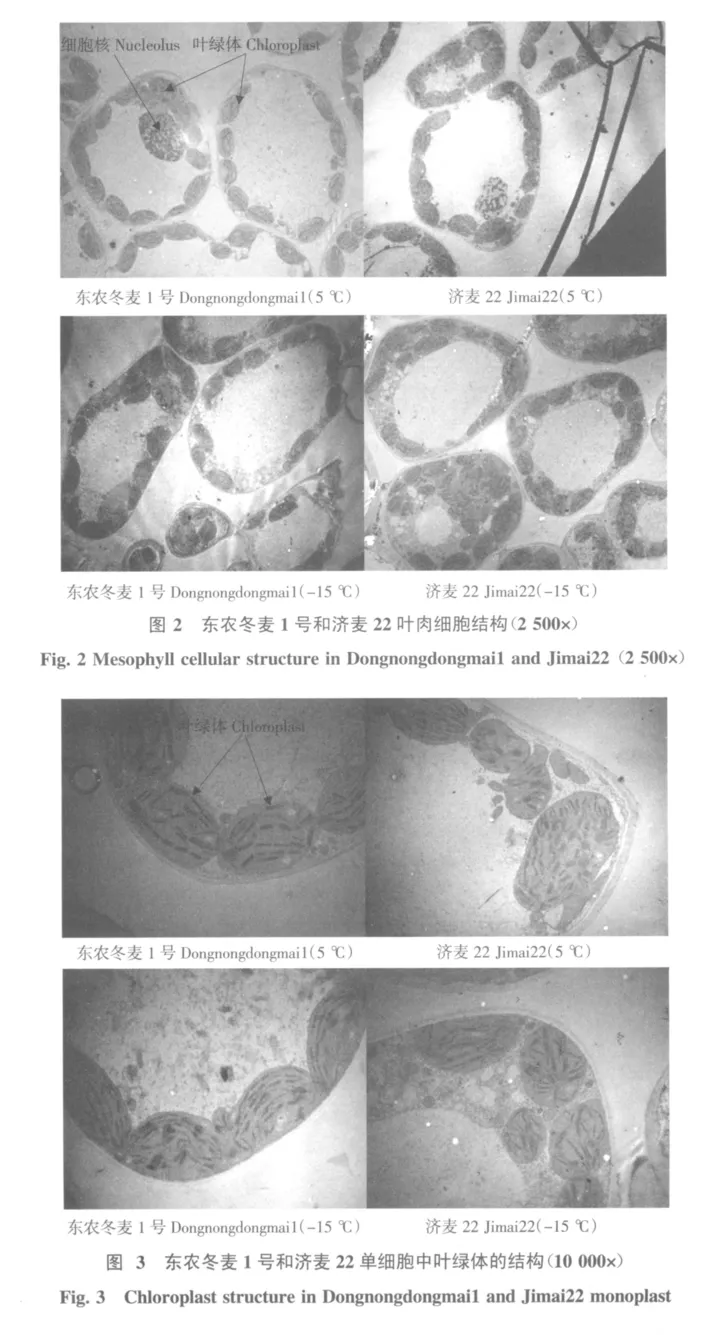
叶肉细胞的整体情况如图2所示。在5℃时,东农冬麦1号和济麦22的叶肉细胞中央均有一个大液泡,液泡周围是一薄层细胞质,内含有丰富的细胞器。叶绿体呈椭圆状,非常整齐地单行排列在细胞壁周边,细胞中清晰可见一个处于旺盛代谢状态的细胞核分布于细胞的一侧。内质网大多分布于细胞壁附近。线粒体多呈椭圆形,分布于两个叶绿体之间(见图3)。在-15℃时抗寒性较弱的济麦22部分细胞的液泡膜收缩明显,大液泡分割成几个小液泡,叶绿体向一侧聚集,排列发生混乱。细胞出现少量小空泡;而东农冬麦1号变化不大。从单个叶肉细胞中叶绿体的数目来比较,东农冬麦1号略多于济麦22,且-15℃低温时比5℃低温时多。
叶肉细胞中叶绿体结构见图3。东农冬麦1号在5℃和-15℃时,叶绿体结构均完整,呈椭圆形,类囊体基粒片层和基质片层结构发达,跺叠整齐,沿叶绿体的长轴方向平行排列,均匀分布于叶绿体中;济麦22在5℃和-15℃时,叶绿体内类囊体排列方向不规则,-15℃较5℃时方向更紊乱。叶绿体中类囊体的基粒片层和基质片层放大结构见图4。在5℃和-15℃低温时,东农冬麦1号和济麦22基粒片层和基质片层均很完整,未见空泡化现象。
由图5可知,在5℃时,东农冬麦1号和济麦22线粒体呈椭圆形,内部均匀分布着大量的嵴,嵴呈线形;而-15℃时,线粒体肿胀呈圆形,嵴的结构模糊或呈小泡状,济麦22有个别线粒体的嵴出现消失现象(如图5箭头所示)。
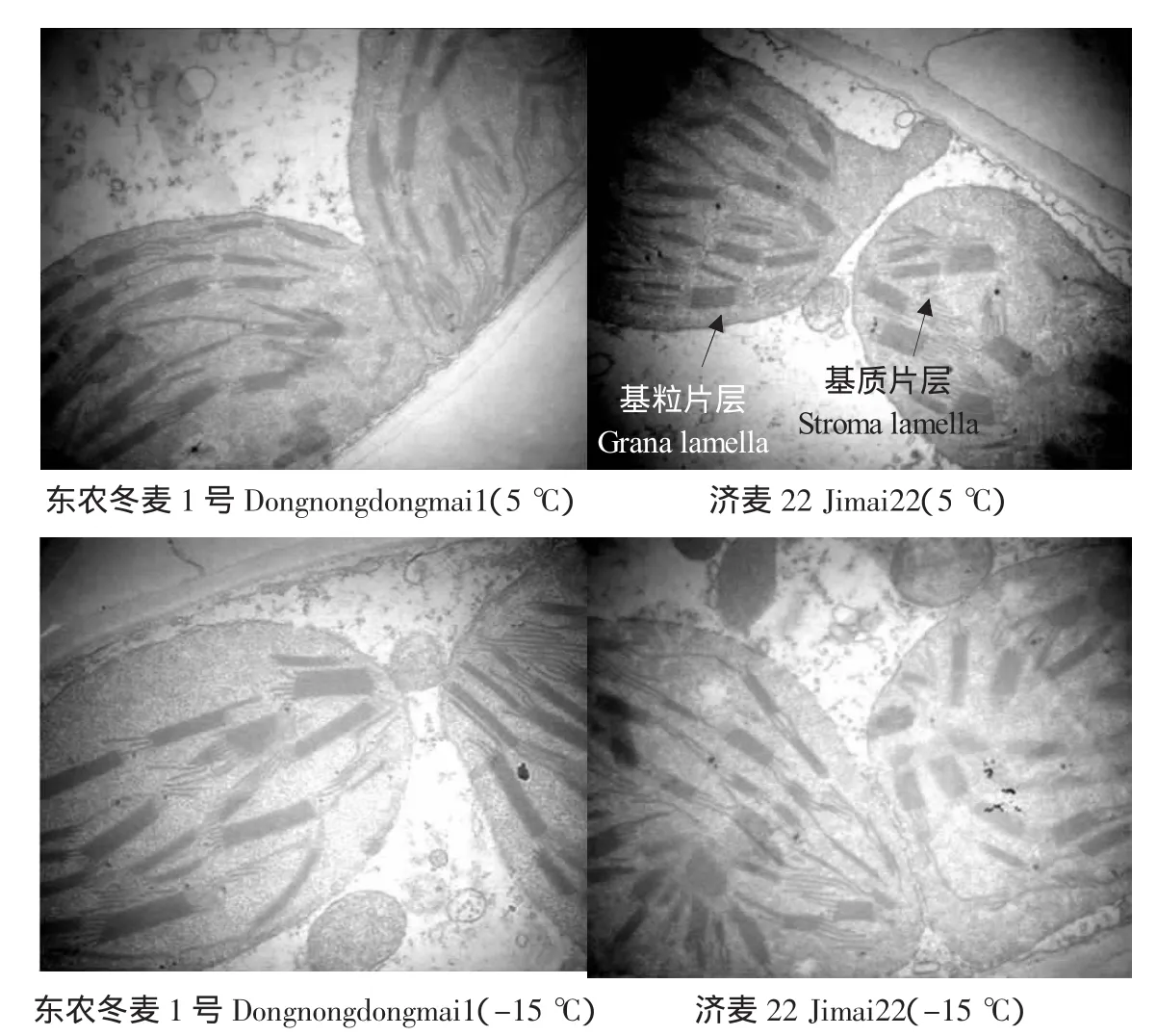
图4 东农冬麦1号和济麦22类囊体片层结构(30 000×)Fig.4 Thylakoid lamellar structure in Dongnongdongmai1 and Jimai22 monoplast(30 000×)
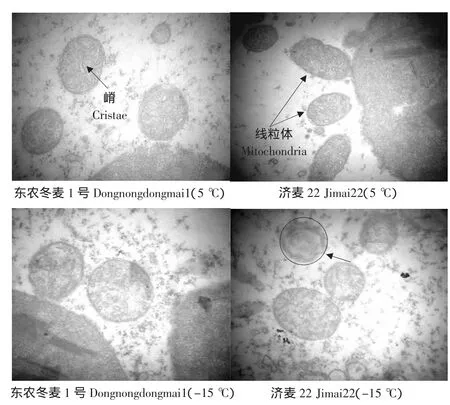
图5 东农冬麦1号和济麦22线粒体结构(50 000×)Fig.5 Mitochondria structure in Dongnongdongmai1 and Jimai22(50 000×)
3 讨论与结论
大量对植物抗寒性与解剖结构关系的研究指出抗寒的指标有多种,如伴随着低温出现细胞质壁分离现象[10]、细胞间隙大[6],叶绿体中淀粉粒减少[11-12],脂质球多[6],线粒体嵴消失[13]等。
苗芳等从小麦叶片横切片上观察到[14-15]:冷型小麦的叶肉细胞小,排列紧密,细胞层数较多;叶绿体数量多,叶绿体基粒片层丰富,叶肉细胞中由于大液泡的存在,叶绿体沿细胞质膜内侧排列;叶脉间的距离小,单位叶片宽度内维管束的数目多,横截面积大。叶肉细胞排列疏松、间隙大,有利于防止细胞内结冰[5];细胞体积小则表面积大,以便细胞内的水分经较短距离迁移出细胞,不易造成细胞内结冰伤害[16]。本试验结果表明,在5℃和-15℃低温下,东农冬麦1号和济麦22叶片的横切结构有很多相似之处,如均表现为叶肉细胞很小,细胞层数较多,叶肉细胞间隙较大,线粒体形状均为圆或椭圆形,均未发现淀粉粒及脂质球,与前人研究结果一致,即都表现出一定的抗寒结构特征。
董合铸等研究表明[7],强抗寒性冬小麦品种的叶绿体在-9℃处理2 d后结构未发生变化,中等抗寒性的品种叶绿体发生肿胀,趋向椭圆形,片层弯曲;弱抗寒性的品种叶绿体严重破坏,类囊体发生空泡化等;强抗寒和中等抗寒性的冬小麦线粒体结构均未发生变化,而不抗冻的材料线粒体膨胀变圆,内嵴遭破坏。一些研究表明,在低温寒害中,线粒体比叶绿体表现出明显的相对稳定,当叶绿体的片层排列方向已强烈改变、类囊体产生空泡时,线粒体尚保持正常,或只表现轻度膨胀;当叶绿体解体时,线粒体内嵴虽遭破坏,但仍维持其个体完整性,甚至细胞已被冻害致死时,其线粒体仍保持很强的呼吸活性[17]。本试验发现,东农冬麦1号在5℃和-15℃低温下的叶绿体及线粒体结构变化不很明显,而济麦22在-15℃低温下,其类囊体基粒片层排列明显发生混乱,并且出现了线粒体的嵴消失现象,这说明济麦22在-15℃低温下其线粒体受到的破坏比叶绿体严重,与前人的研究结果不尽一致。而且Nir等曾报道[18],玉米根尖在干燥条件下因失水也会导致电镜观察线粒体膨胀、变圆、内嵴消失;但线粒体细胞色素氧化酶的细胞化学反应定位发现,电镜下能够清楚地观察到内嵴的存在,其原因是干旱使线粒体内膜和内嵴膜的膜脂和蛋白质的物理化学性质发生了改变,前者用普通的戊二醛-锇酸双固定不能被染色,但内膜和嵴上的细胞色素氧化酶并没有因干旱失水而失活,因此,经细胞化学反应处理后,在电镜下又可看到了嵴的结构。因此,用叶绿体及线粒体结构变化来鉴别分析冬小麦抗寒性机制时,需综合考虑多方面因素。
[1]黄义江,王宗清.苹果属果树抗寒性的细胞学鉴定[J].园艺学报,1982,9(3)∶23-30.
[2]刘艳,李晓燕,王丽雪,等.苹果枝条冬季淀粉粒动态变化与抗寒力的关系[J].内蒙古农业大学学报∶自然科学版,2006,27(2)∶79-83.
[3]刘世彪,陈菁,胡正海.7种番荔枝果树的叶片结构及其与抗寒性关系研究[J].果树学报,2004,21(3)∶241-246.
[4]卢存福,陈玉珍,简令成,等.高山植物唐古特红景天粘液细胞及叶肉细胞表面糖蛋白与抗冻性的关系[J].应用与环境生物学报,2003,9(1)∶16-20.
[5]高述民,程朋军,郭惠红,等.日本桃叶珊瑚的冷驯化及抗寒机制研究[J].西北植物学报,2003,23(12)∶2113-2119.
[6]张有福,陈银萍,张满效,等.两种圆柏属植物不同季节显微和超微结构变化与耐寒性的关系[J].应用生态学报,2006,17(8)∶1393-1397.
[7]董合铸,孙龙华,简令成.不同抗寒性小麦品种的麦苗在冰冻-化冻后叶片细胞亚显微结构的变化[J].植物学报,1980(4)∶339-342.
[8]魏先荣,王国泽.梨组织显微特性与抗寒力的关系研究[J].内蒙古农业大学学报∶自然科学版,2004,25(2)∶73-75.
[9]简令成,孙德兰,施国雄,等.不同柑桔种类叶片组织的细胞结构与抗寒性的关系[J].园艺学报,1986,13(3)∶163-167.
[10]姚胜蕊,曾骧,简令成.桃花芽越冬过程中多糖积累和质壁分离动态与品种抗寒性的关系[J].果树学报,1991,18(1)∶16-20.
[11]Kivimaeenpaeae M,Joensson A M,Stjernquist I,et al.The use of light and electronmicroscopy to assess the impact of ozone on Norway spruce needles[J].Environ Pollut,2004,127∶441-453.
[12]Kivimaeenpaeae M,Selldén G,Sutinen S.Ozone induced changes in the chlorop last structure of conifer needles,and their use in ozone diagnostics[J].Environ Pollut,2005,137∶466-475.
[13]Mai A J.Fine structure of the ray parechyma cells in Populus tremuloides in relation to senscence and seasonal changes[J].Tex J Sci,1979,24∶245-260.
[14]苗芳,冯佰俐,周春菊,等.冷型小麦叶片显微结构的一些特征[J].作物学报,2003,29(1)∶155-156.
[15]苗芳,张嵩午,王长发,等.小麦低温种质的器官结构特征[J].西北植物学报,2005,25(8)∶1499-1507.
[16]郭修武,傅望衡,王光洁.葡萄根系抗寒性的研究[J].园艺学报,1989,16(1)∶17-22.
[17]简令成,王红.逆境植物细胞生物学[M].北京∶科学出版社,2009∶180.
[18]Nir I,Polj ak off-Meyber A.The effects of water stress on mitochondria of root cells[J].Plant Physiology,1970,45∶173-177.
猜你喜欢
耕作与栽培(2022年4期)2022-10-31
蔬菜(2022年12期)2022-01-01
星星·散文诗(2021年34期)2021-12-21
中国马铃薯(2019年1期)2019-03-05
农村百事通(2019年1期)2019-01-30
农家顾问(2014年11期)2014-12-01
中国马铃薯(2013年4期)2013-04-13
中国蔬菜(2011年16期)2011-05-21
