
用于研究快速扫视对瞳孔光反射调制作用的新方法
2010-12-25 06:40陈祖跃胡英周张玉华姜慧慧胡新天
Zoological Research 2010年3期
陈祖跃,胡英周,张玉华,姜慧慧,胡新天,*
(1. 中国科学院昆明动物研究所 灵长类感觉、运动及整合实验室,云南 昆明 650223;2. 中国科学院研究生院, 北京 100049;3. 中国科学院生物物理研究所 脑与认知国家重点实验室,北京 100101)
用于研究快速扫视对瞳孔光反射调制作用的新方法
陈祖跃1,2,胡英周1,2,张玉华1,3,姜慧慧1,2,胡新天1,3,*
(1. 中国科学院昆明动物研究所 灵长类感觉、运动及整合实验室,云南 昆明 650223;2. 中国科学院研究生院, 北京 100049;3. 中国科学院生物物理研究所 脑与认知国家重点实验室,北京 100101)
瞳孔与光反射系统和快速扫视系统在解剖学和功能上都有着紧密联系,但是快速扫视系统对瞳孔的光反射系统是否有调制作用尚无报道。研究这两个系统间的调制作用,必须了解光刺激不均匀和近反应对瞳孔直径变化是否有影响。该研究以人为被试,设计了一种全新的实验方法,研究光刺激不均匀和近反应对瞳孔直径变化的影响。实验方法:将被试的一只眼用密闭的眼罩罩住给予脉冲光刺激,刺激由位于眼罩内全视野范围水平排列的一排发光二极管(light emitting diodes,LEDs)给出,被试的另一只眼用来记录眼动和瞳孔直径的变化,研究水平方向的快速扫视对瞳孔对光反射时瞳孔直径变化的影响。实验结果:比较被试注视视野内不同位置的瞳孔对光反射相对收缩率无显著差异(P=0.148, 非配对样本t检验)。结论:该方法消除了光刺激不均匀和近反应对瞳孔直径变化的影响,可用于研究快速扫视系统对瞳孔光反射系统间的调制作用。
快速扫视;瞳孔对光反射;调制
快速扫视(saccade)是一种常见的视线转移运动(Fuchs et al, 1985; Carpenter, 1991),瞳孔对光反射(pupillary light reflex, PLR)是由于进入视网膜的光亮度的变化而引起瞳孔的直径改变(Yoshitomi & Ito, 1986; Wilhelm, 2008)。快速扫视系统和瞳孔对光反射系统间存在着紧密的解剖学和功能上的联系(Kawamura et al, 1974; Akert et al, 1980; Burde & Loewy, 1980; Gamlin et al, 1995),这两者之间是否存在调制关系尚未见报道。研究它们之间的调制关系不仅有助于进一步了解这两个系统的神经通路和系统间的作用方式,而且对了解脑内其它系统的相互作用并最终了解脑的工作机理有重要意义。
研究快速扫视系统对瞳孔光反射系统的调制作用的最大的困难在于实验方法,方法中所用的光刺激不能影响瞳孔大小。光刺激方式会影响瞳孔大小的变化,视野内不同位置的光刺激引起的瞳孔大小变化也不同:相同亮度的环境光刺激视网膜黄斑外的区域时引起的瞳孔对光反应的阈值要比刺激黄斑区时引起瞳孔对光反应的阈值要低;刺激黄斑区引起的瞳孔对光反射的幅度要比刺激黄斑区外引起的瞳孔对光反射的幅度要大(Pong & Fuchs, 2000; Clarke et al, 2003)。消除这种影响的办法是保证在实验过程中进入黄斑区和黄斑区外的光是均匀的;其次是近反应(the near response)也会影响瞳孔的大小 (Suzuki, 2007):当被试注视一个由远到近的视觉目标时,瞳孔会发生收缩,反之瞳孔会增大,说明瞳孔大小与注视目标的距离远近有关(Myers & Stark, 1990),所以在设计实验时要使所有刺激光点距离眼球的距离相等。
快速扫视是一种双眼协同的联合运动;瞳孔对光反射具有互感性,即光刺激一只眼可以引起双眼的瞳孔收缩。根据这两种特性,我们设计了一种新的实验方法,拟用于研究水平方向的快速扫视对瞳孔光反射是否有调制作用。本研究验证了这种方法是否能有效地排除上述光刺激不均匀和近反应对瞳孔大小的影响,为此比较了被试注视视野内鼻侧10°和颞侧10°时瞳孔对光反射的瞳孔收缩相对率。
1 方 法
1.1 被 试
参与此实验的被试5人,年龄20—30岁之间,平均年龄25.4岁。被试视力或矫正视力正常,能够清晰地分辨屏幕上视角为1°大小的刺激点的位置。实验时被试的精神状态良好。在安静环境中完成此实验。
1.2 仪器和设备
本实验所使用的记录装置为美国ASL公司(Applied Science Laboratories, U.S.A.)生产的红外眼动仪(型号:RS6-HS),采样率为120 Hz。
1.3 刺激装置
刺激是通过戴在一只眼睛上的眼罩内的发光二极管(light emitting diodes,LED)给出,另外一只眼用来记录瞳孔直径大小和视线的位置(图1)。眼罩为半球形,眼罩的直径为10 cm。为了保证在水平方向上进入视网膜外周和黄斑区的光刺激强度一致,在眼罩的内面焊接有12个亮度相对比较均匀的LED:在相距5 cm时,在3 V固定电压下用照度计测量单个LED亮度为(42.13±1.76) lx,相对变化率小于4%。这12个LED沿眼罩内面呈弧形水平排列,每个LED距眼球的距离相等,以减少近反应对瞳孔大小的影响。通过与刺激计算机并口相连的放大电路给LED供电。所给刺激为脉冲光,每个脉冲光刺激持续时间为100 ms。脉冲刺激之间的间隔时间在3—5 s范围内随机。从供电的计算机并口分出一条数据线与记录数据的计算机相连,将刺激信号导入到记录文件中。
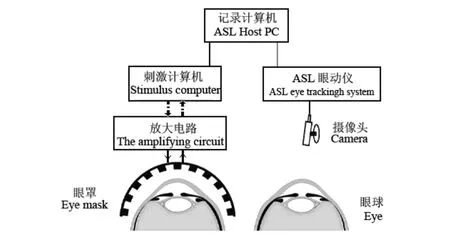
图 1 刺激呈现方式和记录方法示意图Fig. 1 A diagram illustrating the way of stimulus presentation and the recording method
1.4 数据采集
被试下颌置于固定的下巴托上,以保持头部固定。前方约42 cm处放置红外摄像头记录其眼球的运动和瞳孔的直径(图1)。数据由ASL眼动仪器自带的采集软件采集。数据中包含有记录时间、刺激信号、眼球水平位置、垂直位置和瞳孔大小等信息。处理和分析在Matlab7.6下完成。
1.5 眼睛注视位置校准和数据处理
在分析前需要对实验中所涉及的眼睛注视位置和瞳孔直径两个变量进行校准和处理。
为了将记录的相对位置转化为绝对位置,采用9点法对眼睛注视位置进行校准:在被试前方171 cm处放置一个屏幕,上面有3×3个视角大小为1°的点,相邻两点的水平间距为46 cm,视角为15°,垂直间距为25.5 cm,视角为8.5°。实验前让被试在明亮环境下依次注视这9个点,并记录下被试注视每个点时的坐标。根据这9个点位置关系可将实验中记录到的被试的注视点位置转换为绝对位置。
由于眼睛注视位置变化曲线会产生波动。为了确定在瞳孔对光反射过程中是否有快速扫视发生,首先对眼睛注视位置变化曲线进行滤波,其方法是采用五点中央平均滤波器(a five-point central averaging filter)滤波。滤波后信号乘以采样率后再微分,得出每一时刻的眼动速度。然后从微分后的结果中挑出速度大于或等于30°/s数据点。连续有3个数据点大于或等于30序列,就视为一次快速扫视过程。
1.6 瞳孔直径校准和数据处理
为把瞳孔相对直径转换为绝对直径并验证测量结果的可靠性,需要对瞳孔直径做校准,其校准方法是将模拟瞳孔固定在一个可以沿水平方向旋转的底座上,旋转臂的长近似于人的眼球的平均半径。将底座置于下巴托上,使模拟瞳孔和被试眼球保持在同一位置。模拟瞳孔正视摄像头时的角度为0°。记录模拟瞳孔注视0° 和左右两侧5、10、15、20和25° 的瞳孔直径大小。在黑暗条件下,让被试也注视视野内水平方向上不同位置,记录真实瞳孔注视视野中心和左右两侧不同位置时的瞳孔直径大小。把记录的模拟瞳孔和真实瞳孔直径大小对注视位置作图,得出模拟瞳孔和真实瞳孔直径随注视位置变化关系图(图2)。从图中可以看出,真实瞳孔直径大小随注视位置变化与模拟瞳孔直径的变化趋势不同。在注视点靠近视野中央时两条曲线基本重合,当注视点向视野两侧移动时,两条曲线逐渐发生分离。这说明实验中可能有不明因素造成了被试瞳孔大小的变化。为减少这种影响,将挑出的快速扫视范围限定在视野中心两侧10° 以内。在这个范围内,在5个对应注视位置对每名被试的瞳孔直径大小的分布与模拟瞳孔直径进行逐个比较,模拟瞳孔直径大小始终落在被试瞳孔直径大小分布的71%以内。这说明在这个范围可以认为被试的瞳孔直径变化和模拟瞳孔是一致的。
模拟瞳孔的大小是固定的,但是记录出来的模拟瞳孔直径大小却是随注视点变化的,其变化趋势是一条近似的二次函数曲线。这是因为当模拟瞳孔注视视野中心时,瞳孔在摄像头上的投影是圆形,记录的瞳孔直径是圆的直径(R2);而在注视视野两侧时,瞳孔在摄像头上的投影是一个椭圆,记录的直径就相当于椭圆短轴长的2倍(R1, R3)(图3)。为减少注视位置对瞳孔测量值的影响,需要将注视视野两侧时的直径测量值转化为注视视野中心时的直径大小。具体方法是:先对模拟瞳孔直径变化曲线进行二次拟合,根据这种函数变化关系将真实瞳孔注视视野两侧时的测量值转化为注视视野中心时的直径大小。

图 2 被试瞳孔和模拟瞳孔在注视水平不同位置时瞳孔直径大小比较Fig. 2 The relationship between the pupil diameters of eyes/model pupil and fixation positions
为了减少瞳孔括约肌震颤和仪器不稳定性造成的瞳孔直径大小的波动,对上述处理后的被试瞳孔直径采用低通滤波,滤去频率为10 Hz以上的成分。滤波后的瞳孔信号用于进一步的统计和分析瞳孔的收缩状况。
由于不同个体的瞳孔直径不同,用相对收缩率(relative constriction ratio)来衡量瞳孔收缩状况。收缩前的瞳孔直径为刺激前10个数据点(约83 ms)、刺激呈现时刻、刺激呈现后10个数据点(约83 ms)的瞳孔直径的平均值。瞳孔的收缩幅度为收缩前的瞳孔直径和收缩后的瞳孔直径的最小值之差。相对收缩率等于收缩幅度除以收缩前的瞳孔直径乘以100%后的值。
1.7 方法有效性验证
为了验证光刺激是否均匀,以及在±10° 内眼睛位置是否对瞳孔的对光反射有影响,我们设计了以下实验:实验前在明亮环境下,在被试视野中心鼻侧和颞侧10° 的两个点呈现光刺激,让被试记住两个点。然后在黑暗条件下,让被试在这两个点间随意做快速扫视,记录被试注视这两个点时的瞳孔对光反射。每次实验做7 min,前2 min不给光刺激,只让被试在两点间做快速扫视,后5 min为光刺激条件下的实验,通过眼罩给光刺激。两次实验之间让被试休息5 min。每天完成4次实验(左右眼各两次实验),每个被试做5 d。然后比较被试注视视野左右两侧时的瞳孔相对收缩率。如果我们所用的光刺激是不均匀的或单个的LED距眼球距离不等,那么被试在注视视野两侧时视网膜接受的光刺激是不一样的,就会造成瞳孔对光反射的差异;如果光刺激是均匀的或LED距眼球距离相等,那么就不会造成瞳孔对光反射的差异。
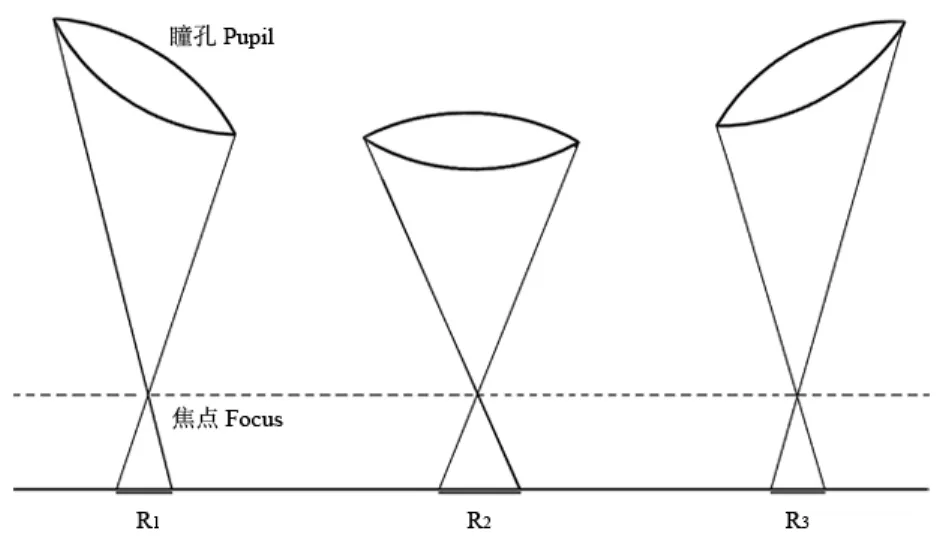
图 3 模拟瞳孔注视视野内不同位置时,摄像头所测得的瞳孔大小不同Fig. 3 A diagram illustrating the difference of the model pupil diameter when the subject fixated at different positions in visual field
根据刺激将每次实验分成若干次瞳孔反应:以刺激呈现时刻为标记点,一次瞳孔反应的时间为刺激前160 ms到刺激呈现后2 000 ms的时间。统计每次瞳孔反应时的瞳孔直径相对收缩率。实验结果用SPSS软件包(SPSS 13.0 for windows)进行分析,注视视野两侧时的瞳孔对光反射的相对收缩率的比较采用非配对样本t检验。
2 结 果
在有效性实验验证中,我们将鼻侧和颞侧10°的两个点的窗口设为2°,即当被试的视线在一个瞳孔对光反射周期内始终停留在(10±2)° 范围内,并且没有快速扫视出现,就认定为一次注视状态下的瞳孔对光反射。从5个被试的7 500次的瞳孔反应中,我们挑出了符合条件的71次注视颞侧视野时瞳孔对光反射的瞳孔反应,83次注视鼻侧视野时瞳孔对光反射的瞳孔反应。统计每位被试在注视鼻侧和颞侧10° 时瞳孔相对收缩率发现,注视颞侧视野时瞳孔对光反射的相对收缩率为(18.04±0.43)%,注视鼻侧视野时瞳孔对光反射的相对收缩率为(18.14±0.44)%,瞳孔相对收缩率并没有表现出显著差异(P=0.148, 非配对样本t检验)。表明所用的光刺激是均匀的,LED距眼球距离是相等的。
3 讨 论
调制作用是神经系统中一种普遍的作用方式(Shors et al, 2000; Nunez & Malmierca, 2007)。瞳孔光反射回路位于脑干,快速扫视也可以由其脑干中枢独立完成,研究瞳孔光反射系统和快速扫视系统间调制作用有助于解释皮层以外的调制作用是如何发生的,但在研究方法上要求排除与眼动有关的一些因素对瞳孔大小的影响。本方法所用的光刺激是均匀的,且LED距眼球距离相等,可满足此要求。首先,用于给光刺激的LED亮度差异较小,而且是均匀整齐地排列在眼罩内,保证了进入黄斑区和黄斑区外的光的分布是均匀的。LED所在球面与眼球共球心,保证了每个LED与眼球的距离是一样的,从而排除近反应对实验结果的干扰。为了验证本方法排除上述干扰因素影响的有效性,我们比较了5名被试注视视野内鼻侧和颞侧距中心10°时的瞳孔对光反射情况,结果显示注视两侧视野时瞳孔相对收缩率并无显著差异,这说明我们的方法是可行的。
比较模拟瞳孔和真实瞳孔发现,在没有快速扫视的情况下,瞳孔直径随注视位置变化趋势是不一样的。这说明环境中有不明因素造成了被试瞳孔直径的变化,这种因素可能是来自摄像头的红外光源引起了瞳孔的变化,被试的心理状态可能也会影响实验的结果。为了减少这种影响,从实验中选取的被试快速扫视范围限定在视野中心±10° 内。在这个范围内,模拟瞳孔直径大小始终落在被试瞳孔直径大小分布的71%以内,可以忽略被试和模拟瞳孔的直径变化差异。另外当注视点偏离视野中心越大,眼动仪器的稳定性越差,丢失的数据也就越多,限定快速扫视的范围可以增加有效实验的次数。
我们采用这种方法记录了5名被试的瞳孔对光反射,从记录结果中能够提取快速扫视的方向、一个瞳孔对光反射周期内出现快速扫视的次数、快速扫视的幅度、快速扫视在一个瞳孔对光反射周期内出现的时间等信息,在此基础上可进一步研究两个系统间的调制作用。改变眼罩内LED的排列方式可以研究其他方向上的快速扫视对瞳孔对光反射的调制作用。
Akert K, Glicksman MA, Lang W, Grob P, Huber A. 1980. The Edinger-Westphal nucleus in the monkey. A retrograde tracer study [J].Brain Res, 184(2): 491-498.
Burde RM, Loewy AD. 1980. Central origin of oculomotor parasympathetic neurons in the monkey [J].Brain Res, 198(2): 434-439.
Carpenter RHS. 1991. Eye Movements [M]. London: Macmillan Press Ltd.
Clarke RJ, Zhang H, Gamlin PD. 2003. Characteristics of the pupillary light reflex in the alert rhesus monkey [J].J Neurophysiol, 89(6): 3179-3189.
Fuchs AF, Kaneko CR, Scudder CA. 1985. Brainstem control of saccadic eye movements [J].Annu Rev Neurosci, 8: 307-337.
Gamlin PD, Zhang H, Clarke RJ. 1995. Luminance neurons in the pretectal olivary nucleus mediate the pupillary light reflex in the rhesus monkey [J].Exp Brain Res, 106(1): 169-176.
Kawamura S, Sprague JM, Niimi K. 1974. Corticofugal projections from the visual cortices to the thalamus, pretectum and superior colliculus in the cat [J].J Comp Neurol, 158(3): 339-362.
Myers GA, Stark L. 1990. Topology of the near response triad [J].Ophthalmic Physiol Opt, 10(2): 175-181.
Nunez A, Malmierca E. 2007. Corticofugal Modulation of Sensory Information: Advances in Anatomy, Embryology and Cell Biology 187[M]. New York: Springer.
Pong M, Fuchs AF. 2000. Characteristics of the pupillary light reflex in the macaque monkey: discharge patterns of pretectal neurons [J].J Neurophysiol, 84(2): 964-974.
Shors TJ, Beylin AV, Wood GE, Gould E. 2000. The modulation of Pavlovian memory[J].Behav Brain Re, 110(1-2): 39-52.
Suzuki Y. 2007. The near response: the contributions of Kenji Ohtsuka, MD [J].J Neuroophthalmol, 27(2): 138-142.
Wilhelm H. 2008. The pupil [J].Curr Opin Neurol, 21(1): 36-42.
Yoshitomi T, Ito Y. 1986. Double reciprocal innervations in dog iris sphincter and dilator muscles [J].Invest Ophthalmol Vis Sci, 27(1): 83-91.
A New Method to Explore the Modulation from the Saccadic System on the Pupillary Light Reflex System
CHEN Zu-Yue1,2, HU Ying-Zhou1,2, ZHANG Yu-Hua1,3, JIANG Hui-Hui1,2, HU Xin-Tian1,3,*
(1.Laboratory of Sensory-motor Integration Research, Kunming Institute of Zoology, the Chinese Academy of Sciences, Kunming650223,China;
2.Graduate University of the Chinese Academy of Sciences, Beijing100049,China;
3.State Key Laboratory of Brain and Cognitive Science, Institute of Biophysics, the Chinese Academy of Sciences, Beijing100101,China)
The saccadic system has anatomical and functional connections with the pupillary light reflex (PLR) system. But it is not known whether the saccadic system modulates the PLR system. To investigate this issue, it is necessary to understand whether the uneven light stimulus to retina and the near responses influence the change of pupil diameter. We designed a new behavioral method to investigate the issue on human subjects. Methods: one eye of the subject was stimulated by pulse light stimulus from a horizontal linear array of light emitting diodes (LEDs) presented across visual field in an ocular mask. The changes of the eye position and pupil diameter of another eye were recorded by an infrared eye tracking system. Results: The relative constriction ratios of PLRs on the condition that the fixation points of subjects were in the nasal visual field were not significantly different from that the fixation points of subjects were in the temporal visual field(P=0.148, non-pairedttest).Conclusion: The influences from uneven light stimulus to retina and the near responses were eliminated by this method. The method can be used to study the modulation from the saccadic system on the pupillary light reflex system.
Saccade; Pupillary light reflex (PLR); Modulation
R339.143;Q436
A
0254-5853-(2010)03-0287-05
10.3724/SP.J.1141.2010.03287
2009-11-12;接受日期:2010-05-10
CASC项目(YZ200737)
*通讯作者(Corresponding author),E-mail:xthu@mail.kiz.ac.cn
E-mail:chenzuyue@gmail.com
猜你喜欢
纺织标准与质量(2022年2期)2022-07-12
纺织标准与质量(2022年2期)2022-07-12
上海塑料(2021年3期)2022-01-06
扬子江(2020年4期)2020-08-04
婚姻与家庭·性情读本(2019年11期)2019-11-18
中国眼镜科技杂志(2019年5期)2019-07-13
陶瓷(2018年11期)2018-12-17
青年歌声(2018年2期)2018-10-20
小朋友·快乐手工(2017年10期)2018-01-05
阅读与作文(初中版)(2017年6期)2017-07-05
