
垂穗披碱草种子的萌发适宜温度及温度阈值
2011-04-25 09:42王彦荣胡小文
草业科学 2011年6期
曾 霞,王彦荣,胡小文
(兰州大学草地农业科技学院,甘肃 兰州 730020)
垂穗披碱草(Elymusnutans)是禾本科(Gramineae)小麦族(Triticeae)披碱草属(Elymus)多年生根茎疏丛型草本植物,广泛分布于我国内蒙古、河北、陕西、甘肃、宁夏、青海、新疆、四川和西藏等省(区),拥有麦类作物所缺乏的优质、高产、抗病、抗虫、抗旱、耐盐碱等优良基因,是麦类作物的野生近缘种遗传资源,具有重要的经济利用价值[1]。其在花期前是质地柔软、无刺毛、刚毛、无异味的优质牧草[2-3],抗旱和抗寒能力强、适口性好,可用于建植栽培草地和放牧草地,是高寒地区退化草地补播和改良用主要草种。近年来,随着国家“西部大开发”、“退耕还草”和“生态补偿”等政策的有效实施,优质牧草种子的需求量越来越大。但我国牧草检验规程中披碱草属草种子的检验方法只有老芒麦(E.sibiricus)和披碱草(E.dahuricus)的,尚无垂穗披碱草的检验标准。目前,垂穗披碱草的种子产量、生产性能、耐盐性及放牧利用等方面的研究有报道[4-12],但关于其适宜萌发温度和检验方法的研究鲜见报道。为此,本研究对采集的不同垂穗披碱草种质材料进行种子萌发对温度响应的试验,旨在探讨垂穗披碱草种子的适宜萌发温度和萌发温度阈值,为种子检验和田间栽培利用提供参考。
1 材料与方法
1.1试验材料 供试的垂穗披碱草种子于2008年9月采收自甘肃和青海两地,采集地的海拔、温度、降水量等信息见表1。种子均成熟饱满,风干后置于兰州大学草地农业科技学院种子冷库(4 ℃)保存待用。发芽试验于2009年4-8月在农业部牧草与草坪草种子质量监督检验测试中心(兰州)进行。
1.2试验方法 试验设5、10、15、20、25、30、35和40 ℃ 8个恒温处理,依据国家《牧草种子检验规程》[13]的相关规定,对供试材料进行不同温度条件下的萌发试验。发芽率测定采用纸上发芽法,每温度处理50粒种子,4次重复。将种子分别置于设定的各温度条件下在培养箱发芽,每隔12 h统计发芽种子数,以胚根长度超过2 mm视为发芽,连续4 d没有新的发芽种子后计算最终发芽率。发芽指数(GI)参照毕辛华[14]的方法计算,即:
GI=∑GT/DT。
式中,GT为第T天发芽数,DT为天数。
1.3种子萌发的温度模型 采用种子萌发速率与平均温度(T,℃)模型[15-16](1/tg=k+mT)计算垂穗披碱草的最低临界温度(Tb)、最适宜萌发温度(To)和最高临界温度(Tc);1/tg为累计萌发率达g%时所需时间(天数)的倒数,T为萌发设定的温度,k和m是常数。方程和横坐标的交点就是萌发的Tb和Tc,两条线的交点是To,Tb、To和Tc被称为种子萌发的温度三基点。

表1 供试垂穗披碱草种子的来源及其相关信息
1.4数据处理与统计 所有数据均用Microsoft Office 2003 Excel录入并作图,采用SPSS 17.0软件进行最小极差法(LSD)多重比较。
2 结果
2.1垂穗披碱草种子在不同温度下的发芽率和发芽速度 发芽率随处理温度的升高,所有种质表现出一致的趋势,先升高后降低,但各指标间存在一定的差异。其中,15、20、25和30 ℃温度处理的发芽率显著高于其他温度处理(P<0.05),且这几个温度间无显著差异(P>0.05),平均发芽率达95.62%;发芽指数也呈现相同的趋势,在20、25和30 ℃温度处理时最高,且这几个温度间无显著差异(P>0.05),平均发芽指数达到17.95,其他温度条件下因种质不同而存在一定差异(表2)。
供试垂穗披碱草各个种质之间无显著差异(P>0.05),按其平均值做出种子发芽动态图(图1)。

表2 不同温度下垂穗披碱草的发芽率和发芽指数
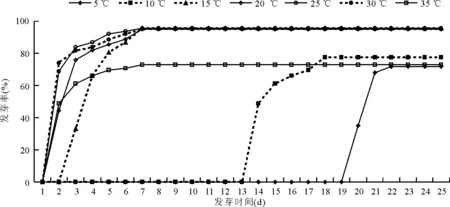
图1 在不同温度下供试垂穗披碱草种质的平均发芽动态
在5 ℃温度处理时,种子第19天开始正常发芽,第20天达到发芽高峰,最终发芽率为72%;在10 ℃温度处理时,种子在第13天开始正常发芽,到第14天达到发芽高峰;在15 ℃温度处理时,种子在第2天开始正常发芽,第3天达到发芽高峰;而在20、25、30和35 ℃ 4个温度下,种子在第1天开始萌发,第2天达到发芽高峰,最终发芽率在95%左右。但随着温度的继续增加,种子的最终萌发率与萌发速率迅速下降,如35 ℃下最终发芽率仅为73%,40 ℃下均不萌发(图1)。
2.2垂穗披碱草种子的萌发温度阈值 5和10 ℃低温下的发芽率与采集地的海拔、年均温、年降水量之间无显著相关关系(P>0.05),各个采集地种子在5 ℃的发芽率和10 ℃的发芽率呈极显著正相关(P<0.01),采集地的海拔和年均温之间呈显著负相关(P<0.05)(表3)。
不同垂穗披碱草种质的萌发温度三基点Tb、To和Tc间无显著差异(P>0.05),Tb在3.8~4.7 ℃,To在22.6~24.5 ℃,而Tc在38.0~38.2 ℃(表4)。

表3 低温处理下的发芽率与采集地因素的相关性
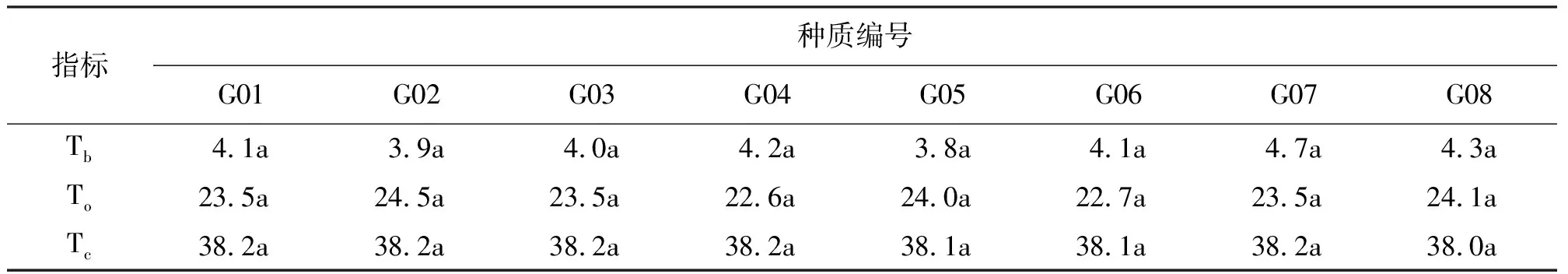
表4 8份垂穗披碱草种质的萌发温度三基点 ℃
由于8个种质的萌发温度三基点无显著差异(P>0.05),用其平均值做出温度对垂穗披碱草萌发速率的影响(图2)。其中,种子的萌发速率在5~25 ℃时与温度呈极显著的直线正相关(P<0.01),相关系数r=0.966 6,两者关系可以用方程y1=0.039 3x1-0.161 8表示;种子的萌发速率在25~40 ℃时与温度极显著的直线负相关(P<0.01),相关系数r=0.935 1,两者关系可以用方程y2=-0.052 1x2+1.987 8表示。根据萌发速率(1/t50)和温度之间的关系确定萌发温度三基点(图2),得到垂穗披碱草的Tb为4 ℃,To为24 ℃,Tc为38 ℃。

图2 温度对垂穗披碱草萌发速率的影响
3 讨论与结论
种子萌发是一个非常活跃的生理生化过程,是在一系列酶的参与下进行的,而酶的催化与温度有密切关系[17],因此,适温时种子才能表现最高的发芽率和发芽速率。蒋小军和文向多[18]对蓖麻(Ricinuscommunis)的研究发现,35 ℃高温降低了发芽率,提早了发芽时间。该结论在玉米(Zeamays)[19]、长鞭红景天(Rhodiolafastigiata)[20]和多种禾本科牧草种子[21]中得到了证实,本研究也得到了类似结果。不同植物种子萌发特性不同[22],且同一物种不同种群的种子萌发行为也具有较大变异[23]。多数植物种子在10 ℃以下萌发不启动或启动极为缓慢,因此,常将10 ℃作为种子的临界萌发温度,也有将10 ℃作为种子受冷害的温度[24]。本研究中,垂穗披碱草种子在5和10 ℃下具有70%左右的发芽率,表明垂穗披碱草种子萌发较能耐受低温。而在温度为5~35 ℃时,种子均能萌发,可见其具有较宽的萌发温幅,这也可能是其能广泛分布的原因之一。
大量研究表明,植物种子的萌发速率在萌发的最低临界温度至最适温度之间与温度呈极显著的直线正相关,在萌发的最适温度到最高临界温度之间与温度极显著的直线负相关,可表示为下列方程式:1/tg=k+mT,萌发速率为0时,-k/m就是萌发最低临界温度值,对于一个种批或者一个物种来说萌发最低临界温度是一个定值[15-16,25],该结论在本试验中也得到证实,与檞果(Quercusrobur)[26],无芒隐子草(Cleistogenessongorica)、多裂骆驼蓬(Peganummultisectum)和雾冰藜(Bassiadasyphylla)[27]研究一致,且通过计算得出它的Tb、To、Tc值分别为4、24和38 ℃,而黄文达等[28]研究的无芒隐子草的Tb、To、Tc分别为6、23和43 ℃,多裂骆驼蓬分别为21、34和48 ℃,雾滨黎分别为3、21和42 ℃。由此可见,不同物种的萌发温度的阈值是不同的,这与Labouriau[15]和Dahal等[16]的研究结论相同。
根据本研究结果,建议垂穗披碱草种子的适宜萌发温度为20、25和30 ℃恒温,初次和末次的适宜统计时间分别为第3天和第11天,本研究可为国家牧草种子检验规程发芽试验(GB/T2930.4-2001)的修订提供关于垂穗披碱草种子的发芽方法依据。
[1]卢宝荣.披碱草属与大麦属系统关系的研究[J].植物分类学报,1997,35(3):193-207.
[2]陈默君,贾慎修.中国饲用植物[M].北京:中国农业出版社,2002:119-120.
[3]周永红,郑有良.10种披碱草属植物的RAPD分析及其系统学意义[J].植物分类学报,1999,37(5):425-432.
[4]张金旭,马玉寿,施建军,等.刈割对江河源区混播草地牧草产量及品质的影响[J].草业科学,2010,27(1):92-96.
[5]董文斌,马玉寿,董全民,等.退耕还草多年生草地地上生物量及牧草营养成分研究[J].草业科学,2010,27(2):54-58.
[6]乔安海,韩建国,巩爱岐,等.氮肥对垂穗披碱草种子产量和质量的影响[J].草地学报,2006,14(1):48-56.
[7]敖特根白音,姬振亭.杂种披碱草及亲本种子萌发期抗逆性比较试验[J].华南农业大学学报,2004,25(增刊):28-30.
[8]谢国平,呼应明,王佺珍,等.施N量和收获时间对西藏野生垂穗披碱草种子产量影响研究[J].草业学报,2010,19(2):89-96.
[9]杜玉红,周学丽,王建峰,等.高寒地区4种禾草科牧草的生产特性比较[J].草业科学,2010,27(8):166-168.
[10]严学兵,汪玺,郭玉霞,等.高寒牧区垂穗披碱草草地生物量及营养价值动态的研究[J].草业科学,2003,20(11):14-18.
[11]陆光平,聂斌.垂穗披碱草利用价值评价[J].草业科学,2002,19(12):13-15.
[12]盘朝邦,王元富.老芒麦、垂穗披碱草产草量形成与水热季节变化的关系[J].草业科学,1992,9(6):13-17.
[13]王彦荣,孙建华,余玲,等.GB/T 2930.4-2001 牧草种子检验规程 发芽试验[S].北京:中国标准出版社,2001.
[14]毕辛华.种子检验[M].北京:农业出版社,1986:94-118.
[15]Labouriau L G.On the physiology of seed germination inViciagramineaSm[J].Anais da Academia Brasileira de Ciências,1970,42(2):235-262.
[16]Dahal P,Bradford K J,Jones R A.Effect of priming and endosperm integrity on seed germination rates of tomato genotypes[J].Journal of Experimental Botany,1990,41(232):1431-1439.
[17]潘瑞炽,董愚得.植物生理学[M].北京:科学出版社,2005:57-58.
[18]蒋小军,文向多.温度对蓖麻种子萌发的影响[J].种子,2008,27(5):67-69.
[19]谢皓.玉米种子发芽与温度相关性的研究[J].种子,1998,94(2):14-16.
[20]卢杰,郑维列,张建新,等.温度对长鞭红景天种子发芽的影响[J].西北林学院学报,2010,25(3):101-106.
[21]颜红波.不同温度对多年生禾草种子发芽的影响[J].草业科学,1998,15(4):22-26.
[22]Figueroa J A.Seed germination in temperate rain forest species of southern Chile: chilling and gap dependency germination[J].Plant Ecology,2003,166:227-240.
[23]Meyer S E,Allen P S,Beckstead J.Seed germination regulation inBromustectorum(Poaceae) and its ecological significance[J].Oikos,1997,78:475-486.
[24]沈漫,王明庥,黄敏仁.植物抗寒机理研究进展[J].植物学通报,1997,14(2):1-8.
[25]Kebreab E,Murdoch A J.A quantitative model for loss of primary dormancy and induction of secondary in imbibed seeds ofOrobanchespp.[J].Journal of Experimental Botany,1999,50:211-219.
[26]Finval Savage W E.Embryo water status and survival in the recalcitrant speciesQuercusroburL.:Evidence for a critial moisture-content[J].Journal of Experimental Botany,1992,43:663-669.
[27]黄文达,王彦荣,胡小文.三种荒漠植物种子萌发的水热响应[J].草业学报,2009,18(3):171-177.
[28]黄文达.三种荒漠植物种子萌发的水热响应[D].兰州:兰州大学,2007.
猜你喜欢
草业学报(2022年11期)2022-11-23
美文(2022年18期)2022-10-20
青海草业(2022年1期)2022-04-27
现代畜牧科技(2021年7期)2021-07-28
今日农业(2021年10期)2021-07-28
西藏农业科技(2021年4期)2021-04-18
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20
草业科学(2017年1期)2017-03-14
西藏科技(2015年11期)2015-09-26
草业科学(2015年4期)2015-02-16
