
个体基因杂合度对山西白猪体重和体尺性状的影响
2011-04-25 10:17杨晓奋
山西农业大学学报(自然科学版) 2011年3期
杨晓奋
(山西农业大学动物科技学院,山西太谷 030801)
个体基因杂合度是指个体基因组所有基因座位上杂合基因座所占比例,反映个体的遗传杂合程度。一般认为,高杂合度往往会带来高生活力和高生产性能。近年来,许多学者研究了基于微卫星标记的个体基因杂合度对猪生长性状[1]、胴体性状[2]、肉质性状[3,4]以及绵羊的生长性能的影响[5],发现个体基因杂合度水平对不同生产性状的作用方向不一致,且在不同品种或群体间有差异。山西白猪是以马身猪、长白猪、太湖猪为亲本,通过复杂杂交和群体继代选育法育成的瘦肉型专门化母本品系,具有产仔数高,胴体和肉质品质好等特点[6]。本研究分析个体基因杂合度对山西白猪体重和6月龄体尺性状的影响,为该品系的选育提高提供理论依据。
1 材料与方法
1.1 试验猪群与性状测定
试验猪群为山西白猪选育过程中5世代后备猪65头,6世代后备猪63头,均来自山西省大同市种猪场。测定性状包括初生重(birth weight,BW)、断奶重(weaning weight at 28-day age,W28)、6月龄体重(body weight at six-month age,W6M)、体长(body length at six-month age,L6M)、体高(body height at six-month age,H6M)、胸围(chest girth at six-month age,CG6M)和背膘厚(back fat thickness at sixmonth age,BFT)等,其中背膘厚用背膘测定仪(Renco lean-meater,USA)测定,测定部位在猪左侧腰荐结合部距背中线5cm处。性状测定值首先在Excel环境下建立数据库,剔除2倍标准差以外的极端数字后进行统计分析。
1.2 方法
1.2.1 耳组织的采集和DNA的提取
在不影响猪耳号的地方,用耳号钳剪耳组织一小块(大约0.5 g),放于装有1 mL 70%乙醇的1.5 mL的离心管中,带回实验室,-70℃保存备用。采用常规的酚/氯仿抽提法提取基因组DNA。
1.2.2 个体基因杂合度的测定与分类
选用FAO-ISAG联合推荐的21个微卫星标记 (S0155、SW24、S0026、SW936、S0355、SW857、 S0218、S0090、SW122、S0228、SW632、S0101、S0025、S0178、SW951、S0386、S0005、S0227、S0002、SW72 和 SW240)进行个体基因型测定,具体方法见文献[7]。个体基因杂合度根据个体中杂合座位数占总座位数的比例计算。计算时,首先利用SAS系统(v6.12)的GLM过程分析各微卫星座位与体重、体尺性状的关联性,如果有与生产性状显著关联的微卫星座位(表1),则剔除该座位,只以中性座位计算个体基因杂合度。以0.05为步长将所有个体分为6个杂合度水平(表2)。

表1 与体重和体尺性状有显著关联的微卫星座位Table 1 Microsatellite loci significantly associated with the traits of body weight and size
1.2.3 数据处理与统计
利用PopGene Version 1.31软件计算各微卫星座位的等位基因频率、观察杂合度(Observed heterozygosity,HO)和期望杂合度(Expected heterozygosity,HE);利用Cervus 2.0软件计算多态信息含量(Polymorphism Information Content,PIC)。
采用包括母体效应的单变量动物模型分析个体基因杂合度水平对山西白猪体重和体尺性状的影响。对初生重和断奶重(28日龄)配合模型如下:
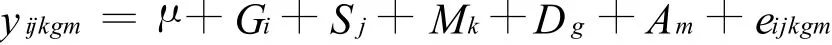
其中,μ为总体均值;yijkgm为第i个世代第j个性别第k种个体基因杂合度水平第g个母亲的第m个个体的观察值;Gi为世代固定效应(i=1,2);Sj为性别效应(j=1,2);Mk为个体基因杂合度(k=1-6);Dg为母体效应(g=1-68)(因运算中所使用的记录均为母猪第一胎所产后代的记录,这里母体效应包括母体直接遗传效应和共同环境效应)为个体加性遗传效应(m=1-155);为随机残差效应。
对6月龄体重、体高、体长、胸围等4个性状,测定性状时个体的年龄作为协变量包括在上述模型中,以校对年龄对性状的影响。对6月龄背膘厚,因体重与背膘厚有相关性[8],分析时,模型中除包括世代、性别、个体基因杂合度水平等固定效应外,还包括测定日龄和6月龄体重等协变量。
采用WOMBAT软件的单变量分析程序进行分析[9]。数据分析时,首先在SAS系统(v6.12)的VARCOPM 程序下,采用REML方法估计各性状的表型方差、母体遗传效应方差、母体永久环境效应方差和误差方差,再结合遗传力计算公式和各性状遗传力经验值[10,11],计算出个体加性遗传方差。将所获得的加性遗传方差、母体遗传效应方差、母体永久环境效应方差和误差方差作为初值,采用平均信息约束最大似然法(AIREML),在WOMBAT软件下进行迭代计算,迭代收敛标准为10-9。采用T检验对不同世代、性别和个体基因杂合度水 平的最小二乘解进行差异显著性检验。

表2 个体基因杂合度分类Table 2 Heterozygosity range(level),observe number and mean heterozygosity classified by 21 microsatellite loci
2 结果与分析
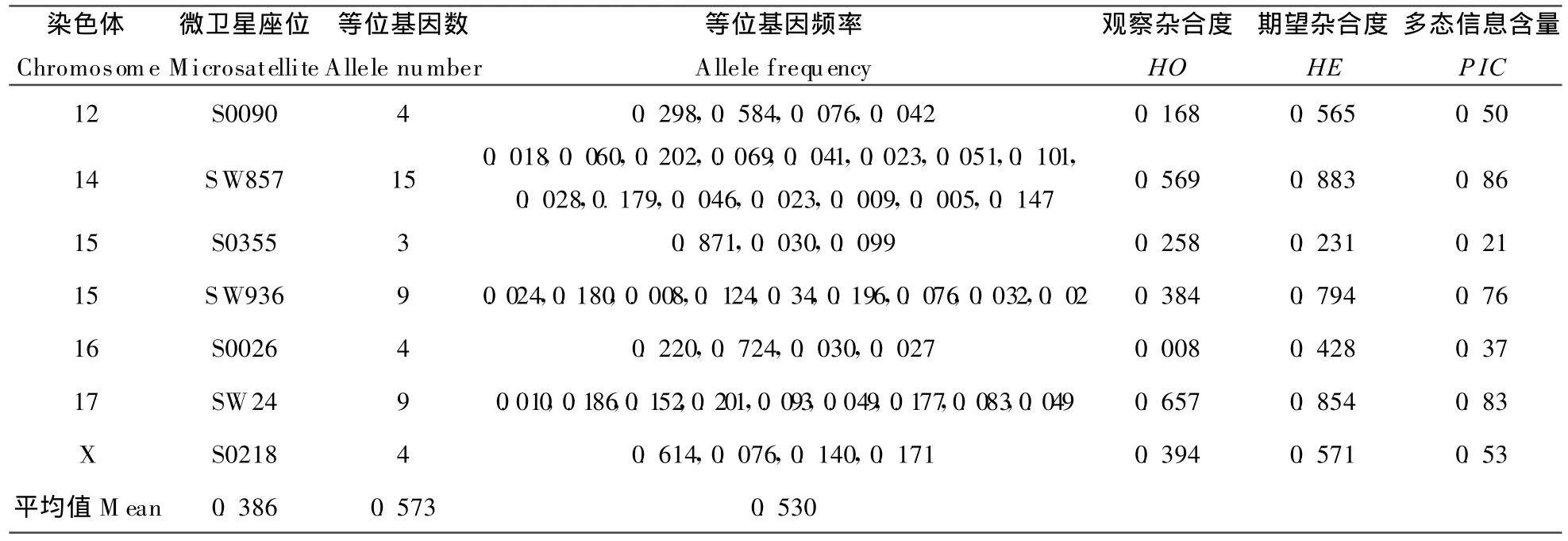
2.1 微卫星座位杂合度
本研究中,1~17号常染色体和X染色体上的21个微卫星座位的杂合度和多态信息含量见表3(资料来源于文献[7])。整个群体平均多态信息含量为 0.530,呈高度多态;平均观察杂合度为0.386,而平均期望杂合度为0.573,高于观察杂合度,主要原因是在某些座位上,个体主要以纯合子形式存在。

表3 微卫星座位的杂合度和多态信息含量Table 3 Heterozygosity and polymorphic information content(PIC)for 21microsatellite loci
染色体Chromosome微卫星座位Microsatellite等位基因数Allele number等位基因频率Allele frequency观察杂合度HO期望杂合度HE多态信息含量PIC 12 S0090 4 0.298,0.584,0.076,0.042 0.168 0.565 0.50 14 SW857 15 0.018,0.060,0.202,0.069,0.041,0.023,0.051,0.101,0.028,0.179,0.046,0.023,0.009,0.005,0.147 0.569 0.883 0.86 15 S0355 3 0.871,0.030,0.099 0.258 0.231 0.21 15 SW936 9 0.024,0.180,0.008,0.124,0.34,0.196,0.076,0.032,0.02 0.384 0.794 0.76 16 S0026 4 0.220,0.724,0.030,0.027 0.008 0.428 0.37 17 SW24 9 0.010,0.186,0.152,0.201,0.093,0.049,0.177,0.083,0.049 0.657 0.854 0.83 X S0218 4 0.614,0.076,0.140,0.171 0.394 0.571 0.53平均值M ean 0.386 0.573 0.530
2.2 个体基因杂合度对体重和体尺性状的影响
不同杂合度水平下各体重和体尺性状的平均数见表4,最小二乘解和标准误见表5。由表可见,个体基因杂合度水平仅对初生重有显著影响(P<0.05),杂合度水平为 6(杂合度为 0.568)时,初生重最高,比杂合度水平为1和2的个体分别高出0.11 kg和0.112 kg,差异显著,而与其它杂合度水平的个体初生重无显著差异(P>0.05)。
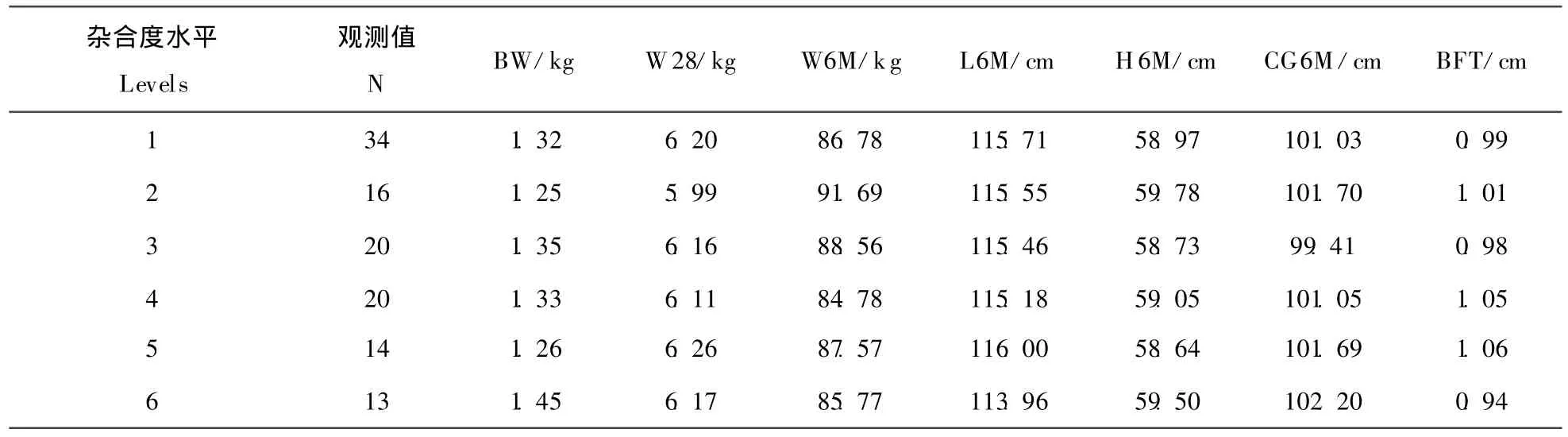
表4 不同杂合度水平体重和体尺性状的平均数Table 4 M ean of body weights and body size at six individual gene heterozygosity levels
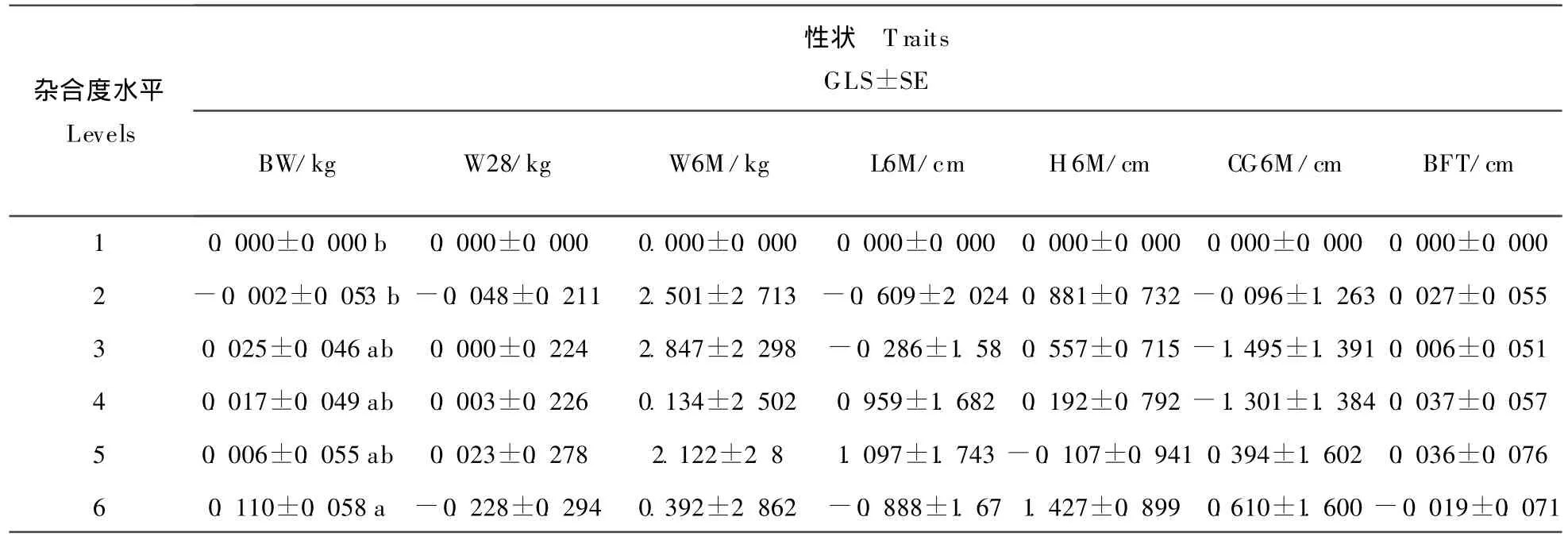
表5 不同杂合度水平体重和体尺性状的最小二乘解和标准误Table 5 General least square solutions(GLS)and standard errors(SE)of body weights and body size of individuals at six individual gene heterozygosity levels
3 讨论与结论
一般认为,高杂合度与近交衰退呈负相关,与杂种优势呈正相关,但基因杂合度与生长性能间不是简单的相关关系[5]。研究表明,个体基因杂合度水平对部分生长性状[1]、胴体性状[2]、肉质性状[3,4]及泌乳性能[12]有显著影响。个体平均杂合度在0.5796时,猪180日龄活重、日增重、相对生长率等指标最高,之后随着个体基因杂合度的增加而降低[1]。个体基因杂合度小于0.6时,皮率与骨率随个体基因杂合度增加而降低;个体基因杂合度超过0.6时,骨率和皮率随个体基因杂合度的增加而增加,眼肌面积随个体基因杂合度的增加而上下波动[2]。个体基因杂合度为0.5353时,肌苷酸含量最高,为4.6531 mg◦g-1,此后随杂合度增加而显著降低[4]。牦牛乳蛋白基因杂合度对产奶量、乳脂率、α-乳清蛋白等有显著影响,随个体基因杂合度的增加,产奶量、乳脂率、α-乳清蛋白3个泌乳性状值显著增加(P<0.05),当个体基因杂合度为0.3333时,产奶量最高;当个体基因杂合度为0.5000时,乳脂率和α-乳清蛋白含量最高[12]。可见,对不同性状、不同品种来说,适当的基因杂合度能保证其最优的生产性能和种质特性的体现。
在山西白猪群体中,随个体基因杂合度的增加,个体初生重基本呈提高趋势。在杂合度水平为6时(杂合度大于0.5时),初生重最高,但在该杂合度水平下,断奶重和6月龄体重均很低,表明该类个体出生后生长发育缓慢,综合生产性能不佳。杂合度水平为3(平均杂合度为0.373)时,个体初生重也显著高于杂合度水平为1和2的个体,且断奶重、6月龄体重、体长和背膘厚等性状均好于其它类型的个体,这种类型的个体符合我国当前猪育种目标和市场需求,在山西白猪的选育中应优先选留。
[1]姜勋平,熊远著,刘桂琼,等.猪个体基因杂合度对生长性状的影响[J].遗传学报,2003,30(5):431-436.
[2]刘桂琼,姜勋平,熊远著,等.猪个体基因杂合度对胴体性状的影响[J].南京农业大学学报,2004,27(1)67-70.
[3]刘桂琼,姜勋平,熊远著,等.猪个体基因杂合度对肉质的影响[J].南京农业大学学报,2003,26(1):56-60.
[4]姜勋平,熊远著,邓昌彦,等.猪微卫星座位杂合度与肌肉肌苷酸含量的关系[J].畜牧兽医学报,2003,34(4):325-330.
[5]刘国庆,孙业良,杨利国,等.级进杂交绵羊微卫星基因杂合度与生长性能的关系[J].南京农业大学学报,2006,29(1):67-71.
[6]曹果清,薛尚君,杨文平,等.山西白猪高产仔母系繁殖性能测定[J].中国畜牧兽医,2009,36(4):111-114.
[7]曹果清,李步高,莫清珊,等.应用21个微卫星标记分析山西白猪高产仔母系的遗传多样性[J].中国畜牧杂志,2010,46(13):14-18.
[8]金海国,曹阳,姜成国,等.用动物模型估计约克夏猪的主要经济性状遗传参数的研究[J].吉林农业大学学报,2002,24(3):88-91.
[9]M EYER K.WOMBAT-A tool for mixed model analyses in quantitative genetics by REM L[J].J Zhe jiang Uni Science B,2007,8(11):815-821.
[10]鲁绍雄,连林生.SAS统计分析系统在畜牧科学中的应用[M].昆明:云南科技出版社,2003:84-92.
[11]李宁.动物遗传学[M].北京:中国农业出版社,2003:185-191.
[12]姜勋平,毛永江,熊远著,等.牦牛乳蛋白基因杂合度与泌乳性能的关系[J].南京农业大学学报,2003,26(3):68-72.
猜你喜欢
特产研究(2022年6期)2023-01-17
农业工程学报(2022年13期)2022-10-09
东北农业大学学报(2021年9期)2021-11-11
今日农业(2020年17期)2020-12-15
今日农业(2020年15期)2020-12-15
四川动物(2017年6期)2017-12-12
四川动物(2017年4期)2017-07-31
小雪花·小学生快乐作文(2017年1期)2017-03-24
动物营养学报(2015年3期)2016-01-07
