
根系吸氮研究进展
2011-05-09 09:54马娟娟孙西欢李京玲
山西水利 2011年10期
申 星,马娟娟,孙西欢,李京玲,崔 杰
(太原理工大学水利科学与工程学院,山西 太原 030024)
1 引言
我国水资源短缺,水旱灾害频繁,水资源在时空分布上极不均匀,而且水资源的污染与浪费问题严重,水资源的极度短缺和水土环境的持续恶化已成为我国农业乃至整个国民经济发展的瓶颈。
植物的一切正常生命活动,只有在一定的细胞水分含量状况下才能进行。植物的根从土壤中吸收大量水分,以满足植物体的生长需要。在吸收大量水分的同时也会吸收土壤中所溶解的溶质,其中氮素是植物体最重要的营养元素之一。植物根系吸氮是氮素在土壤中循环的一个重要环节,植物根系吸氮量不仅是农田氮素运移模型中的重要参数,也是衡量农田氮肥利用率和制定最优施肥措施的重要依据[1]。
氮素是植物生长不可或缺的营养元素,植物吸收的氮素主要来源于土壤。土壤中的含氮化合物主要有有机含氮化合物、铵盐和硝酸盐,其中有机氮的比例最大,除部分氨基酸及小分子的蛋白质可被植物直接吸收利用外,一般需经矿化作用形成NH4+或经硝化作用形成NO3-,才能被植物根系大量吸收利用。
2 根系吸氮试验
为了解植物根系在土壤中的吸氮情况,许多学者做了大量试验来获得相关信息,以尽可能还原吸氮过程。石建初、左强[2]研究了不同灌溉处理条件下砂培土柱中苗期冬小麦的根系吸氮情况。试验分为四种条件处理:高水高氮、高水低氮、低水高氮、低水低氮。应用反求方法通过迭代求解,并进一步优化获得源汇项中难以直接测定的参数——根系吸氮因子,确定了适用于室内砂培土柱试验中冬小麦的根系吸氮模型。结果表明,根系吸氮总量的模拟值与实测值之间相对误差在10%以内,模型拟合良好,并反求推出此时根系吸氮因子为1.27。由于该试验只涉及室内情况下的苗期冬小麦,有关该方法是否适用于田间情况和冬小麦生长后期尚需更进一步的研究。
贾永国等[3]研究了黄淮冬麦区5个主栽品种(系)在水培试验中的根系吸氮情况。结果表明,在选育氮高效小麦品种时,应尽量选择根系发达的品系,若根干重和根体积较大,则可提高小麦的氮利用效率。该试验只研究了不同基因型小麦在孕穗期的根系吸氮情况,对其他生育期未作进一步研究。
焦峰等[4]应用Logistic模型研究了5种不同氮肥水平对水稻根系吸氮率(SARN)特性的影响,并用SARN值来表征水稻根系活力。结果表明,水稻在整个生育过程中对氮素的单位面积吸收量符合Logistic生长曲线模型,不同处理情况下水稻根系主要在分蘖期表现出对氮素反应能力的差异。试验研究了水稻在完整的生育周期内的根系吸氮情况,较全面地反映了水稻根系对氮素的利用情况,可以更加合理地安排施肥时间与施肥量,提高氮肥的有效利用率,达到节省肥料、减少污染的目的。
彭云峰等[5]研究了玉米根系构型及其在土壤中的空间分布与氮素吸收效率的关系。结果表明,根系在土壤中分布广是氮素能被高效吸收的重要原因之一。
3 根系吸氮模型
根系要吸收氮素必须通过吸水来实现,因此在介绍根系吸氮模型前对根系吸水模型作一简单介绍。
根系吸水模型是在根系吸水机理和相关试验研究基础上建立的,通过数学等方法来定量描述根系吸水(吸水速率)的过程。根系吸水速率的值反映了根系吸水能力的强弱。国内外许多学者对根系吸水模型做了大量研究,使得根系吸水模型不断发展和改进。虽然根系吸水模型有各种型式,但根据对根系吸水研究方法的不同,可将根系吸水模型大致分为微观模型与宏观模型两类。
微观模型由于模型假设太多,所需参数也很难获得,尤其是根系几何形态与分布的详尽资料无法准确得到[6],因此实际应用中很少被采用,应用比较多的是宏观模型。宏观模型的表达式是在田间土壤水分运动基本方程的右边加上一个根系吸水项S而得到的,其数学方程为:Δ[K(θ)Δψ-S],其中:Δ 为矢量微分算子;ψ为总土水势;S为根系吸水项,表示单位时间内根系从单位体积土壤中吸收的水量,即根系吸水速率;K(θ)为土壤导水率;θ为土壤体积含水量。
如何确定根系吸水速率是求解宏观模型的关键。根据建立根系吸水模型方法的不同,可分为两类:一是利用根系密度资料建立经验模型;二是利用土壤水分动态反推根系吸水强度,再依据吸水强度的空间分布情况建立根系吸水模型。
根系吸氮与根系吸水一样,通常用根系吸氮速率(植物根系单位时间内从单位土体中吸收氮素的质量)来表示。就目前而言,根系吸氮速率很难通过实际测定得到,一般通过建立根系吸氮模型(即根系吸氮速率函数)来获得。根系吸氮模型可分为微观模型和宏观模型两种。
微观模型用拟稳定流或稳定流的方式来研究氮素从土壤溶液向植物根表运移的过程[7]。最具代表性的是建立在5个基本假设基础之上的DAISY模型。这5个基本假设是:每一条根都支配着一个以其为中心轴的圆柱体所构成的平均有效土壤体积;该圆柱体的半径约为根系间平均距离的1/2;氮素以质流和扩散的形式从土壤溶液中运移向根表处;氮素浓度在径向剖面上随时间逐渐变化,并近似为稳定流剖面;运移到根表处的氮素都能被根系所吸收[8]。由于假设较多、模型参数不易获得,使得微观模型离实际应用还有很大差距。
宏观模型将植物的根系作为整体来考虑,不考虑根系中的细微过程,并且各模型的方法各不相同[9]。Hansen等[7]对已有的宏观模型进行了总结,在假设植物根系表面氮素浓度与土壤溶液中的氮素浓度一致的基础上,归纳出了以下四种计算根系吸氮的方法:
一是假设土壤中存在着一个比例关系。该比例关系是一个与土壤深度有关的函数,其中的无机态氮(硝态氮和铵态氮)都可以被植物根系吸收利用[10]。该方法的经验性太强,存在很大的不确定性。
二是假设在植物根区范围内有一个适宜的浓度范围,超出这个浓度范围的所有无机态氮都可以被植物根系吸收利用。该方法同样经验性太强,且计算结果精度不高。
三是根据米氏动力学方程计算根系吸氮量[11]:
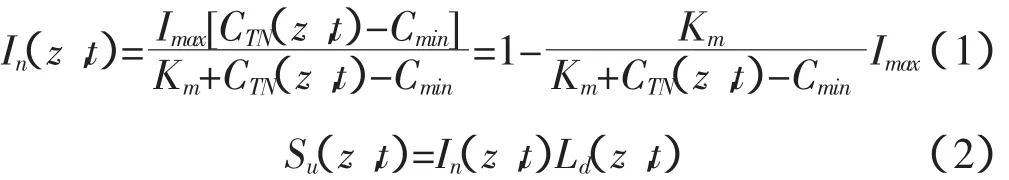
其中:t为时间;In(z,t)为单位根长潜在根系吸氮系数,是指在最优水分条件下,单位长度根系单位时间内吸收氮的质量;Imax是指在最优土壤养分条件下的单位根长潜在根系吸氮系数;CTN(z,t)为土壤溶液的氮素浓度,是指单位体积土壤溶液中铵态氮或硝态氮的质量;Km为当In=0.5Imax时的氮素浓度;Cmin为当In=0时的氮素浓度;Su(z,t)为根系吸氮速率。由于模型中的动力学参数如Imax,Km与Cmin受温度、耕作方式、植物生长、植物养分含量、土壤含水量、根系发育、根系形态等因素的影响,所以很难准确获得[12],目前对这些方面的研究还比较欠缺。
四是根据植物的蒸腾速率或吸水速率和土壤溶液中的氮素浓度计算植物根系的吸氮量[13]:
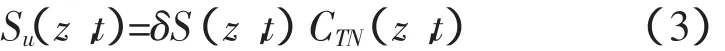
其中:δ为根系吸氮因子,δ≥0,反映了植物根系对氮素吸收的主动性和被动性。该方法引入根系吸氮因子δ,体现了溶质与植物自身的特性,概念清楚,计算简便,所需参数较少。该模型即为前文石建初、左强试验中应用的模型,之所以利用根系吸水速率来表示根系吸氮速率,是因为植物根系在吸收土壤中水分的同时吸收着土壤溶液中的氮,尽管两个过程的机制可能有所不同,但都表现出一些共同的性质:根尖对水分与养分的吸收能力最强,距离根尖越远、根龄越大,根系的吸收能力就越弱[14]。对于模型中的根系吸氮因子,目前国内外的研究很少,取值多为经验值[15]。一些学者认为根系吸氮因子主要由植物种类和溶质决定,于是假设根系吸氮因子对特定的植物种类和溶质在根区范围内为一个常数[13]。因此,该模型能否被广泛应用的关键就在于如何确定根系吸氮因子的值。
4 结论
通过对植物根系吸氮情况的研究,运用试验、建模等手段可使植物整个生育期的吸氮过程更加清晰,有利于进行更深层次的研究。这些研究虽然取得了很大进展,但仍存在很多需要解决与改进的地方。
第一,植物根系生长分布由于受植物种类、大气条件、土壤质地等因素制约,根长密度很难准确得到,而根长密度又是研究吸氮效率和建立吸氮模型的一个重要参数,因此大多需依据根系生长分布模型来确定根长密度,这就给试验和建立模型带来了不便。
第二,目前大多已进行过的试验都是粮食作物,如玉米、小麦、水稻等,对一些经济作物及各类果树的相关研究还没有或很少,因此应进行一些相关方面的研究,以获得更多的数据,深入了解不同植物根系吸氮机理。而且,许多试验是在室内模拟条件下完成的,尚未在大田试验中获得可信数据,因此需进行更深一步的研究。
第三,根系吸氮模型的许多假设条件都是在理想状况下建立的,与实际状况有很大差别,根系的实际吸氮情况很难掌握,不同土层深度的根系吸氮能力也有所不同。很多模型所需参数很难实际测得,影响参数的因素太多,因此大多数模型都是经验模型,只能应用于特定植物或特定地区,很难得到广泛应用。
因此,为深入了解植物根系吸氮情况,必须加强对根系吸氮试验和吸氮模型的研究,以更直观地了解根系吸氮机理,从而达到有效节水、减少氮肥浪费、提高环境质量的目的。
[1] 张瑜芳,张蔚榛.农作物对氮肥吸收速率的研究现状[J].灌溉排水,1996,15(3):l-8.
[2] 石建初,左强.不同灌溉处理条件下苗期冬小麦土柱中的硝态氮运移模拟[J].灌溉排水学报,2009,28(5):1-6.
[3] 贾永国,安调过,李俊明,等.不同小麦基因型孕穗期根系性状与吸氮量的关系[J].华北农学报,2006,21(3):37-40.
[4] 焦峰,吴金花,张兴梅,等.不同氮肥水平影响下的水稻根系吸氮率特性分析[J].土壤通报,2009,40(6):1353-1355.
[5] 彭云峰,张吴平,李春俭.不同氮吸收效率玉米品种的根系构型差异比较:模拟与应用 [J].中国农业科学,2009,42(3):843-853.
[6] 邵明安,黄明斌.土一根系统水动力学[M].陕西:陕西科学技术出版社,2000:131-155.
[7] Hansen,S.,Jensen,H.E.andShaffer,M.J.Developmentsinmodelingnitrogentransformationsinsoil.InBacon,P.E.(Ed.).NitrogenFertilizationintheEnvironment[M].MarcelDekker,NewYork,1995:3-107.
[8] Hansen,S.,Jensen,H.E.,Nielsen,N.E.,Svendsen,H.Simulationof nitrogendynamicsandbiomassproductioninwinterwheatusingtheDanishsimulationmodelDAISY [J].FertilizerRcsearch,1991(27):245-259.
[9] Barber,S.A.Soilnutrientbioavailability:AmechanisticApproach[M].NewYork,1995:69-132.
[10] Johnsson,H.,Bergstr觟m,L.,Jansson,P.andPaustian,K.Simulatednitrogendynamicsandlossesinalayeredagricultural soil[J].Agric.Ecosyst.Environ,1987(18):333-356.
[11] 李韵珠,李保国.土壤溶质运移[M].北京:科学出版社,1998:195-198.
[12] 龙怀玉.土—根系统中水分对硝酸根运移、吸收的影响[C].北京:中国农业大学,1995.
[13]Schoups,G.,andHopmans,J.W.Analyticalmodelforvadose zonesolutetransportwithrootwaterandsoluteuptake[J].VadoseZone,2002(1):158-171.
[14] 马元喜,王晨阳,贺德先,等.小麦的根[M].北京:中国农业出版社,1999:12-33,59-69.
[15] 张瑜芳,张蔚榛,沈荣开,等.排水农田中氮素转化运移和流失[M].武汉:中国地质大学出版社,1997:15-19.
猜你喜欢
四川劳动保障(2021年3期)2021-06-09
中学生数理化·高一版(2020年6期)2020-07-25
现代园艺(2017年13期)2018-01-19
中学生数理化·八年级物理人教版(2017年3期)2017-11-09
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
上海金属(2014年5期)2014-12-20
食品工业科技(2014年23期)2014-03-11
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26