
铜胁迫下植物与土壤的耦合响应研究
2011-08-03 07:14单奇华刘先虎张建锋陈光才刘胜刚张平选
水土保持通报 2011年5期
单奇华,刘先虎,张建锋,陈光才,刘胜刚,张平选,王 瑛
(1.中国林业科学研究院 亚热带林业研究所,浙江 富阳311400;2.铜陵市铜官山森林公园,安徽 铜陵244000;3.铜陵市林业技术管理站,安徽 铜陵244000)
土壤有机质和全氮影响草甸植物群落地上生物量、物种多样性和群落总盖度[8]。在低磷胁迫中,杉木(Cunningha mial anceol ata)、马尾松(Pinusmassoniana)和杨树(Popul us)都有根系酸化机制以利于吸收土壤中固定态磷[9-11]。低氮胁迫将显著抑制马褂木苗叶片发育和根系生长,影响苗高生长和干物质积累,但苗木光合产物则较多地分配至地下根系部分以促进对氮素的吸收利用[12]。海州香薷(Elsholtziasplendens)、鸭跖草(Commelinacommunis)通过耐性对策,狗牙根(Cynodondactylon)、狗尾草(Setariaviridis)通过微生境对策,白茅(Imperatacylindricavar.major)通过根茎对策生于各类重金属污染土壤[13]。
以往大都以土壤因子为自变量,植物生理生态指标为因变量,研究并阐明了植物针对不同土壤条件的适应性调节机制,缺少对土壤与植物相互作用的动态过程研究。本研究以铜胁迫退化土壤和抗逆性植物(乔、灌、草)为基本材料,用盆栽法室内模拟了自然界中铜污染退化土壤对其适生植物的选择过程,研究并阐明了这一选择过程的驱动力,同时探讨了适生植物对铜胁迫退化土壤的改良作用。通过研究旨在揭示特定土壤与植物的耦合响应机制,了解自然界中某些特定生态系统发育的起因和过程,为人类对退化生态系统进行人工修复提供科学依据。
1 材料与方法
1.1 试验材料
CuSO4×5 H2O(分析纯)购自富阳市科导化工有限公司。塑料花盆,口径20 c m,深度20 c m。黄壤取自浙江省富阳市,地理坐标为北纬30°3′41.72″,东经119°57′4.28″,过5 mm 粗筛,砂土(美国制分类标准),弱酸性(p H值为4.89),土壤生物代谢活性低(脱氢酶活性为0.96μg/g),养分水平较低,土壤铜元素含量处于正常水平(表1)。植物材料有楸树(Catal pabungei)、梓树(Catal paovataG.Don)、麻栎(Quercusacutissi maCarr uth.)、紫穗槐(Amor pha f r uticosaL.)、高羊茅(Festucaar undinacea)、佛甲草(SedumlineareThunb)和葛藤(Puer arialobata)。楸树、梓树、麻栎和紫穗槐为一年生苗木,葛藤采用分株繁殖(取自中国林科院亚林所沿海防护林课题组苗园),高羊茅为种子繁殖,佛甲草为根/茎营养体繁殖(购自鑫和花卉苗木公司)。

表1 盆栽用土背景值
1.2 试验设计
(1)根据植物生态学原理,植物群落的原生演替的旱生演替系列依次为地衣植物阶段、苔藓植物阶段、草本植物阶段和木本植物阶段。植物群落的次生演替的森林的采伐演替系列依次为采伐迹地、杂草群落、小叶树种阶段和目标树种阶段。人为干预能够加速或打乱植物群落的演替序列,如人们通过植物筛选并应用乔、灌、草搭配的方式生态修复矿区裸地、盐碱地、水涝或干旱荒地等,试图按照自己的意愿构建人工植物群落。但植被及植物群落的分布,在区域内是受地方气候、地形地势和土壤等因素的综合影响而成规律性分布的。因此,试验假设在铜胁迫土壤上人工配植抗逆乔、灌、草植被的情况下,研究铜胁迫土壤与植物的相互作用方式,以揭示人工植物群落在铜胁迫土壤上的演替机制。
(2)正常土壤中的总Cu含量一般为2~250 mg/kg,平均为30 mg/kg,而有效态Cu(DTPA)含量则为0.1~10 mg/kg。野外实地研究证实,土壤中总Cu含量达到150~400 mg/kg,或者是有效态Cu(DTPA)超过15 mg/kg时,就会对植物产生毒害[14]。根据这一条件,采用自然贫瘠黄壤中添加外源CuSO4×5 H2O药品的方式,模拟自然铜胁迫土壤。
将一定计量的Cu SO4×5 H2O药品直接混入过筛黄壤,拟形成以纯铜计含量为50 mg/kg的铜胁迫退化土壤(处理2,模拟自然界中受铜污染的退化土壤),放置稳定一周,同时作空白对照(处理1,土壤中不添加CuSO4×5 H2O),处理2和处理1均设3个重复,每重复4盆,每盆装土4 kg(以烘干土计),木本植物和葛藤每盆栽植1棵苗木,高羊茅每盆施种50粒,佛甲草每盆种植营养繁殖体10小段(3 c m/段)。植物上盆1周后作背景值调查(木本植物的树高和地径,见表2),培养4个月后分别统计处理1和处理2梓树、楸树、麻栎、紫穗槐和葛藤的存活率,高羊茅和佛甲草的发芽率,以及各植物的生长量(木本植物的树高和地径生长率,草本和葛藤的生物量),同时采集植物样本和土壤样本,室内用去离子水冲洗植物样品,自然风干,105℃杀青0.5 h,75℃烘干至恒重,用于测定植物生物量,土壤样品自然风干,磨碎,过筛,用于分析各土壤指标。
由于试验用黄壤本身含有一定量的铜元素(表1)以及CuSO4×5 H2O药品与黄壤混合过程的试验误差,使得处理2上盆土壤的实际铜元素含量与设计的理论值有一定偏差,经过分析测定,处理2土壤中实际全铜含量为80.3 mg/kg,有效铜含量为49.45 mg/kg。

表2 树高和地径背景值(平均值)
1.3 土壤和植物样品测定方法
土壤p H值:用25 ml去离子水浸提过2 mm筛的风干土壤样品(10 g),p H计法;土壤颗粒组成:过2 mm筛土样,比重计法;土壤有机质:重铬酸钾外加热法;土壤全氮:凯氏消煮法;有效氮:碱解—扩散法;全磷:氢氧化钠碱熔—钼锑抗比色法;土壤有效磷:碳酸氢钠浸提钼蓝比色法;全钾:氢氧化钠碱熔—火焰光度法;速效钾:1 mol/L乙酸铵浸提—火焰光度法;脱氢酶:2,3,5-三苯基四唑化氯(TTC)比色法,以每克土壤的2,3,5—三苯基甲谮的毫克数表示[15];土壤全铜:硫酸—高氯酸消煮,原子吸收分光光度法法;土壤有效铜:0.1 mol/L盐酸浸提,原子吸收分光光度法;植物生物量:干重称量法[16]。
It should be noted that this might be the first time that the word “independence”was associated with Tibet within a United States government document.
1.4 数据处理
不同类型的植物对逆境适应性反映的敏感指标不同,如试验用木本植物梓树、楸树、麻栎和紫穗槐为一年生苗木,其存活率(S)和与树高(H)和地径(D)生长率指标能够较好地反映铜胁迫对这些苗木生长的影响;草本植物葛藤为分株繁殖,并每株保留2张小叶片,用存活率和生物量(W)指标能有效表征葛藤对铜胁迫的适应性;佛甲草用根、茎营养体繁殖,高羊茅用种子繁殖,均浅埋于盆栽土壤中,用发芽率(G)和生物量指标能直观地反映铜胁迫对佛甲草和高羊茅生长的影响。

式中:S——存活率(%);n——某一处理内某一植物的存活数(株);N——某一处理内某一植物的总数(株)。
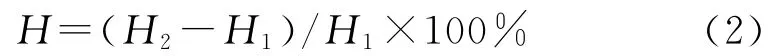
式中:H——树高生长率(%);H2——培养4个月后树高(c m);H1——树高背景值(c m)。
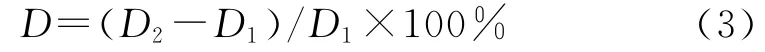
式中:D——地径生长率(%);D2——培养4个月后地径(mm);D1——地径背景值(mm);G——每盆茎段或种子的发芽数量与每盆茎段或种子的种植总数的比值(%)。
统计分析用t—test和图表用SAS 8.0和Excel软件处理。
2 结果与分析
2.1 铜胁迫退化土壤对适生植物的选择过程
在无添加外源Cu SO4×5 H2O药品的土壤条件下(处理1),梓树、楸树、麻栎和紫穗槐均全部成活,且树高生长率均超过10%,地径生长率接近10%左右(表3);葛藤成活率为100%,生物量较高(15.75±7.85 g/盆);高羊茅和佛甲草发芽率分别为88%和100%,生物量分别为3.46±0.29 g/盆和2.68±0.77 g/盆。表明试验植物在无铜胁迫的贫瘠土壤中均能正常生长,表现出较好的抗贫瘠性,另外,表明试验采用的土壤自身不存在极端因子以限制试验植物的正常生长,为下文铜胁迫对适生植物的选择排除了干扰因子。
对比处理1,梓树和麻栎在处理2(铜胁迫)下成活率降低近一半(表3),树高生长率显著降低(t—test,p<0.05),铜胁迫对梓树和麻栎生长影响较大,梓树和麻栎适宜在铜浓度小于50 mg/kg的土壤环境中生长。在铜胁迫下,楸树和紫穗槐能正常存活,但楸树高生长受到较大抑制,显著低于处理1树高生长率(p<0.05),而紫穗槐高生长不受铜胁迫的影响,紫穗槐表现出较好的耐铜特性。铜胁迫对各木本植物的地径生长影响不大。与处理1相比,铜胁迫下高羊茅发芽率仅为42%,生物量显著降低(p<0.05,图1),表现出较高的铜毒害敏感性;佛甲草发芽率为70%,生物量略有降低但不显著;葛藤全部存活且长势不减,生物量仍高达12.73±3.89 g/盆,葛藤表现出较好的耐铜特性。
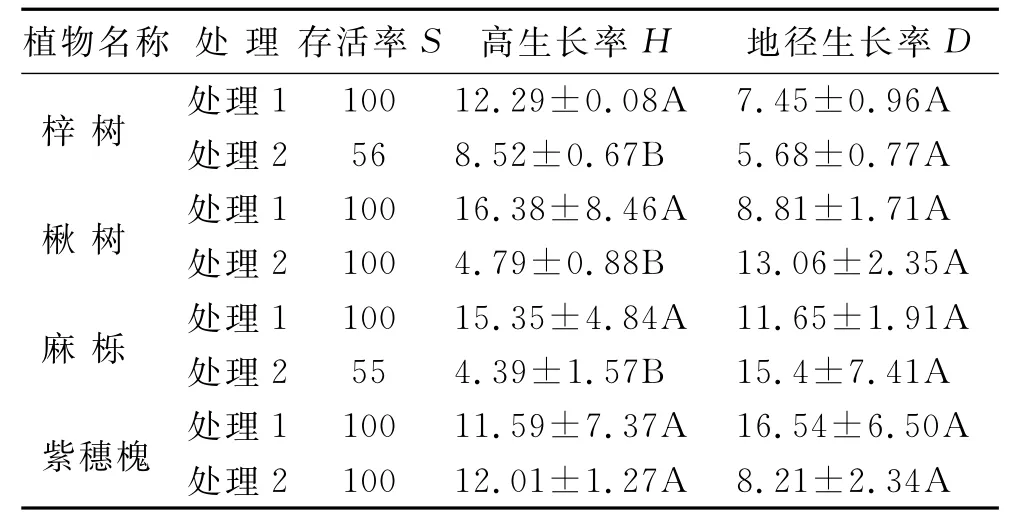
表3 不同处理植物存活率和生长率统计分析
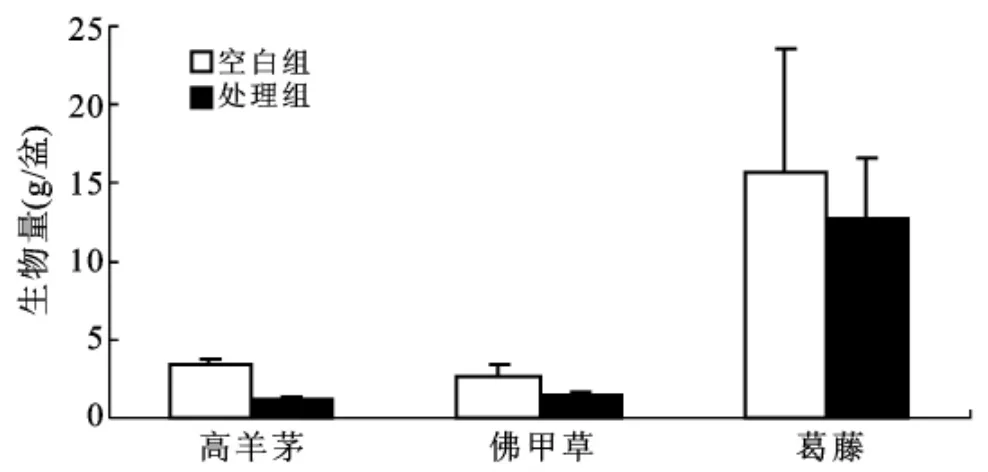
图1 不同处理草本生物量分析
研究表明在铜胁迫退化土壤条件下,紫穗槐和葛藤将成为适生植物,可着生于高浓度铜胁迫环境中;楸树,麻栎和梓树为铜耐性植物,可散生于局部低浓度铜胁迫环境中,而高羊茅和佛甲草不适宜在铜胁迫环境中生长,将被逐步淘汰。
2.2 适生植物对铜胁迫退化土壤的改良作用
紫穗槐和葛藤为铜胁迫适生植物,楸树和麻栎为铜耐性植物,以紫穗槐、葛藤、楸树和麻栎为例,研究植物生长对铜胁迫退化土壤的改良作用。结果显示:
植物措施后(4个月培养),铜胁迫土壤的p H值分布在4.79~4.99之间,平均值为4.91,与盆栽前土壤(背景值)p H=4.89比,植物措施前后土壤的p H值没有显著性差异(t—test,p=0.72),表明植物生长在初期对土壤p H值的改良作用有限。
对比背景值(表1),植物措施后土壤的有机质含量都普遍提高(表4,p<0.01),植物生长过程中由根系向土壤中释放的各类有机物质是植物生长初期促进土壤有机质含量升高的主要原因。土壤全氮水平植物措施前后保持稳定,植物初期生长对土壤全氮含量无显著影响(p=0.43),紫穗槐和葛藤虽为固氮植物,但在试验期内根部未发现根瘤,因而对土壤中氮元素的补充作用有限。植物措施后土壤中全磷含量均低于背景值(p<0.01),表明植物生长初期就能解析并吸收土壤中磷元素,导致土壤全磷降低,铜胁迫下植物的磷素利用机制还有待进一步研究。植物措施后土壤中全钾含量降低(p<0.01),而有效钾含量反而升高(p<0.05),土壤全钾含量的降低表明植物生长初期会导致土壤钾缺乏,而有效钾含量异常升高的现象表明在铜胁迫下土壤中存在除植物因素外的其它解钾机制。

表4 适生植物对铜胁迫土壤养分的改良分析
土壤中具有活性的脱氢酶是生物细胞的必要组成部分,只能存在于生物体内,由于土壤中不可能存在脱离活体的脱氢酶,测得的脱氢酶活性多表征土壤微生物的瞬时代谢活性[15]。不同植物措施后,铜胁迫土壤的脱氢酶活性(TPFμg/g)分别为麻栎:0.37±0.06,紫穗槐:1.61±0.3,楸树:0.37±0.05和葛藤:0.37±0.39。对比背景值(0.96TPFμg/g),紫穗槐有效提高土壤脱氢酶活性(t—test,p<0.01),而其它植物措施下的土壤脱氢酶活性反而降低(p<0.01)。紫穗槐的生长几乎不受铜胁迫的影响,因此表现出较好的改良土壤酶活性能力。其它植物在铜胁迫下生长率明显下降,因而也制约了改良土壤酶活性的能力。表明植物改良土壤脱氢酶活性的能力与其耐铜性和铜胁迫下的生长率有一定的相关性。对比背景值,不同植物措施对铜胁迫退化土壤的全铜含量都有降低作用(t—test,p<0.01),其中紫穗槐降低量最高为ΔCu(全量)=32.5 mg/kg,其次为楸树(28.83 mg/kg),最后为麻栎(23.89 mg/kg)和葛藤(22.54 mg/kg,表5),植物降低土壤中铜元素含量的能力与该植物在铜胁迫下的生长率和生物量有相同的变化趋势。

表5 植物措施后土壤中铜元素含量分析 mg/kg
3 结论
土壤通过调节土壤指标对植物生长产生影响,以植物生长的需求度为标准,可以把土壤指标分为植物生长需求性指标(如孔隙度、养分指标和酶活性等)和植物生长限制性指标(如过量重金属和有机污染物)。美国生态学家V.E.Shelf ord(1913)提出耐受性定律(law of tolerance),认为一种生物能不能存在与繁殖,要依赖于一种综合环境的全部因子的存在,但只要其中一项因子的量或质不足或超过了某种生物的耐受限度,则会使该物种不能生存,甚至灭绝。这一定律把因子最小量和最大量并提,任何接近或超过耐受性下限或上限的因子都称为限制因子。因此,对于退化(或污染)土壤,揭示土壤中植物生长的限制因子及其阈值,是研究该类型土壤对植物生长影响的关键。
本试验中,各植物在无铜胁迫退化土壤上均能正常生长,而在铜胁迫下不同植物表现出不同的存活率和生长率(生物量),研究表明铜元素是铜胁迫退化土壤选择适生植物的驱动因子(限制因子)。在铜元素的主导下,铜含量不同的土壤分别选择其适生植物。研究显示有效铜含量在49.45 mg/kg,全铜含量在80.3 mg/kg左右的土壤对几种试验植物的优先选择顺序依次为紫穗槐=葛藤>楸树> 麻栎=梓树>佛甲草>高羊茅。紫穗槐和葛藤将成为铜胁迫退化土壤的优势植物,楸树、麻栎和梓树则受到抑制,呈局部散生,而佛甲草和高羊茅将被逐步淘汰。
适生植物生长初期对铜胁迫退化土壤p H值的改良作用不明显,对土壤全氮含量无显著影响,能解析土壤固定态磷和钾并较快地吸收磷和钾元素而导致土壤微域磷和钾缺乏,促进土壤有机质含量和脱氢酶活性的提高,有效降低土壤中铜元素含量。植物与土壤的耦合响应表现出高度不对称性,即土壤往往通过自身某一极端因子就可以直接影响植物的生长,而植物对土壤多因子可同时产生影响。
植物与土壤的耦合响应是主导陆地生态系统类型和发育程度的重要因素之一。通过室内模拟试验,揭示了铜胁迫下植物与土壤的耦合响应机制,初步了解在该类型土壤上生态系统自我恢复的一些发生规律。但对于适生植物与特定土壤耦合响应的长期效应,以及其它类型陆地生态系统中植物与土壤的耦合响应机制还有待进一步研究。
致谢:感谢中国林业科学研究院亚热带林业研究所虞木奎研究员对试验植物材料所提供的帮助。
[1] 万里强,李向林.系统耦合及其对农业系统的作用[J].草业学报,2002,11(3):1-7.
[2] 彭晚霞,宋同清,曾馥平,等.喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系[J].生态学报,2010,30(13):3472-3481.
[3] 董孝斌,高旺盛.关于系统耦合理论的探讨[J].中国农学通报,2005,12(1):290-292.
[4] Gruber B R,Schultz H R.Coupling of plant to soil water stat us at different vineyar d sites[J].Acta Hort,2005,689:381-390.
[5] Ladanai S,Ågren G,Olsson B.Relationships bet ween tree and soil properties inPiceaabiesandPinussyl vestrisforests in Sweden[J].Ecosystems,2010,13:302-316.
[6] 徐学选,张北赢,白晓华.黄土丘陵区土壤水资源与土地利用的耦合研究[J].水土保持学报,2007,21(3):166-169.
[7] 郑姚闽,崔国发,雷霆,等.甘肃敦煌西湖湿地植物群落平均盖度与土壤含盐量耦合关系[J].生态学报,2009,29(9):4665-4671.
[8] 江源,章异平,杨艳刚,等.放牧对五台山高山、亚高山草甸植被—土壤系统耦合的影响[J].生态学报,2010,30(4):0837-0846.
[9] 俞元春,余健,房莉,等.缺磷胁迫下马尾松和杉木苗根系有机酸的分泌[J].南京林业大学学报:自然科学版,2007,31(2):9-12.
[10] 张焕朝,王改萍,徐锡增,等.根际酸化作用对杨树无性系磷营养效率的影响[J],应用生态学报,2003,14(10):1607-1611.
[11] 张焕朝,王改萍,徐锡增,等.杨树无性系根系吸收动力学特征与磷营养效率[J].林业科学,2003,39(6):40-46.
[12] 樊瑞怀,杨水平,周志春,等.氮素营养对马褂木家系苗木生长效应分析[J].林业科学研究,2009,22(1):85-90.
[13] 蔡锡安,夏汉平.森林生态系统中草层植物的生态功能[J].热带亚热带植物学报,2003,11(1):67-74.
[14] 束文圣,杨开颜,张志权,等.湖北铜绿山古铜矿冶炼渣植被与优势植物的重金属含量研究[J].应用与环境生物学报,2001,7(1):7-12.
[15] 周礼恺.土壤酶学[M].北京:科学出版社,1987:267-281.
[16] 中华人民共和国林业部科技司.林业标准汇编[S].北京:中国林业出版社,1991:102-255.
猜你喜欢
广东蚕业(2022年4期)2022-11-23
现代农村科技(2022年6期)2022-11-18
润·文摘(2021年9期)2021-09-22
环境保护与循环经济(2021年5期)2021-07-23
湖北农机化(2020年16期)2020-10-21
江西农业(2020年13期)2020-09-08
红岩(2018年6期)2018-11-16
中国饲料(2016年14期)2016-02-03
少儿科学周刊·儿童版(2015年1期)2015-07-07
青少年科技博览(中学版)(2014年4期)2014-07-31
