
中原牡丹传统品种的核型及进化关系1)
2012-06-28 09:12史倩倩
东北林业大学学报 2012年11期
史倩倩 王 雁 周 琳
(林木遗传育种国家重点实验室(中国林业科学研究院林业研究所),北京,100091)
黄国伟
(湖北林业科学研究院)
牡丹(Paconia suffruticosa Andr.)是芍药科芍药属名贵的观赏植物和药用植物,作为中国传统名花的优秀代表,其姿态万千、色彩丰富,深受人们喜爱。中国中原牡丹品种数量最多,栽培历史悠久、分布最广,形态变异最丰富。牡丹品种分类的方法较多,目前以花型分类的方法得到了广泛的应用,共分为2类 4 亚类 16 型[1]2-17。
核型是遗传物质在细胞水平上的表征,与外部形态相比核型受外界环境影响小,是推测物种起源、演化、分类和鉴别的重要依据[2]。谭远德等[3]在1993年提出,以核型数据为基础的核型似近系数聚类法,能较好地克服传统形态分析带来的局限性,并做到了物种间核型相似的数量化,且多向、立体、多维地考察物种间的亲缘关系,使结果客观可靠。牡丹传统品种[1]8-9有别于现代品种,具有更丰富的遗传信息,但前人的相关研究均未区分传统品种与现代品种。前人对牡丹野生种及少量栽培品种进行了细胞学研究[4]286-292,[5-17],探讨了野生种及栽培品种的核型参数及野生种间的演化关系,但都未深入研究形态和核型参数之间的关系,没有反映传统品种及现代品种的进化关系及现代品种遗传多样性的丢失,并且涉及传统品种的研究中品种类型的代表性不足。本试验通过对21个典型不同花型的传统中原牡丹品种核型进行比较和聚类分析,探讨传统品种间核型差异,进而探讨染色体变异和花型之间的关系,旨在为牡丹品种演化研究提供线索,同时为深入研究牡丹传统品种的细胞分类学问题奠定基础。
1 材料与方法
试验材料取自中国林业科学研究院玉泉山牡丹基地。21个中原牡丹传统品种涵盖了7个花型[1]52-207(表 1)。

表1 供试材料
2011年3月中下旬取直径为0.5~1.5 cm的花蕾,剥除花瓣,只留花药和子房,子房切开,用V(0.05%秋水仙素)∶V(0.02 mol·L-18 - 羟基喹啉)=1∶1混合液室温下预处理5 h,经卡诺固定液(V(95%乙醇)∶V(冰乙酸)=1∶3)于4℃固定24 h,经95%乙醇冲洗2次后,转入70%乙醇中,再放入1 mol·L-1盐酸中,并于60℃恒温金属浴解离10~15 min,蒸馏水洗净后,用卡宝品红染色和常规压片法制片,用蔡司AXIOIMAGER A1光学显微镜分别对各材料的50个染色体分散良好的中期分裂相进行拍照。
核型分析:每个品种选出形态清晰、伸展较佳的5个细胞[2],借助AXIOIMAGER软件分别测量染色体及长短臂的大小。核型按Levan的分类法[18]和李懋学等[4]5-22的植物染色体标准化的规定分析;核型分类参照Stebbins[19]的核型分类标准;核型不对称系数参照 Arano[20]公式。
聚类分析:按谭远德等[3]提出的核型似近系数聚类分析方法和公式计算核型似近系数(λ);按吴昌谋[21]提出的公式计算核型进化距离(De),对似近系数利用平均聚类法(UPGMA)进行聚类。
2 结果与分析
2.1 21个中原传统牡丹品种染色体核型的属性特征与品种间的相关性
研究结果表明,供试21个品种的染色体核型稳定,但品种间存在一定的差异。21个中原牡丹品种的染色体照片及核型模式见图1、图2,主要核型参数见表2。从图1、表2可以看出,除‘首案红’是三倍体,有15条染色体外,其他全部为二倍体,染色体数目为10;除‘茄蓝丹砂’的核型类型为2B型外,其余品种的核型类型均为2A,核型不对称系数介于59.21% ~95.70%,属于比较对称的核型,说明‘茄蓝丹砂’的进化程度相对较高。其观察统计结果表明:只有‘脂红’没有发现随体,其余20个品种均具有随体染色体。其中‘蓝田玉’含随体最多,具有8个随体染色体,分别分布在第1、3、4、5对染色体上。同时发现花型为皇冠型的牡丹品种具有的随体染色体数目普遍较多。
21个传统中原牡丹品种的核型公式有5种形式(表2):2n=6m+2sm+2st,有‘黄花魁’、‘酒醉杨妃’、‘茄蓝丹砂’、‘大棕紫’、‘锦袍红’、‘万花盛’、‘姚黄’、‘赵粉’、‘魏紫’、‘小魏紫’、‘假葛巾紫’、‘桃红献媚’和‘青龙卧墨池’共13个品种;2n=8m+2st,有‘大棕紫’、‘白玉’、‘豆绿’、‘金玉交章’和‘蓝田玉’共5个品种;2n=4m+6sm,品种为‘脂红’;2n=8m+2sm,品种为‘赤龙焕彩’;2n=15=9m+3sm+3st,品种是‘首案红’。
21个中原牡丹传统品种的臂比均值为1.65~2.21,其中‘锦袍红’的臂比均值最大,为 2.21,‘赤龙焕彩’的臂比均值最小,为1.65(表2)。表3、表4表明各花型之间的臂比均值差异显著,蔷薇型的臂比均值最大,为2.14;其次为菊花型,均值为2.04;千层台阁型的臂比均值最小,为1.76。六大花型中楼子台阁型的臂比均值方差最大,为0.024 0;菊花型的臂比均值方差最小,为0.000 1;说明楼子台阁型内的臂比均值差异最大,其次为皇冠型、蔷薇型、千层台阁型、荷花型,而菊花型最一致。
通过6个花型的核型不对称系数的比较得出(表3),6个花型间差异不显著,核型不对称系数为60.22~79.12。但花型内部的方差从大到小依次为皇冠型、楼子台阁型、蔷薇型、菊花型、荷花型和千层台阁型。其中花型为皇冠型的‘首案红’的核型不对称系数明显最大,为95.70%。
2.2 21个中原传统牡丹品种亲缘关系相关性的聚类分析
依据核型似近系数和核型进化距离的计算原理与方法计算出21种中原牡丹传统品种两两之间的核型似近系数和进化距离,再利用平均聚类法(UPGMA)进行聚类,结果见图3。
在21种中原牡丹传统品种中,‘豆绿’与‘姚黄’的核型似近系数最大,为0.998 2,进化距离最小,为0.001 8,亲缘关系最近;‘白玉’与‘魏紫’的核型似近系数最小,为0.852 6,进化距离最大,为0.159 5,亲缘关系最远。经统计,各供试材料间的核型近似系数均在0.850 0以上,进化距离间隔很小,这说明了21个中原牡丹传统品种在核型上稳定性和相似性很高。由图3可见,21个中原牡丹传统品种在核型进化距离为0.0552、似近系数为0.9420处聚成两大类:‘白玉’和‘魏紫’独立于其他19个品种聚成一类。其中‘首案红’在进化距离0.040 5、似近系数0.960 3时与其他18个品种聚成一类,而这18个品种在进化距离0.014 0、似近系数0.986 1时又分为两大类:‘黄花魁’、‘桃红献媚’、‘大棕紫’和‘黑花魁’4个品种聚成一类,其他14个品种聚成一类。属于托桂型的‘青龙卧墨池’和属于蔷薇型的‘锦袍红’在似近系数0.975 6时聚为一类。7类花型21个中原牡丹传统品种的核型聚类结果并没有完全依据花型分类。
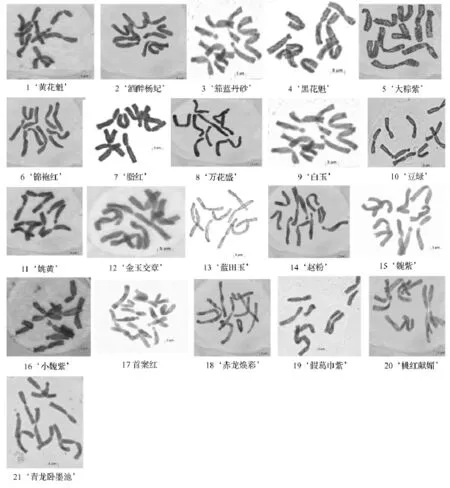
图1 21个中原传统牡丹品种有丝分裂中期图
3 结论与讨论
本研究通过对7类花型的21个中原牡丹传统品种核型参数的研究,发现‘首案红’的染色体为2n=15,三倍体;其他20个品种的染色体为2n=10,二倍体,与前人研究的结果相吻合[17],同时说明了‘首案红’的进化较其余品种高。
杨涤清等[7]和张赞平等[11,15]都认为牡丹种间核型的差异主要表现在随体的数目和位置上,并认为随体数目及随体染色体的杂合性是牡丹种间差异的一个重要的细胞学特征。本试验的21个品种中,只有‘脂红’没有发现随体,说明该品种的进化程度相对其他品种有所不同。随体位置主要涉及到第1、2、4、5对染色体短臂上,而‘蓝田玉’、‘假葛巾紫’和‘桃红献媚’在第3对染色体的短臂也出现了随体,说明这3个品种的进化程度高。观察发现花型为皇冠型的品种具有随体的数目较多。因此,随体数量及位置的差异是否反映了供试21个品种起源的差异有待进一步研究。

图2 21个中原传统牡丹品种的染色体核型模式图
著名植物分类学家和进化学家 Stebbins[19]提出,被子植物核型进化的基本趋势是由对称向不对称发展的。在系统演化上处于较古老或原始的植物,往往具有较对称的核型,而不对称的核型则通常出现在较进化或特化的植物中。本试验中的20个牡丹传统品种核型类型是较为对称的2A型,只有‘茄蓝丹砂’的核型类型为2B型,说明‘茄蓝丹砂’比其他20个品种进化程度高,可能相对而言产生的年代要晚一些。
依据侯小改等[17]的牡丹进化观点,牡丹品种的核型存在着多样性,这也许跟牡丹拥有多种花型及花型易变异的现象有着密切的联系。得出以下结论:具有2n=6m+2sm+2st的品种不对称性最高;具有2n=4m+6sm和2n=8m+2st的品种不对称性次之;具有2n=8m+2sm的品种最原始,即‘赤龙焕彩’。核型不对称系数是反映染色体对称与否、进化与否的另一个参数指标[4]171。在本试验中核型不对称系数以‘首案红’最大,而‘赤龙焕彩’则属于较小的,说明‘赤龙焕彩’的产生时间早于其他供试品种,较原始,而‘首案红’比较复杂,是相对进化或原始,有待进一步研究。
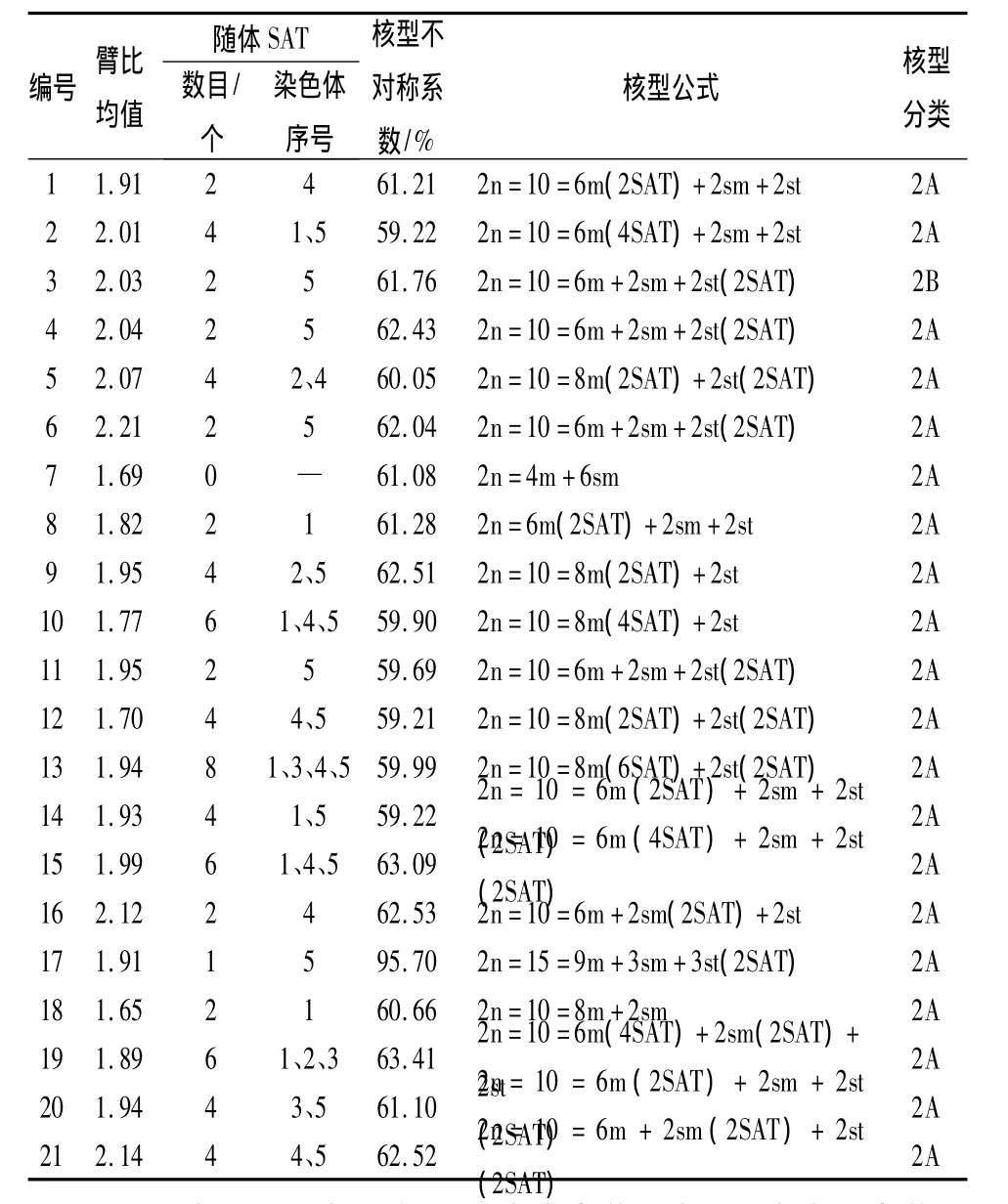
表2 21个中原传统牡丹品种的主要核型参数
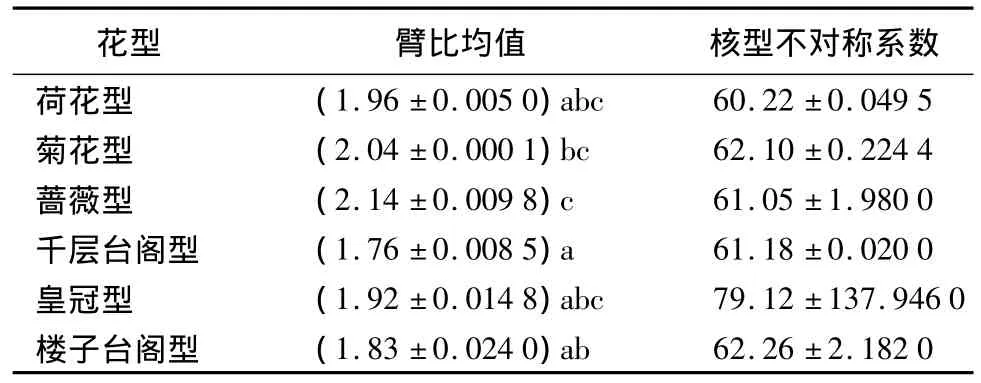
表3 6个花型染色体臂比均值、核型不对称系数方差分析及多重比较
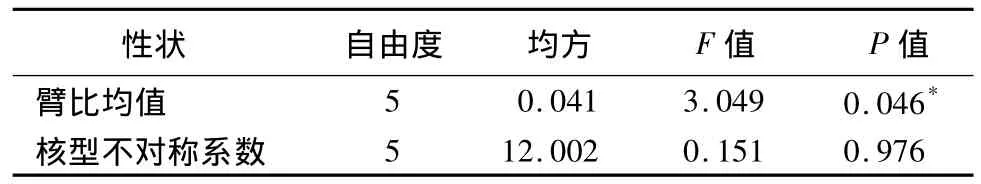
表4 6个花型间染色体臂比均值、核型不对称系数方差分析
传统品种花型间的核型不对称系数差异不显著,由低到高依次为:荷花型、蔷薇型、千层台阁型、菊花型、楼子台阁型、皇冠型,这与王莲英[1]14-19提出牡丹品种按花型演进的途径的观点基本一致,而试验中的菊花型和千层台阁型的品种没有遵循传统的花型演进途径,极有可能是因为这2个花型的品种栽培年代晚一些或栽培条件不同。传统品种花型内部的核型不对称系数差异较大,从小到大依次为千层台阁型、菊花型、荷花型、蔷薇型、楼子台阁型和皇冠型,说明千层台阁型品种的核型类型较一致,而皇冠型品种的核型类型差异最大。

图3 21个中原传统牡丹品种核型似近系数聚类图
臂比均值体现的是一个核型内所有染色体整体的不对称性。观察的21个品种中‘赤龙焕彩’臂比均值(1.65)最小,核型较对称,跟核型类型分析的结果一致,也证明了其原始性。还发现各花型之间的臂比均值差异显著,菊花型的臂比均值较大,说明该花型的核型不对称性。通过各花型的臂比均值方差比较得到菊花型的核型类型最一致,楼子台阁型和皇冠型一致性最差,与核型不对称系数得出的结论基本吻合,表明材料中的菊花型品种进化程度较高。
依据数值分类学原理和似近分析理论得出的核型似近系数及核型进化距离能够多向、立体、多维地考察物种间的相似性,从而判断物种间的亲缘关系和遗传距离[2]。目前核型似近系数聚类分析方法应用较少,仅分析过蝗虫、鸡、禽类、猪、牦牛、榛子、风毛菊属植物、紫花苜蓿、青稞和大花蕙兰,发现聚类结果与品种分化历史基本一致[2,22-30]。文中对21个中原牡丹传统品种的亲缘关系与进化距离进行了分析,结果与传统分类结果不完全相同。似近系数为0.850 0以上,进化距离间隔小,这说明了21个中原牡丹传统品种在核型上稳定性和相似性很高。同属于皇冠型的‘豆绿’和‘姚黄’,‘蓝田玉’和‘赵粉’亲缘关系最近,遗传背景相似。在进化距离为0.014 0、似近系数为 0.9861 时,‘黄花魁’、‘桃红献媚’、‘大棕紫’和‘黑花魁’4个品种聚成一类。而这4个品种分属于4个不同的花型。属于托桂型的‘青龙卧墨池’和属于蔷薇型的‘锦袍红’在似近系数为0.975 6时聚为一类。‘首案红’在似近系数0.960 3处自成一组,这充分说明其在进化过程中的特殊性。文中的研究结果显示,在细胞学水平上大致符合牡丹组依据花型分类的结果,可能原因是由于这些传统品种栽培年代、条件不同,也许经品种杂交选育,花型容易发生变异,核型分析中的人为误差以及核型似近系数聚类分析方法的适用性等。因此,中原牡丹传统品种分类和鉴定问题及细胞学与形态学之间的关联还需进一步验证。
[1]王莲英.中国牡丹品种图志[M].北京:中国林业出版社,1997.
[2]徐琪,陈国宏,张学余,等.3个地方鸡种的核型及其似近系数分析[J].畜牧兽医学报,2004,35(4):362 -366.
[3]谭远德,吴昌谋.核型似近系数的聚类分析方法[J].遗传学报,1993,10(4):21 -25.
[4]李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996:287.
[5]李懋学,张效方.三倍体牡丹的细胞遗传学观察[J].遗传,1982,4(5):19 -21.
[6]张赞平.栽培牡丹的核型研究[J].豫西农专学报,1988(2):5-12.
[7]杨涤清,朱燮桴.草芍药、野牡丹和黄牡丹的核型研究[J].云南植物研究,1989,11(2):139 -144.
[8]李思峰,于兆英,周俊彦.黄牡丹的核型分析[J].武汉植物研究,1989,7(2):107 -111.
[9]肖调江,龚洵,夏丽芳.滇牡丹复合群的Giemsa C-带比较研究[J].云南植物研究,1997,19(4):395 -401.
[10]裴颜龙.牡丹复合体的研究[D].北京:中国科学院植物研究所,1993:57.
[11]张赞平,侯小改.杨山牡丹的核型分析[J].遗传,1996,18(5):3-6.
[12]于兆英,李思峰,周俊彦.珍稀植物:紫斑牡丹和矮牡丹的核型分析[J].西北植物学报,1989,7(1):12 -16.
[13]洪德元,张志宽,朱相云.芍药属研究(Ⅰ):国产几个野生种核型报道[J].植物分类学报,1988,26(1):33 -43.
[14]王莲英,刘淑敏,秦魁杰,等.牡丹及其栽培品种的染色体组型[J].北京林学院学报,1983(1):63 -69.
[15]张赞平,李懋学,袁甲正.牡丹染色体的Ag-NoR和Giemsa C-带研究[J].武汉植物学研究,1990,8(2):101-105.
[16]于玲,何丽霞,李嘉珏.甘肃紫斑牡丹与中原牡丹类群染色体比较研究[J].园艺学报,1997,24(1):79 -83.
[17]侯小改,段春燕,刘素云,等.中国牡丹染色体研究进展[J].中国农学通报,2006,22(2):307 -309.
[18]Levan A,Fredga K,Sandberg A A.Nomenclature for centromere position on chromosome[J].Hereditas,1964,52(3):201 -220.
[19]Stebbins G L.Chromosome evolution in higher plants[M].London:Academicn Press,1971:87 -123.
[20]Arano H.Cytological studies in subfamily Carduoideae of Japan IX[J].Bot Mag,1963,76:32.
[21]吴昌谋.核型似近系数和进化距离的估计[J].动物分类学报,1996,21(3):338 -344.
[22]姚世鸿,吴昌谋.三科五属18种蝗虫的核型似近系数聚类分析研究[J].贵州师范大学学报:自然科学版,2004,22(3):19 -24.
[23]熊家军.七种禽类的核型似近系数聚类分析和鸡W染色体相对长度与其主要经济性状的相关分析[D].武汉:华中农业大学,2002.
[24]吴圣龙,包文斌,束靖婷,等.安徽地方猪种间核型似近系数和进化距离的聚类分析[J].中国畜牧兽医,2006,42(21):4-6.
[25]钟金诚,陈智华,张成忠.牦牛品种(类群)间核型似近系数分析[J].草食家畜,1996(3):11-13.
[26]郭媛媛,邢世岩,马颖敏,等.15种榛子种质的染色体核型分析[J].园艺学报,2009,36(1):27 -32.
[27]乔永刚,宋芸.15种风毛菊属植物核型似近系数聚类分析[J].中国草地学报,2011,33(2):38 -43.
[28]张雪婷,杨文雅,杨芳萍,等.6个紫花苜蓿材料的核型及其亲缘关系分析[J].西北植物学报,2011,31(4):671 -676.
[29]段瑞君,任又成,熊辉岩.4种青海青稞品种的染色体核型分析[J].中国农学通报,2010,26(15):78 -82.
[30]王利民,王四清,董晓宇,等.核型似近系数在大花蕙兰杂交育种中的应用初探[J].河南农业科学,2010(8):106-113.
猜你喜欢
黄河之声(2021年23期)2021-06-21
宁夏医学杂志(2020年3期)2021-01-21
人大建设(2019年10期)2019-12-27
小资CHIC!ELEGANCE(2019年32期)2019-11-22
人大建设(2019年3期)2019-07-13
咸阳师范学院学报(2018年4期)2018-09-13
知识经济·中国直销(2017年12期)2018-01-03
哈尔滨医药(2015年2期)2015-12-01
中国当代医药(2015年9期)2015-03-01
应用海洋学学报(2014年3期)2014-11-22
