
不同土壤养分水平下2种外来鬼针草和近缘本地种的比较研究
2012-07-05 03:43刘明超韦春强唐赛春潘玉梅广西壮族自治区中国科学院广西植物研究所广西桂林54006广西师范大学生命科学学院广西桂林54004
生物安全学报 2012年1期
刘明超,韦春强,唐赛春,潘玉梅广西壮族自治区中国科学院广西植物研究所,广西桂林54006;广西师范大学生命科学学院,广西桂林54004
外来物种入侵不仅造成生物多样性降低,而且严重威胁全球的生态环境和经济发展(Pimentel et al.,2005),受到各国政府、专家、学者的广泛关注和重视。因此,生物入侵问题已成为全球研究的热点。就入侵植物而言,国内外学者从其形态、生长、生理和繁殖性状等方面开展了广泛的研究,旨在揭示外来植物的入侵能力和入侵性(Barret,2008;Gurevitch et al.,2008;Singh et al.,2011),以实现对外来植物有效的控制和管理。虽然目前已对外来入侵植物的特性进行了较多研究,并发现外来入侵植物具有某些非入侵植物所不具有的功能性状,但仍无法对外来植物的入侵性进行有效预测和评价(刘建等,2010;Hawkes,2007;van Kleune et al.,2010)。
比较亲缘关系较近的入侵种和本地种是研究植物入侵性的重要方法(王坤等,2009)。将入侵种与其近缘本地种或入侵能力较弱的外来种进行比较能够排除亲缘关系带来的影响,使结果更具说服力(王坤等,2009;Feng&Fu,2008)。此方法已受到越来越多学者的重视(王坤等,2010;Feng&Fu,2008;Geng et al.,2006;Leicht-Young et al.,2007;Zheng et al.,2009)。已有的比较研究结果为揭示外来植物的入侵机制提供了重要依据,但目前,入侵种与近缘种的比较研究资料仍然缺乏,影响了对外来植物入侵性的有效预测和评价。
鬼针草属三叶鬼针草Bidens pilosa L.var.radiata Sch.-Bip.和白花鬼针草B.alba L.原产于热带美洲,主要分布于路旁、耕地、弃耕地、树林下、城镇周边的垃圾场等生境。入侵我国后给生态环境、农业经济发展等造成严重的危害和影响。由于对这2种植物的分类鉴定存在许多困难,有关它们的入侵性和危害至今仍未引起人们重视。郝建华等(2009)、洪岚等(2004)、王瑞龙等(2011)、徐玲明和林训仕(2005)、叶子飘和赵则海(2009)从生长、种子萌发、繁殖、化感作用等方面对三叶鬼针草开展了研究。但目前大多数的研究主要针对单个物种。为了更好地理解三叶鬼针草和白花鬼针草的入侵能力和入侵机制,需要深入开展入侵种与近缘本地种之间的比较研究。
鉴于三叶鬼针草和白花鬼针草入侵的生境可能具有不同的土壤养分水平,本文设置低肥和高肥2个水平,通过盆栽控制试验,比较研究三叶鬼针草和白花鬼针草与近缘本地种金盏银盘B.biternata L.在不同土壤养分水平下的生长状况及其对土壤养分的可塑性,探讨不同土壤养分条件下三叶鬼针草和白花鬼针草的入侵能力和入侵性,为揭示三叶鬼针草和白花鬼针草的入侵机制和影响因素提供依据,并为入侵预测和控制提供基本资料。
1 材料与方法
1.1 材料与试验地
三叶鬼针草和金盏银盘种子分别于2010年11月和8月采自广西桂林地区,白花鬼针草种子于11月采自广西横县。试验在广西植物研究所试验地大棚内进行,地点位于广西桂林市雁山区(25°4'35″N、110°18'20″E),海拔约160 m,属中亚热带季风气候。
1.2 试验设计
于2011年4月12日,将3种植物的种子分别播种于12个花盆内;5月9日株高约15 cm时,选择各种植物大小较一致的幼苗移栽到内径23 cm、深18 cm的塑料花盆中(移栽土壤为耕地土壤和贫瘠黄泥土以1∶1体积混合);2周后,各植株均已长出新根,开始进行试验。试验设高肥与低肥2个梯度处理,高肥处理为每周每盆施加4 g“施丰源(硫酸钾型)”复合肥,低肥处理每周每盆施加0.3 g“施丰源”复合肥。试验过程中,每天浇足水,以保证足够的水分使其生长。每个物种每个养分梯度种植15盆,每盆1株,其中5盆用于施肥处理前原始株高、生物量、叶面积等参数的测定,5盆用于营养生长旺盛期生长参数的测定,5盆用于繁殖持续期和种子产量的测定。3种植物材料共种植90盆。
5月20日各处理选择生长较一致的5株测定原始株高(0.1 cm钢卷尺)、生物量、总叶面积(CI-203便携激光叶面积仪),6月26日植物达到营养生长最旺盛时期,有少数植株开始出现花蕾,此时各处理随机选取5株,测定其株高、分枝数、叶面积,并在智能烘箱80℃下烘干至恒重后称重,测定根、茎、叶的干重。并计算如下参数:比叶面积(总叶面积/总叶重)、叶面积比(总叶面积/植株总重)、根生物量比(根重/植株总重)、茎生物量比(茎重/植株总重)、叶生物量比(叶重/植株总重)。
采用Poorter(1999)的方法计算平均相对生长速率(relative growth rate,RGR)、净同化速率(net assimilation rate,NAR)和平均叶面积比(mean leaf area ratio,LARm)。
繁殖期,记录各植株始花期、盛花期和末花期;统计每植株产生的头状花序数,并随机选择20个头状花序,统计种子数,计算单株种子量 (单株头状花序数×平均单个头状花序的种子数)。根据Valladares et al.(2000)的方法计算各生长参数的可塑性指数。
1.3 数据分析
统计计算和方差分析均采用SPSS 13.0统计软件进行。物种和养分对各生长参数的影响用twoway ANOVA分析;同一物种不同养分间的差异以及相同养分水平下不同物种间的差异用one-way ANOVA分析,处理间差异显著水平为0.05。用EXCEL 2003软件做图。
2 结果与分析
2.1 物种、养分对各生长参数的影响
物种除了对株高以及根、茎、叶生物量比无明显影响外,对其余参数均有显著影响;养分除对叶生物量比、净同化速率、繁殖持续期无明显影响外,对其余生长参数均有显著影响;物种与养分的交互作用对分枝数、总叶面积、比叶面积、总生物量、种子产量均具有显著影响(表1)。通过F值可知,养分对大多数生长特征的作用最大,物种对净同化速率、繁殖持续期的作用最大。
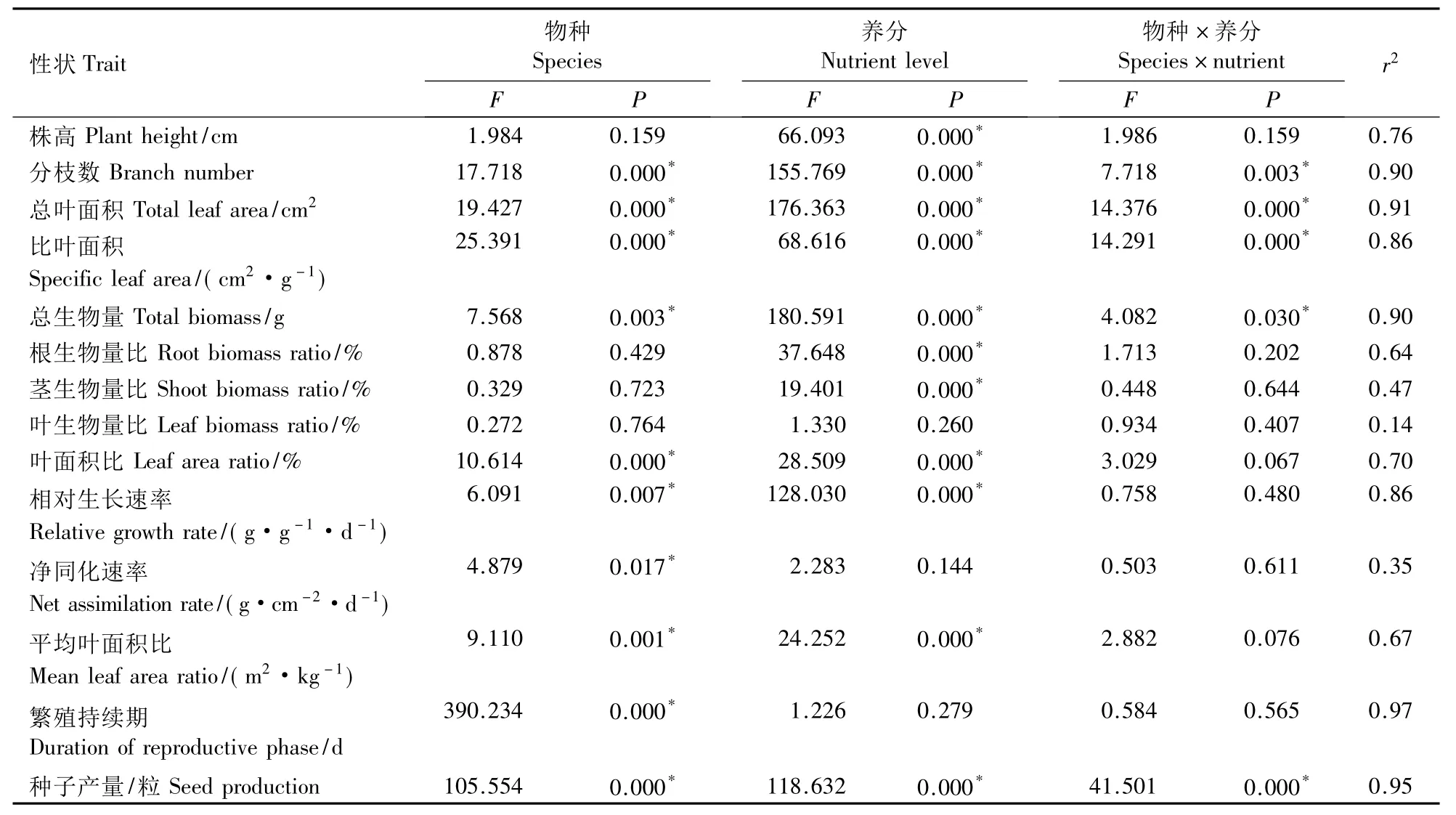
表1 物种(n=3)、养分(n=2)对3种植物生长特征的影响Table 1 Results of a two-way ANOVA analysis of the species(n=3)and nutrient levels(n=2)on traits of B.pilosa var.radiata,B.alba and B.biternata
2.2 养分水平对3种植物形态特征的影响
3种植物在高肥条件下的株高明显高于低肥条件下(P<0.05)(图1)。在低肥条件下,三叶鬼针草和本地种金盏银盘的株高无明显差异(P>0.05),白花鬼针草的株高明显低于前两者(P<0.05);但在高肥条件下,外来种和本地种的株高无显著差异(P>0.05)(图1)。在低肥和高肥条件下,金盏银盘的分枝数明显高于2个外来种 (P<0.05)(图1)。在低肥条件下,3种植物的总叶面积和比叶面积无明显差异(P>0.05);高肥条件下,白花鬼针草的总叶面积和比叶面积最高,金盏银盘次之,三叶鬼针草最低,3种植物间差异显著 (P<0.05)(图1)。

图1 不同养分条件下三叶鬼针草、白花鬼针草和金盏银盘的形态特征Fig.1 Morphological traits of B.pilosa var.radiata,B.alba and B.biternata grown under different soil nutrient levels
2.3 养分水平对3种植物生物量及其分配的影响
在低肥条件下,3种植物的总生物量无显著差异(P>0.05);在高肥条件下,白花鬼针草生物量最大,金盏银盘次之,三叶鬼针草最小,并与前两者差异显著(P<0.05)。无论是在低肥条件下还是在高肥条件下,3种植物之间根、茎、叶生物量分配均无明显差异 (P>0.05)。对于2种入侵种来说,低肥条件下根生物量分配明显大于高肥条件下(P<0.05)。低肥与高肥条件之间,3种植物叶的生物量分配均无明显区别(P>0.05)。低肥条件下,3种植物叶面积比无显著差异 (P>0.05);但在高肥条件下,白花鬼针草的叶面积比明显高于其他2种植物(P<0.05),而三叶鬼针草与本地种之间无明显差异(P >0.05)(图2)。
2.4 养分水平对3种植物生长的影响
低肥条件下,3种植物的相对生长速率、净同化速率以及平均叶面积比无显著差异(P>0.05);但在高肥条件下,金盏银盘和白花鬼针草的相对生长速率显著高于三叶鬼针草(P<0.05);白花鬼针草的净同化速率低于金盏银盘和三叶鬼针草,并与金盏银盘有明显差异(P<0.05);白花鬼针草的平均叶面积比明显高于金盏银盘和三叶鬼针草(P<0.05)(图3)。

图2 不同养分条件下三叶鬼针草、白花鬼针草和金盏银盘的生物量及其分配Fig.2 Biomass and biomass allocation of B.pilosa var.radiata,B.alba and B.biternata grown under different soil nutrient levels

图3 不同养分条件下三叶鬼针草、白花鬼针草和金盏银盘的生长特征Fig.3 Growth traits of B.pilosa var.radiata,B.alba and B.biternata grown under different soil nutrient levels
2.5 养分水平对3种植物繁殖特征的影响
3种植物的繁殖持续期在低肥和高肥条件下无明显差异 (P>0.05)。但2种外来种的繁殖持续期在不同土壤养分下均显著高于本地种金盏银盘(P<0.05)。高的土壤养分明显促进了3种植物的种子产量(P<0.05)。白花鬼针草的种子产量在低肥和高肥条件下均最高,三叶鬼针草的种子产量次之,本地种金盏银盘种子产量最少;且白花鬼针草与其他2种差异显著(P<0.05)(图4)。
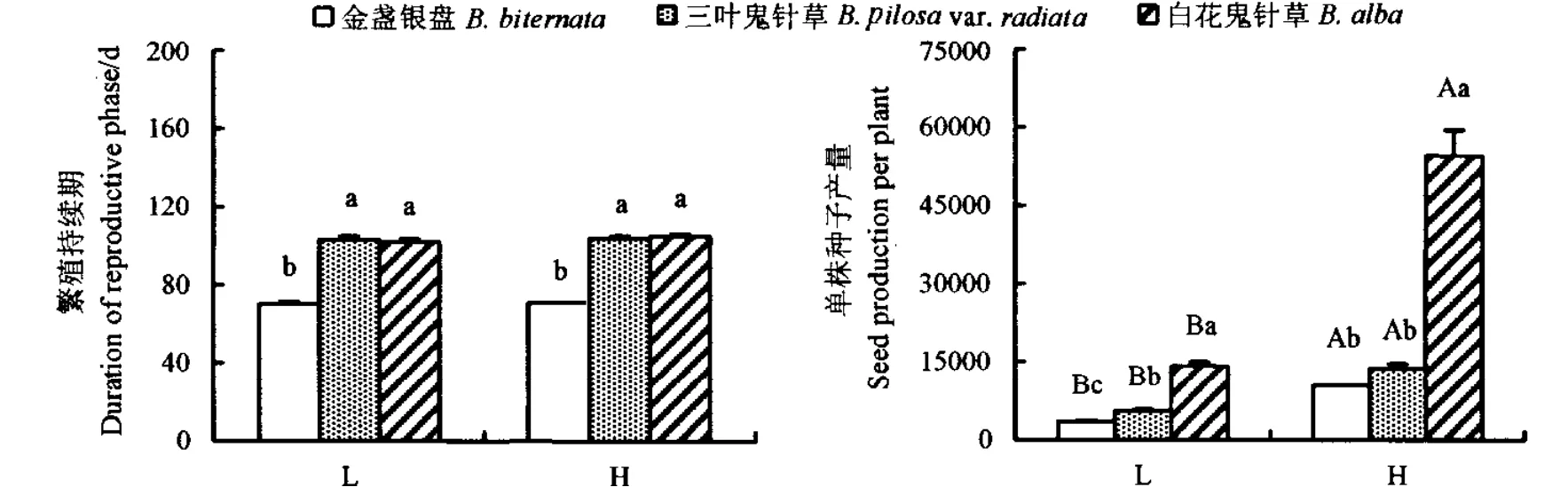
图4 不同养分条件下三叶鬼针草、白花鬼针草和金盏银盘的繁殖持续期和种子产量Fig.4 Duration of reproductive phase and seed production of B.pilosa var.radiata,B.alba and B.biternata grown under different soil nutrient levels
2.6 3种植物的生长状况对养分水平的表型可塑性
3种植物的大多数参数对土壤养分均具有较高的可塑性,尤其是分枝数、总叶面积、总生物量和种子产量(表2)。白花鬼针草在株高、总叶面积、比叶面积、叶面积比、总生物量、相对生长速率、平均叶面积比和种子产量方面的可塑性高于其他2种植物。三叶鬼针草与金盏银盘相比,大多数参数的可塑性指数较接近。

表2 三叶鬼针草、白花鬼针草和金盏银盘形态和生长的可塑性指数Table 2 Phenotypic plasticity index for traits related to the morphology and growth in B.pilosa var.radiata,B.alba and B.biternata grown under different soil nutrient levels
3 讨论
3.1 形态和生长特征与2种外来鬼针草的入侵性分析
生境中土壤养分的高低对植物的形态和生长性状具有重要影响。人类活动,如施肥、倾倒各种含养分的垃圾等,使土壤中的养分含量增加。养分增加对3种同属植物的株高、分枝数、总叶面积、比叶面积、生物量、相对生长速率、平均叶面积比、种子产量等具有显著影响(表1)。高养分条件下,3种植物大部分生长参数均明显高于低养分条件下(图1~4)。植物的形态和生长性状可影响植物的定居建群和竞争能力,从而影响植物的入侵性(刘建等,2010)。
与本地种相比,外来物种一般具有较高的相对生长速率、比叶面积、叶面积比和最大净光合速率(Pyšek et al.,2009;Williamson & Fitter,1996;Williams et al.,1995)。但在本研究中,无论是低肥还是高肥条件下,白花鬼针草的相对生长速率与本地种金盏银盘均相近,而三叶鬼针草的相对生长速率却低于甚至显著低于(高肥条件下)金盏银盘(图3)。3种植物的相对生长速率高于同科其他入侵植物,如紫茎泽兰Eupatorium adenophorum Spreng.、飞机草 E.odoratum L.等(王满莲和冯玉龙,2005)。对于总叶面积、比叶面积和叶面积比来说,在低肥条件下3个物种之间没有显著差异,但在高肥条件下白花鬼针草显著高于金盏银盘,而三叶鬼针草却显著低于金盏银盘(图1)。
Burns(2004)对鸭跖草科外来种和本地种的研究表明,高养分水平下外来种的生物量高于本地种,但在低养分水平下,两者没有差别。本研究中,低养分条件下,3种植物的生物量没有明显差别,但高养分水平下与鸭跖草科入侵种和本地种的状况不同,本地种金盏银盘的生物量与外来种白花鬼针草相近,且明显高于外来种三叶鬼针草(图2)。在生物量分配策略上,3种植物在相同养分条件下的分配策略相似,低肥时,根生物量比增大,提高对养分的吸收;但养分变化对3种植物叶生物量分配没有影响(图2、表1)。
从营养生长来说,高养分条件下白花鬼针草总叶面积、比叶面积、叶面积比明显高于金盏银盘,而三叶鬼针草的多数生长参数都低于金盏银盘,这可能由于金盏银盘本身也是一种适应能力较强的本地杂草。但从繁殖生长来看,2种外来种在低肥和高肥条件下都具有比近缘本地种长的繁殖持续期和高的种子产量,尤其是在高养分条件下种子产量更高(图4),且种子萌发率高,有利于形成白花鬼针草和三叶鬼针草的优势群落,从而促进其入侵。Barret et al.(2008)研究表明,繁殖能力与外来植物的入侵能力呈正相关。许多近缘种比较研究也证实,与繁殖相关的性状的绝对优势是外来物种成功入侵的重要原因(王坤等,2009;Walck et al.,2001)。因此,强大的繁殖能力可能是区别2个入侵种与近缘本地种的重要特征。
3.2 表型可塑性与2种外来鬼针草的入侵性分析
表型可塑性指入侵植物通过改变形态、生长、生物量分配和生理特性等获取资源,增强其对多样环境的耐受性和适应性,使其快速生长和繁殖,占据生境,增强入侵能力(Hulme,2008;Prentis et al.,2008;Richards et al.,2006)。3个物种的多数参数都具有较高的可塑性指数(表2),比较而言,白花鬼针草重要生长参数如株高、总叶面积、比叶面积、总生物量、叶面积比、相对生长速率、单株种子产量的可塑性指数均高于其他2种,说明白花鬼针草对土壤养分水平的适应性比其他2种强,既能占据高养分的生境,也能占据低养分的生境。而金盏银盘、三叶鬼针草多数参数的可塑性指数较接近,说明这2种植物的适应性较相似。在野外条件下金盏银盘作为一种杂草,有时分布范围与三叶鬼针草相同,可能与这2种植物多数参数的表型可塑性相近有关。表型可塑性是预测植物入侵的重要指标,大约50%的入侵植物的入侵性与其表型可塑性有关(刘建等,2010;Ren& Zhang,2009)。本研究中,白花鬼针草表型可塑性最高,说明其具有很强的入侵性。而三叶鬼针草的表型可塑性与本地杂草相近,可能预示三叶鬼针草的表型可塑性受其他因素的影响更大。
3.3 白花鬼针草、三叶鬼针草的管理和控制建议
外来物种的入侵性和生境的可入侵性是入侵生物学的2 个核心问题(Alpert et al.,2000;Pyšek et al.,2009)。白花鬼针草和三叶鬼针草常入侵人为干扰的生境,如耕地、弃耕地、城镇周边的垃圾场等,这些生境中一般都含有较高的养分,而入侵植物对土壤养分具有较强的适应性,使其极易成功入侵。尤其是白花鬼针草,虽然目前的分布范围还不及三叶鬼针草广泛,但其具有比本地种金盏银盘和入侵种三叶鬼针草更高的生长特征和表型可塑性,预示该植物分布范围可能进一步扩张。
从控制入侵种来说,大多数入侵植物在资源丰富的生境中入侵性较大,可以考虑降低水分和养分来削弱或抑制物种的入侵性(Burns,2006;Garcia-Serrana et al.,2007)。对于白花鬼针草和三叶鬼针草来说,降低生境中的养分水平,可以降低其大多数生长参数,从而削弱其入侵能力,在一定程度上实现控制。
郝建华,刘倩倩,强胜.2009.菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系.植物学报,44(6):656-665.
洪岚,沈浩,杨期和,曹洪麟,叶万辉.2004.外来入侵植物三叶鬼针草种子萌发与贮藏特性研究.武汉植物研究,22(5):433-437.
刘建,李钧敏,余华,何维明,于飞海,桑卫国,刘国方,董鸣.2010.植物功能性状与外来植物入侵.生物多样性,18(6):569-576.
王坤,杨继,陈家宽.2009.近缘种比较研究在植物入侵生态学中的应用.生物多样性,17(4):353-361.
王坤,杨继,陈家宽.2010.不同土壤水分和养分条件下喜旱莲子草与同属种生长状况的比较研究.生物多样性,18(6):615-621.
王满莲,冯玉龙.2005.紫茎泽兰和飞机草的形态、生物量分配和光合特性对氮营养的响应.植物生态学报,29(5):697-705.
王瑞龙,韩萌,梁笑婷,宋圆圆,苏贻娟,曾任森.2011.三叶鬼针草生物量分配与化感作用对大气温度升高的响应.生态环境,20(6-7):1026-1030.
徐玲明,林训仕.2005.三种鬼针草植株、种子外观形态及发芽率之比较.中华民国杂草学会会刊,26(1):33-42.
叶子飘,赵则海.2009.遮光对三叶鬼针草光合作用和叶绿素含量的影响.生态学杂志,28(1):19-22.
Alpert P,Bone E and Holzapfel C.2000.Invasiveness,invisibility,and the role of environmental stress in preventing the spread of non-native plants.Perspectives in Plant Ecology,E-volution and Systematics,3:52-66.
Barrett S C H,Colautti R I and Eckert C G.2008.Plant reproductive systems and evolution during biological invasion.Molecular Ecology,17:373-383.
Burns J H.2004.A comparison of invasive and non-invasive dayflowers(Commelinaceae)across experimental nutrient and water gradients.Diversity and Distributions,10:387-397.
Burns J H.2006.Relatedness and environment affect traits associated with invasive and noninvasive introduced Comelinaceae.Ecological Applications,16:1367-1376.
Feng Y L and Fu G L.2008.Nitrogen allocation,partitioning and use efficiency in three invasive plant species in comparison with their native congeners.Biological Invasions,10:891-902.
Garcia S H,Sans F X and Escarre J.2007.Interspecific competition between alien and native congeneric species.Acta Oecologica,31:69-78.
Geng Y P,Pan X Y,Xu C Y,Zhang W J,Li B and Che J K.2006.Phenotypic plasticity of invasive Alteranthera philoxeroides in relation to different water availability,compared to its native congener.Acta Oecologica,30:380-385.
Gurevitch J,Howard T G,Ashton I W,Leger E A,Howe K M,Woo E and Lerdau M.2008.Effects of experimental manipulation of light and nutrients on establishment of seedlings of native and invasive woody species in Long Island,NY forests.Biological Invasions,10:821-831.
Hawkes C V.2007.Are invaders moving targets?The generality and persistence of advantages in size,reproduction,and enemy release in invasive plant species with time since introduction.The American Naturalist,170:832-843.
Hulme P E.2008.Phenotypic plasticity and plant invasions:is it all jack?Functional Ecology,22:3-7.
Leicht Y S A,Silander J A and Latimer A M.2007.Comparative performance of invasive and native Celastrus species across environmental gradents.Oecologia,154:273-282.
Pimentel D,Zuniga R and Morrison D.2005.Update on the environmental and economic costs associated with alien-invasive species in the United States.Ecological Economics,52:273-288.
Poort L.1999.Growth response of 15 rain-forest tree species to a light gradient:the relative importance of morphological and physiological traits.Functional Ecology,13:396-410.
Prentis P J,Wilson J R U,Dormontt E E,Richardson D M and Lowe A J.2008.Adaptive evolution in invasive species.Trends in Plant Science,23:288-294.
Pyšek P,Krivanek P M and Jarosik V.2009.Planting intensity,residence time,and species traits determine invasion success of alien woody species.Ecology,90:2734-2744.
Ren M X and Zhang Q G.2009.The relative generality of plant invasion mechanisms and predicting future invasive plants.Weed Research,49:449-460.
Richards C L,Bossdorf O,Muth N Z,Gurevitch J and Pigliucci M.2006.Jack of all trades,master of some?On the role of phenotypic plasticity in plant invasions.Ecology Letters,9:981-993.
Singh V,Singh H,Sharma G P and Raghubanshi A S.2011.Eco-physiological performance of two invasive weed congeners(Ageratum conyzoides L.and Ageratum houstonianum Mill.)in the Indo-Gangetic plains of India.Environmental Monitoring Assessment,178:415-422.
Valladares F,Wright S J,Lasso E,Kitajima K and Pearcy R W.2000.Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest.Ecology,21:1925-1936.
van Kleunen M,Dawson W,Schlaepfer D,Jeschke J M and Fischer M.2010.Are invaders different?A conceptual framework of comparative approaches for assessing determinants of invasiveness.Ecology Letters,13:947-958.
Walck J L,Baskin J M and Baskin C C.2001.Why is Solidago shortii narrowly endemic and S.altissmima geographically widespread?A comprehensive comparative study of biological trait.Journal of Biogeography,28:1221-1237.
Williams D G,Mack R N and Black R A.1995.Ecophysiology and growth of introduced Pennisetum setaoeum on Hawaii:the role of phenotypic plasticity.Ecology,76:1569-1580.
Williamson M and Fitter A.1996.The varying success of invaders.Ecology,77:1661-1666.
Zheng Y L,Feng Y L,Liu X W and Liao Z Y.2009.Growth,biomass allocation,morphology,and photosynthesis of invasive Eupatorium adenophorum and its native congeners grown at four irradiances.Plant Ecology,203:263-271.
猜你喜欢
食品研究与开发(2021年19期)2021-11-17
雪豆月读·低年级(2021年1期)2021-09-10
河北画报(2020年19期)2020-06-16
大理文化(2020年3期)2020-06-11
读写月报(初中版)(2019年9期)2019-09-10
快乐语文(2018年34期)2018-03-06
中成药(2018年1期)2018-02-02
作文周刊·小学三年级版(2017年15期)2017-06-22
大自然探索(2015年5期)2015-09-10
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
