
毛竹笋快速生长过程中冠层参数动态及其与光合有效辐射的关系
2012-07-30 10:40陆国富杜华强周国模吕玉龙谷成燕商珍珍
浙江农林大学学报 2012年6期
陆国富,杜华强,周国模,吕玉龙,谷成燕,商珍珍
(1.浙江农林大学 浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安 311300;2.浙江农林大学 环境与资源学院,浙江 临安 311300;3.浙江省安吉县林业局,浙江 安吉313300)
森林冠层担负着整个森林生态系统物质能量的传输、太阳辐射的传递以及维护环境因子、生物参数等在空间上的分布状态而具有重要的功能[1]。冠层参数包括生物物理参数(如叶面积指数、净第一性生产力等),结构参数(如冠层郁闭度、冠层高度等),以及决定植被光谱反射特征的生物化学参数(如叶绿素、水、氮等)等[2-5],是区域乃至全球生态环境变化的重要指示,也是生态模型、碳循环、生物多样性等研究领域中的重要特征参量[6]。光合有效辐射是太阳辐射光谱中可被绿色植物的质体色素吸收、转化并用于合成有机物质的400~700 nm波段的辐射能,是林木营养生长阶段的主要因素,其分布特征直接影响林木的光合作用[7]。在一定的环境条件下,林木光合生产量取决于叶片吸收的光合有效辐射和叶片的光合特性,而冠层内的辐射状况又表现出空间上的异质性和时间上的动态特性,同时又受到叶面积指数、叶片的形状和大小、叶片散射、吸收和反射以及这些特征随树木种类、生长发育阶段和林分密度的影响[8]。可见,林内光辐射与森林生产力、冠层结构紧密相关,而林冠结构、辐射传输与冠层光合作用三者模拟也一直是树木生理生态学的一个热点和难点[7-8]。森林固定二氧化碳的过程实际上就是植物利用太阳光能将二氧化碳和水转化为碳水化合物的过程[5],冠层结构在很大程度上影响并决定了水、热、光等生态环境因子在林内的传递和再次分配,从而影响植被碳积累。因此,森林冠层参数动态变化及其与光合有效辐射的关系对深入研究森林碳动态具有重要意义。毛竹Phyllostachys edulis林是中国亚热带地区特殊的森林类型,其碳汇功能得到广泛认可[9]。毛竹具有速生特点,新竹从出笋到长成只需50 d稍多时间,此后开始抽枝长叶,且生物量碳储量快速积累。然而,在这一过程中,毛竹冠层动态及其与光合有效辐射之间的关系尚不清楚,在一定程度上影响了毛竹碳积累过程的理解。本研究将分竹笋展枝放叶前后2个阶段,观测毛竹林生长过程中光合有效辐射(photosynthetic active radiation,W·m-2),单位面积叶绿素质量(total chlorophyll content,μg·cm-2),叶面积指数(leaf area index)和郁闭度(canopy closure)等冠层参数,并分析单位面积叶绿素质量、叶面积指数和郁闭度动态变化及其与光合有效辐射之间的关系,研究结果将为评价毛竹固碳机制及碳循环过程提供理论参考,为竹林碳同化遥感-光合机制模型构建提供依据。
1 研究方法
1.1 研究区概况
研究区选在浙江省安吉县山川乡。山川乡位于安吉县南端,东接余杭、南邻临安,西北与天荒坪镇接壤。山川乡境内山青水秀,环境宜人,年平均气温为14.7℃,雨水充沛,年降水量为1 700 mm;竹类资源十分丰富,被喻为竹乡中的竹乡。
1.2 数据获取与处理
1.2.1 实验设计 山川乡设有毛竹林碳通量塔。为配合通量数据,本研究在通量塔周围约900 m2范围阳坡内随机选取40株毛竹,并测量单位面积叶绿素质量;叶面积指数和郁闭度采用WinSCANOPY植物冠层分析仪获取。毛竹3月下旬开始出笋,新竹50 d稍多时间完成高度生长,到7-8月冠型基本稳定。为此从2011年4月初到7月底,间隔1周观测1次,用以分析毛竹林快速生长过程冠层参数动态变化。通量塔有7层梯度系统和二氧化碳闭路廓线系统,可采集土壤温度、湿度、热通量,以及15 m处天空向上、向下净辐射等数据。毛竹冠层生长普遍高度为8~12 m,因此,在通量塔15 m处安装净辐射仪传感器(型号CNR4)和辐射传感器(光谱波长:短波辐射传感器为310~2 800 nm,长波辐射传感器为4 500~42 000 nm)。①冠层上方光合有效辐射。通量塔净辐射仪传感器和辐射传感器安装高度高于毛竹林的高度,因此冠层上方光合有效辐射(PAR,W·m-2)认为是无遮挡的天空中的光合有效辐射。另外,碳通量塔设置在毛竹林纯林中,受下垫面影响也较小。用式(1)计算[10]。

式(1)中,E为太阳辐射值(为长波净辐射和短波净辐射之和),由通量塔观测获取。②单位面积叶绿素质量测量。将配置25 mL叶片叶绿素溶液置于避光处36 h,直至溶质全部变为灰白色,然后采用UV-2102C/PC/PCS型分光光度计测量溶液中叶绿素a(chlorophyll a)和叶绿素b(chlorophyll b),叶绿素总量为叶绿素a与叶绿素b之和。单位面积叶绿素a,b的计算公式如下:

式(2)(3)中叶绿素 a和叶绿素 b的质量,单位为 mg·cm-2; D663,D645分别表示 663 nm,645 nm 处的吸光度值;S为叶面积(cm2),采用便携式AM300-002叶面积仪测量,仪器扫描仪对比度设为3。
1.2.2 冠层叶面积指数和郁闭度 实践证明:冠层叶面积指数ILA和郁闭度的变化与枝条和叶量的变化有关。本研究选择新竹展枝放叶时开始测量叶面积指数和郁闭度,以观测毛竹快速生长期冠层的叶面积指数和郁闭度的变化影响。研究区毛竹5月7日开始展枝,到5月28日枝条全部展开,但没有放叶,6月8日前后叶片开始展开,因此,从5月7日开始,利用WinSCANOPY冠层分析仪获取图像并计算叶面积指数和郁闭度(图1)。为保证叶面积指数和郁闭度测量精度,在通量塔周围设置5个固定观测点,用三脚架将仪器固定在离地面1.5 m高度处,连续拍摄3次图像并取均值作为该次的实测值。Win SCANOPY 的叶面积指数计算方法包括 ILA(Bonhom)-lgIC(聚集度指数),ILA(2000)-lgIC,ILA(2000G)-lgIC,ILA(sphere)-lgIC和 ILA(ellips)-lgIC。算法的基本原理是 Beer-Lamber定理。如下式:

式(4)中:I为冠层下方的光照强度,I0为冠层上方的光照强度,K为消光系数(本研究取0.5),本研究采用 ILA(2000)-lgIC计算叶面积指数。
WinSCANOPY还可以分析冠层孔隙度,冠层郁闭度等于1减去孔隙度。

图1 毛竹林冠层影像及叶面积指数和郁闭度计算Figure 1 Phyllostachys edulis forest canopy image and its analyse for LAI and canopy closure
2 结果与分析
2.1 光合有效辐射动态变化
光合有效辐射是林木营养生长阶段的主要因素,对森林植物的更新、植物生产力等具有重要作用[7]。因此,太阳辐射是基础,其能量通过光合作用进入生态系统,从而影响生态系统的物质循环和能量流动,其动态变化有助于了解竹林快速生长过程中能量消长和碳积累。图2为毛竹快速生长过程中冠层上方光合有效辐射的动态变化。因5月底和6月上、中旬天气阴雨缘故,这段时间的光合有效辐射扰动很大,故在图2中没有这段时间的光合有效辐射值。放叶前的光合有效辐射截至5月21日,而放叶后的光合有效辐射仅包括7月的数据。
由图2可以看出:4月1日到5月底,毛竹林冠层上方的光合有效辐射不断增加。这段时间竹笋快速生长,需要母竹为其提供大量的有机物质,通过光合作用合成能量也不可少,因此对光能的利用会比较强。放叶后,光合有效辐射也呈增加趋势,一方面,气温的升高对叶绿素含量有影响[11],另一方面,新竹放叶后,占据一定的冠层空间,叶片开始进行光合作用。总的来说,放叶前后的光合有效辐射平均值差异不大,均在71 W·m-2左右。
2.2 叶绿素动态及其与光合有效辐射的关系
图3a为毛竹叶绿素总量动态变化,可以看出,毛竹林快速生长期,叶绿素总含量整体呈增长趋势。对比图3a和图2发现,两者在一定程度上具有相似的变化趋势。叶绿素是光合作用的场所,需要太阳辐射提供能量,两者相似的变化说明光合有效辐射为光合作用持续提供能量,同时,母竹需要大量的叶绿素进行光合作用,为竹笋快速生长提供营养支持。
但由图3b发现,放叶后叶绿素含量变化趋势明显小于放叶前,与光合有效辐射的相关系数也由放叶前的0.57降低到0.14。事实上,如同光合有效辐射的动态分析,竹笋快速高生长结束后,母竹能量消耗很大,尽管如此,母竹叶绿素含量还会缓慢增加,并通过光合作用为下一年新竹的生长存储能量;另一方面,叶绿素含量仅为母竹叶片尺度上的,而光合有效辐射是冠层尺度上,包括了新竹和母竹对太阳辐射的利用。
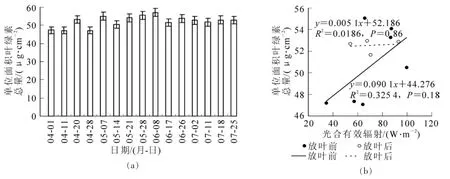
图3 毛竹林绿素总量动态变化(a)及其与光合有效辐射的关系(b)Figure 3 Dynamic change of Phyllostachys edulis TCC(a) and its relationship with PAR(b)
2.3 叶面积指数和郁闭度的动态变化及其与光合有效辐射的关系
图4~5分别是叶面积指数和郁闭度动态变化图。两者在整体上均呈增加趋势,这符合毛竹快速生长过程中结构参数动态变化的实际情况。在枝叶生长停止后达到最大,其中叶面积指数最大为4.482,郁闭度最大为0.923。另外,对比图4~5和图2发现,叶面积指数和郁闭度也与光合有效辐射具有相同的变化趋势,尤其在放叶后,叶面积指数和郁闭度与光合有效辐射是正相关关系。如图6所示。
叶面积指数定义为单位面积柱体内叶子表面积的一半[12]。叶片作为光合作用的基本器官,在光照条件下发生光合作用,产生植物干物质的积累,随着叶面积的增大,植被生物量也增大。叶面积指数可以用来反映植物叶面数量、冠层结构变化、植物群落生命活力及其环境效应,为植物冠层表面物质和能量交换的描述提供结构化的定量信息,并在生态系统碳积累。植被生产力和土壤、植物、大气之间相互作用的能量平衡等方面发挥重要作用[13-15]。随着新竹的出叶量增多,叶面积指数增加,单位面积光能利用率高,因此,叶片光合作用所需要的太阳辐射增加,反之亦然。
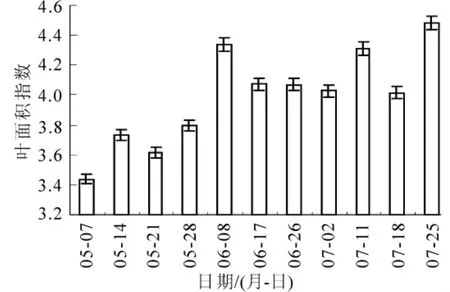
图4 叶面积指数动态变化Figure 4 Dynamic change of LAI
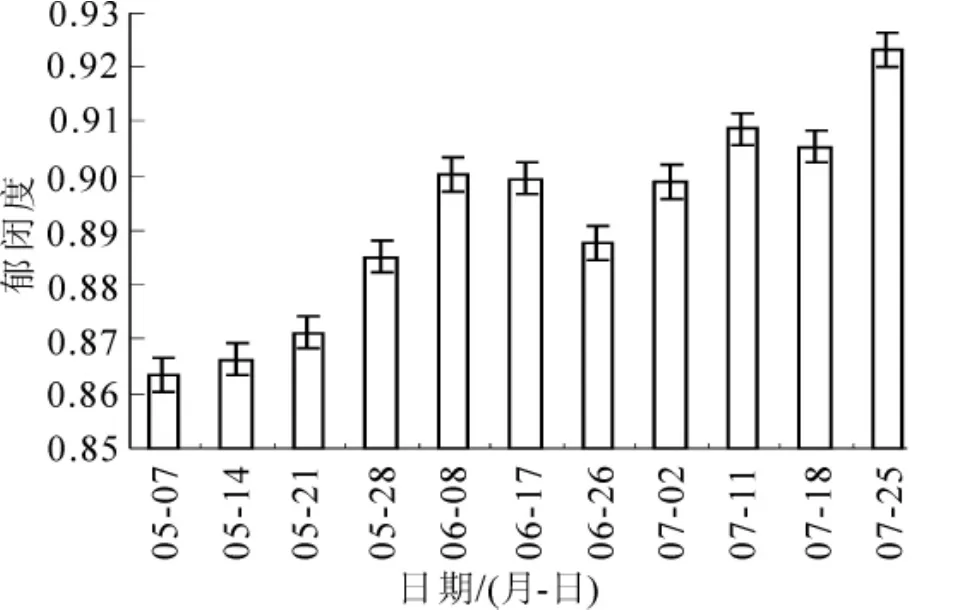
图5 郁闭度动态变化Figure 5 Dynamic change of canopy closure
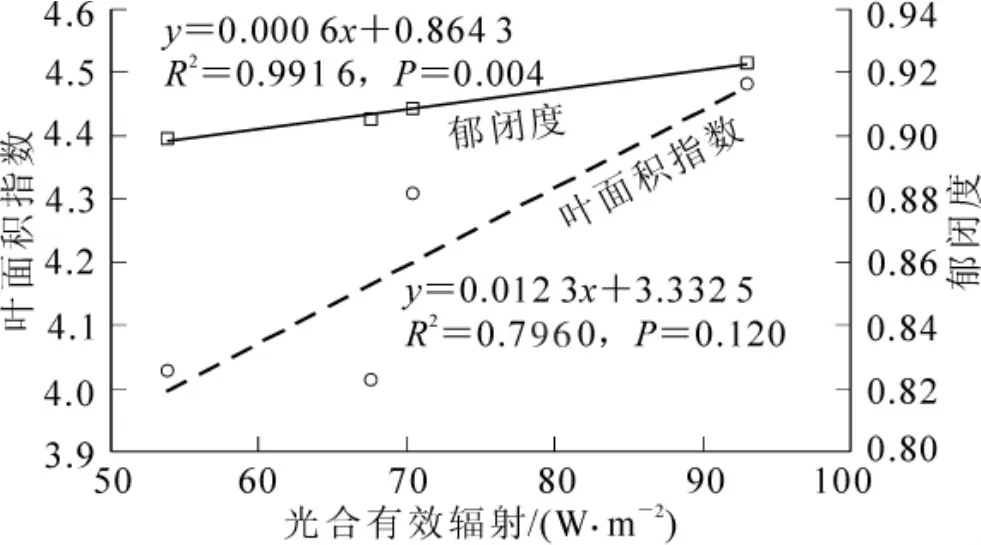
图6 放叶后叶面积指数和郁闭度与合有效辐射的相关关系Figure 6 Relationships between LAI,canopy closure,and PAR in posterior leaf development
郁闭度是决定林分结构的重要因子之一,它可以用来表征光、水等生态因子通过林冠进入林内的再分布状况[16]。本研究郁闭度与林冠层孔隙度直接相关,随着叶量增多,林冠层孔隙度减小,冠层越来越郁闭,毛竹对太阳辐射的吸收、二向反射及其辐射在毛竹林冠层的传递会加强,光能的利用也会随之增加。
3 讨论与结论
3.1 讨论
研究表明:放叶前母竹叶绿素的变化趋势及与光合有效辐射的相关性均高于展枝放叶后,但叶绿素质量为叶片尺度上,而光合有效辐射是冠层尺度上,两者之间还存在尺度转换的问题。借助PROSPECT等辐射传输模型模拟毛竹林冠层光谱,并用冠层光谱特征参数反演快速生长过程中叶绿素质量,这样就解决叶片尺度叶绿素和冠层尺度光合有效辐射之间尺度差异,可能会得到更好的结果。叶绿素动态变化为高光谱遥感反演提供基础,植被反射光谱曲线的“红边位置”“绿峰反射高度”等光谱特征参数与叶绿素含量等具有显著关系[17-18]。因此,本研究为高光谱遥感反演竹林叶绿素质量提供参考。
研究表明:毛竹展枝放叶后郁闭度逐渐增加。根据相关研究,当郁闭度大于0.8时,常用与植被碳储量遥感估算的归一化植被指数(NDVI)饱和,其对植被生物量的敏感性下降[19]。由图4可见:5月份以后毛竹林郁闭度大于0.8,可以推知归一化植被指数饱和,难以反映竹林碳储量的变化,这一结果也印证了先前的研究即毛竹林生物量与归一化植被指数相关性不显著[20]。因此,本研究连续观测得到的毛竹林冠层参数为基于遥感信息的毛竹林碳汇估算提供了重要的数据支持。在全球变化研究中,尤其是陆地生态系统碳储量和碳循环观测及过程研究中,遥感正日益成为新兴的有力研究手段[21],而植被碳同化全遥感-光合机制模型是通过分析植物光谱特征来反映植被生产力的,其生物学基础是从植被生产力形成的生理过程出发,并根据植物对太阳辐射的吸收、反射、透射及其辐射在植被冠层内及大气中的传输和植被碳同化的生态影响因子,最后在卫星接收到的信息之间建立完整的数学模型实现森林碳汇遥感监测的,这些遥感信息包括归一化植被指数、叶面积指数、吸收光合有效辐射等[3]。在这个过程中,冠层结构直接决定了太阳辐射在不同高度冠层内的分布,影响了不同层次树木平均日光合有效辐射的截获、吸收和透射,决定了林内的光分布状况[7],因此,冠层参数在森林碳汇遥感研究中至关重要。
毛竹笋展枝放叶前和展枝放叶后2个阶段毛竹林冠层参数与合有效辐射的关系不同,这为分阶段建立毛竹林碳同化遥感-光合机制模型提供了依据。叶面积指数、郁闭度是反映植被生物量或碳储量的重要参数[22-23],毛竹生物学特征决定了其快速生长阶段这些会不断变化的特点,植被指数饱和、模型多样等问题都会影响估算结果[20,24]。可见,有必要分阶段建立毛竹林碳储量遥感估算模型,以提高估算精度。
3.2 结论
本研究以毛竹林碳通量为中心,借助UV-2102C/PC/PCS型分光光度计,WinSCANOPY冠层分析仪等,连续监测了竹笋快速生长阶段毛竹林冠层参数的动态变化,并分别从展枝放叶前和展枝放叶后2个阶段分析了叶绿素质量、叶面积指数和冠层郁闭度与冠层上方光合有效辐射的关系,结果表明:①2个阶段毛竹林冠层上方光合有效辐射均有增加趋势,但放叶前后的光合有效辐射平均值差异不大,在71 W·m-2左右;②整个快速生长阶段单位面积叶绿素质量、叶面积指数和冠层郁闭度都呈增长趋势,相应的增量分别为5.5 μg·cm-2,1.0和0.6;③放叶前母竹叶绿素与光合有效辐射的相关性高于展枝放叶后两者的相关性,相关系数由0.570(P=0.180)降为0.140(P=0.860),在一定程度上反映了放叶前母竹为竹笋快速生长提供更多的光合产物;④在新竹放叶过程中,叶面积指数和冠层郁闭度与光合有效辐射的关系呈正相关,复相关系数分别为0.796 0(P=0.120)和0.991 6(P=0.004),说明这一阶段毛竹林冠层对光能的分配和利用增强。
[1]于强,谢贤群,孙菽芬,等.植物光合生产力与冠层蒸散模拟研究进展[J].生态学报,1999,19(5):744-753.YU Qiang,XIE Xianqun,SUN Shufen,et al.Andances in simulation of plant photosynthetic productivity and canopy evapotranspiration [J].Acta Ecol Sin,1999,19(5):744-753.
[2]吴炳方,曾源,黄进良.遥感提取植物生理参数LAI/FPAR的研究进展与应用[J].地球科学进展,2004,19(4):585-590.WU Bingfang,ZENG Yuan,HUANG Jinliang.Overview of LAI/FPAR retrieval from remotely sensed date [J].Adv Earth Sci,2004,19(4):585-590.
[3]张佳华,王长耀.区域归一化植被指数(NDVI)对植被光合作用响应的研究[J].干旱区资源与环境,2003,17(1):91-92.ZHANG Jiahua,WANG Changyao.The respondence of regional NDVI to plant photosynthesis [J].J Arid Land Resour Environ,2003,17(1):91-92.
[4]曾源,SCHAEPMAN M E,吴炳方,等.基于高光谱遥感数据提取森林结构参数的研究[J].遥感学报,2007,11(5):648-658.ZENG Yuan,SCHAEPMAN M E,WU Bingfang,et al.Forest structural variables retrieval using EO-1 hyperion datain combination with linear spectral unmixing and an inverted geometric-optical model[J].Remote Sens,2007,11(5):648-658.
[5]张佳华,徐祥德,延晓冬,等.多圈层陆面过程参数化研究中遥感信息应用的进展和方向[J].应用气象学报,2003,14(6):745-752.ZHANG Jiahua,XU Xiangde,YAN Xiaodong,et al.The advances and development of remote sensing of applications to land surface process parameterization [J].J Appl Meteorolog Sci,2003,14(6):745-752.
[6]徐希孺,范闻捷,陶欣.遥感反演连续植被叶面积指数的空间尺度效应[J].中国科学D辑:地球科学,2009,39(1):79-87.XU Xiru,FAN Wenjie,TAO Xin.The spatial scaling effect of continuous canopy leaves area index retrieved by remote sensing [J].Sci China Ser D,2009,39(1):79-87.
[7]邱建丽,李意德,陈德祥,等.森林冠层结构的生态学研究现状与展望[J].广东林业科技,2008,24(1):75-82.QIU Jianli,LI Yide,CHEN Dexiang,et al.The research progress and the significance of canopy structure in forest ecology [J].J Guangdong For Sci Technol,2008,24(1):75-82.
[8]张小全,徐德应,赵茂盛.林冠结构、辐射传输与冠层光合作用研究综述[J].林业科学研究,1999,12(4):411-421.ZHANG Xiaoquan,XU Deying,ZHAO Maorong.Review on forest canopy structure,radiation transfer and canopy photosynthesis [J].For Res,1999,12(4):411-421.
[9] LOU Yiping,LI Yanxia,BUCKINGHAM K,et al.Bamboo and Climate Change Mitigation [R].Beijing:INBAR,2010:1-47.
[10]毛军需.太阳辐射度与光照度换算浅析[J].河南农业科学,1995(1):11-12.MAO Junxu.The analyses of the sun radiation degrees and light conversion [J].J Henan Agric Sci,1995(1):11-12.
[11]陈桥,韩红娟,翟水晶,等.太湖地区太阳辐射与水温的变化特征及其对叶绿素a的影响[J].环境科学学报,2009,29(1):199-206.CHEN Qiao,HAN Hongjuan,ZHAI Shuijing,et al.Influence of solar radiation and water temperature on chlorophyll-a levels in Lake Taihu,China [J].Acta Sci Circumst,29(1):199-206.
[12]CHEN J M,PAVLIC G,BROWN L,et al.Derivation and validation of Canada-wide coarse-resolution leaf area index maps using high-resolution satellite imagery and ground measurements [J].Remote Sens Environ,2001 ,80:165-184.
[13]巩合德,杨国平,张一平,等.哀牢山4类植物群落叶面积指数比较.东北林业大学学报,2007,35(3):34-36.GONG Hede,YANG Guoping,ZHANG Yiping,et al.Comparison of leaf area index of four types of plant communities in Ailao Mountain [J].J Northeast For Univ,2007,35(3):34-36.
[14]王希群,马履一,张永福.北京地区油松、侧柏人工林叶面积指数变化规律[J].生态学杂志,2006,25(12):1486-1489.WANG Xiqun,MA Lüyi,ZHANG Yongfu.Changepatternsof leaf area index(LAI) of Pinus tabulaeformis and Platycladus orientalis plantations in Beijing [J].Chin J Ecol,2006,25(12):1486-1489.
[15]关德新,吴家兵,王安志,等.长白山红松针阔叶混交林林冠层叶面积指数模拟分析[J].应用生态学报,2007,18(3):499-503.GUAN Dexin,WU Jiabing,WANG Anzhi,et al.Simulation of crown leaf area index of Korean pine and broadleaved mixed forest in Changbai Mountains [J].Chin J Appl Ecol,2007,18(3):499-503.
[16]琚存勇,邸雪颖,蔡体久.变量筛选方法对郁闭度遥感估测模型的影响比较[J].林业科学,2007,43(12):333-388.JU Cunyong,DI Xueying,CAI Tijiu.Comparing impact of two selecting variables methods on canopy closure estimation [J].Sci Silv Sin,2007,43(12):333-388.
[17] SHAFRI H Z M,SALLEH M A M,GHIYAMAT A.Hyperspectral remote sensing of vegetation using red edge position techniques [J].Amer J Appl Sci,2006,3(6):1864-1871.
[18]杜华强,葛宏立,范文义等.马尾松针叶光谱特征与其叶绿素含量间关系研究[J].光谱学与光谱分析,2009,29(11):3033-3037.DU Huaqiang,GE Hongli,FAN Wenyi,et al.Study on relationships between total chlorophyll with hyperspectral features for leaves of Pinus massoniana forest[J].Spectrosc Spectr Anal,2009,29(11):3033-3037.
[19]赵英时.遥感应用分析原理与方法[M].北京:科学出版社,2006:374-375.
[20] DU Huaqiang,CUI Ruirui,ZHOU Guomo,et al.The responses of moso bamboo(Phyllostachys heterocycla var.pubescens) forest aboveground biomass to Landsat TM spectral reflectance and NDVI [J].Acta Ecol Sin,2010,30(5):257-263.
[21]牛铮,王长耀.碳循环遥感基础与应用[M].北京:科学出版社,2008.
[22]陈晓玲,赵红梅,田礼桥.环境遥感模型与应用[M].武汉:武汉大学出版社,2008.
[23]刘茂秀,史军辉,王新英,等.林分郁闭度和龄级对防护林碳汇功能及固碳价值的影响 [J].东北林业大学学报,2011,39(6):30-32.LIU Maoxiu,SHI Junhui,WANG Xinying,et al.Influences of crown density and age class on function and value of carbon sequestration of shelter forests in Jiashi County,Xinjiang [J].J Northeast For Univ,2011,39(6):30-32.
[24] HABOUDANE D,MILLER J R,PATTEY E,et al.Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies:modeling and validation in the context of precision agriculture [J].Remote Sens Environ,2004,90:337-352.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
浙江农林大学学报(2020年5期)2020-11-05
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
湖州职业技术学院学报(2020年4期)2020-03-30
福建林业(2020年5期)2020-03-18
特种经济动植物(2019年9期)2019-10-29
热带林业(2019年4期)2019-03-05
农民致富之友(2014年7期)2014-10-21
