
温度对垂穗披碱草种子萌发的影响
2012-08-20 08:01葛庆征张卫国张灵菲卫万荣
草业科学 2012年5期
葛庆征,张卫国,张灵菲,卫万荣
(兰州大学草地农业科技学院,甘肃 兰州730020)
垂穗披碱草(Elymus nutans)是高寒草甸植被群落中具有代表性的禾本科草种之一[1]。由于垂穗披碱草是高寒草甸类草地中较为恒定的组分,其优势度的变化与植被演替进程有着高度的相关性,因而,也是决定草地牧用价值及判定草地健康状况的重要因素之一[2]。目前,草地退化问题不但已经成为制约高寒牧区畜牧业发展的根本性问题,而且也是导致生态和人类生存条件恶化的重要诱因。补播适宜的草种是恢复和改良草地植被的基本措施,垂穗披碱草由于其具有本土草种的高度适应性且品质优良,因而成为高寒草地补播实践中的首选草种。然而,由于垂穗披碱草为野生草种,其种子萌发条件与典型的栽培草种存在较大差异,采用常规种子质量检验方法很难得出准确可靠的结果。目前,有关垂穗披碱草发芽条件的研究虽然也注意到了该草种的特殊性,并在变温处理、光照处理等方面有所涉及[3-4],但多数研究仍局限于常规方法[5]。即使考虑到了野生草种的特殊性,所涉及的广度和深度仍显欠缺和不足。因此,进一步探讨垂穗披碱草对最佳萌发条件的要求,不仅是科学测定和评价种子真实品质的需要,而且是补播实践中正确计算播种量、合理制订投资预算的基本依据。鉴于此,本研究仅就温度和光照条件对垂穗披碱草种子发芽的影响进行了较为深入系统的研究,以期对当前垂穗披碱草种子发芽条件的确定在理论上有所补充和完善。
1 材料与方法
1.1 供试种子 2010年9月15-25日分别在甘肃省甘南藏族自治州的玛曲欧拉(海拔3 941m)、玛曲阿孜(海拔3 528m)、夏河牙力吉(海拔3 213 m)、合作南郊(海拔2 925m)手工采集完全成熟的垂穗披碱草种子各2 000g,带回实验室作进一步清选后备用。
1.2 温度处理 恒温处理:设5、10、15、20、25和30℃共6个处理水平。用Φ12cm培养皿垫铺吸水滤纸作发芽床,每个培养皿均匀放置100粒种子,发芽期间保持滤纸湿润,在恒温、恒湿及24h光照(人工光照培养箱)培养,每日09:00将发芽种子取出,并计数,直至第30天不发芽为止。每处理均重复3次。
变温处理:以恒温处理的6个温度梯度为基准,分别降低10℃作为变温幅度,并以高、低温度在24 h内各处理12h的组合构成6个变温处理,即:-5℃/5℃、0 ℃/10 ℃、5 ℃/15 ℃、10 ℃/20 ℃、15℃/25℃和20℃/30℃。其他条件同恒温处理。
1.3 光照处理 以变温处理的6个组合为基础,只是将全天候不间断光照改设为08:00-20:00光照、20:00-08:00黑暗。其他条件同上恒温处理。
1.4 测定指标及统计分析
发芽率=最终正常发芽的种子粒数/供试种子粒数×100%;
发芽势=在7d内正常发芽的种子粒数/供试种子粒数×100%;
发芽指数(GI)=∑(Gt/Dt)。
式中,Gt为在不同时间的发芽数;Dt为相应的发芽日数。
活力指数(VI)=GI×S。
式中,S为7d内幼苗长度(cm),GI为发芽指数。
发芽周期:种子置于培养箱开始发芽之日至最后一个种子发芽之日的天数,发芽持续期统一为30d。
以上指标皆以SPSS 13.0版统计软件进行方差分析和差异显著性检验。
2 结果与分析
2.1 恒温处理 在恒温处理的6个梯度中,发芽率在5℃和30℃的温度条件下分别为74.1%和71.2%,二者间的差异不显著(P>0.05),但二者却均显著低于(P<0.05)10、15、20和25℃的4个处理,其发芽率的分布区间为80.3%~81.5%(图1)。发芽势随温度的上升先增加后下降(图2),最低值出现在最低温度(5℃,66.7%)梯度点上,最高值出现在15℃(77.5%)梯度点上,6个温度梯度间除5和30、15℃和20、10和25℃间无显著差异(P>0.05)外,其他各梯度间均存在显著差异(P<0.05)。种子活力指数在5~20℃梯度区间内表现为随温度的上升而增加,各相邻梯度间差异显著(P<0.05),20℃与25℃间无显著差异(P>0.05),但30℃梯度点却显著低于25℃(P<0.05)(图3)。发芽周期总体上表现为随温度的升高而缩短的趋势(图4),5℃时平均历时约27d,10、15℃时则降至18.7和14.7d,各温度梯度间均存在显著差异(P<0.05),其中15~30℃间,下降幅度趋于平缓,但15℃与20℃间仍存在显著差异(P<0.05),仅20与25℃、25与30℃间无显著差异(P>0.05)。

图1 恒定温条件下的发芽率变化Fig.1 Germination rate under constant temperature conditions

图2 恒温条件下的发芽势变化Fig.2 Germination power under constant temperature conditions
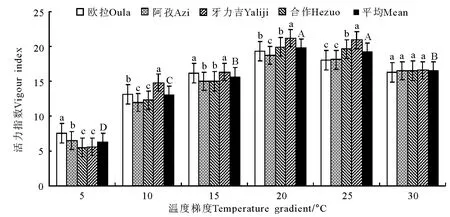
图3 恒温条件下的活力指数变化Fig.3 Vigour indexes under constant temperature conditions
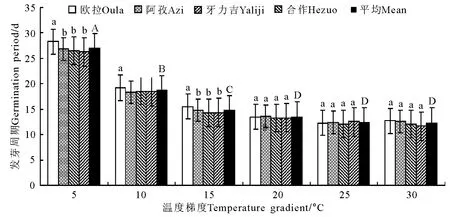
图4 恒温条件下的发芽周期变化Fig.4 Germination periods under constant temperature conditions
就4个采自不同海拔高度的草籽而言,在5℃的低温条件下,欧拉、阿孜和牙力吉3地的种子发芽率差异较小(P>0.05),但均显著高于合作地区(P<0.05),在10℃条件下亦表现出类似的特征,但各产地间的差异不显著(P>0.05);在15℃时,四地间发芽率趋于一致,但在20℃以上的3个梯度下情况发生逆转,高海拔产地开始明显低于低海拔产地,尤以30℃时最为明显,合作和牙力吉两地的种子发芽率显著高于(P<0.05)海拔相对较高的欧拉和阿孜。发芽势的情况与发芽率有所不同,在5和10℃的低温条件下,发芽势表现为随海拔高度下降而上升的趋势,欧拉和阿孜显著低于(P<0.05)牙力吉和合作,而欧拉与阿孜、牙力吉与合作之间无显著差异(P>0.05);15和20℃条件下,各海拔高度间均无显著差异(P>0.05),而温度上升至25℃高海拔地与低海拔地间的差异又重新显现,且仍以高海拔地显著小于(P<0.05)低海拔地为特征,当温度升至30℃时,虽也表现为与25℃时的相同趋势,但其差异程度则明显减小(P>0.05)。活力指数只有在5℃时表现出了高海拔地高于低海拔地的明显差异,而10℃至25℃的4个梯度水平则均表现为低海拔明显高于高海拔的变化,30℃时,不同海拔间的差异基本消失;在小于15℃的3个温度梯度下,发芽周期均表现为随海拔高度的上升而延长的基本态势,但只有欧拉与其他3地存在显著差异(P<0.05),而其他3地间差异均不显著(P>0.05);20℃至30℃的3个梯度,发芽周期在4个海拔高度间均无显著性差异(P>0.05)。
2.2 变温处理 在6个变温处理中,发芽率在-5℃/5℃条件下最低,其次为20℃/30℃,二者均显著低于其他4个处理(P<0.05),且二者间亦存在显著差异(P<0.05);0 ℃/10 ℃、5 ℃/15 ℃、10℃/20℃和20℃/30℃的4个处理间虽以10℃/20℃略高于其他3个处理,但差异均不显著(P>0.05)(图5)。发芽势总的变化态势与发芽率基本相同(图6)。活力指数在-5℃/5℃、0℃/10℃、5℃/15℃、10℃/20℃和15℃/25℃的4个处理中,基本呈随温度的上升而显著增加的趋势,除10℃/20℃、15℃/25℃和20℃/30℃三者之间差异不显著外(P>0.05),其他相邻各处理间均存在显著差异(P<0.05);至20℃/30℃的峰值后,活力指数随温度继续上升而逐渐(15℃/25℃)下降(图7)。发芽周期总体上表现为随温度的上升而下降,发芽周期最长的处理为-5℃/5℃,历时18.8d,最短的处理为20℃/30℃,历时12.2d,二者相差6.6d,存在显著差异(P<0.01)。温度相对较低的3个处理,发芽周期差异较为明显(P<0.05),且均显著高于温度较高的3个处理(P<0.05),而温度较高的3个处理间,发芽周期则无显著差异(P>0.05)(图8)。不同海拔高度的草籽,发芽率、发芽势在各温度梯度下差异不显著(P>0.05);在2个低温梯度下,种子活力有随海拔升高而上升的微弱趋势(P>0.05),但在5℃/15℃、10℃/20℃、15℃/25℃和20℃/30℃的4个梯度上却普遍表现为相反的结果,即随海拔的上升而明显下降的趋势(P<0.05);发芽周期总体上呈虽海拔上升而增加的趋势,但只有在低温的3个梯度下欧拉和阿孜显著高于牙力吉和合作(P<0.05),而在温度相对较高的3个温度梯度下,各海拔高度间差异不显著(P>0.05)。
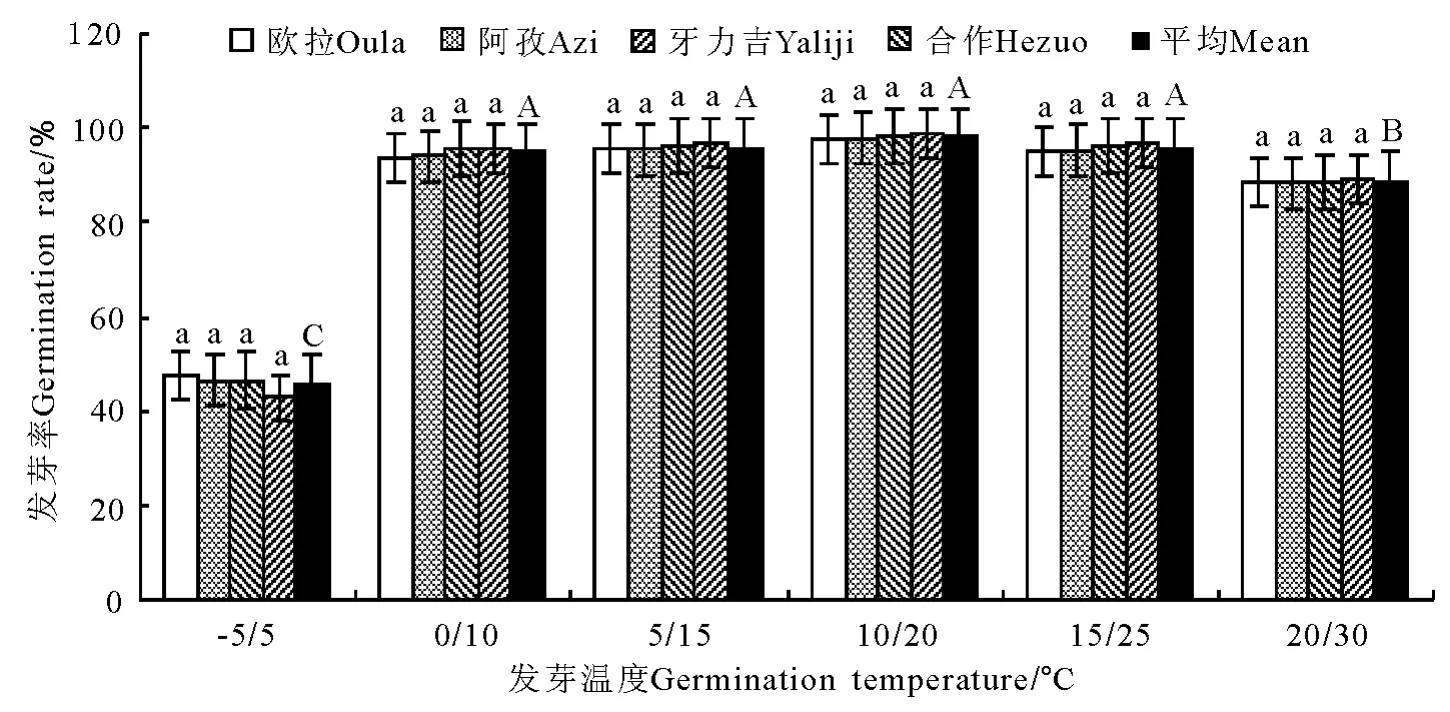
图5 变温条件下发芽率的变化Fig.5 Germination rate under variable temperature conditions
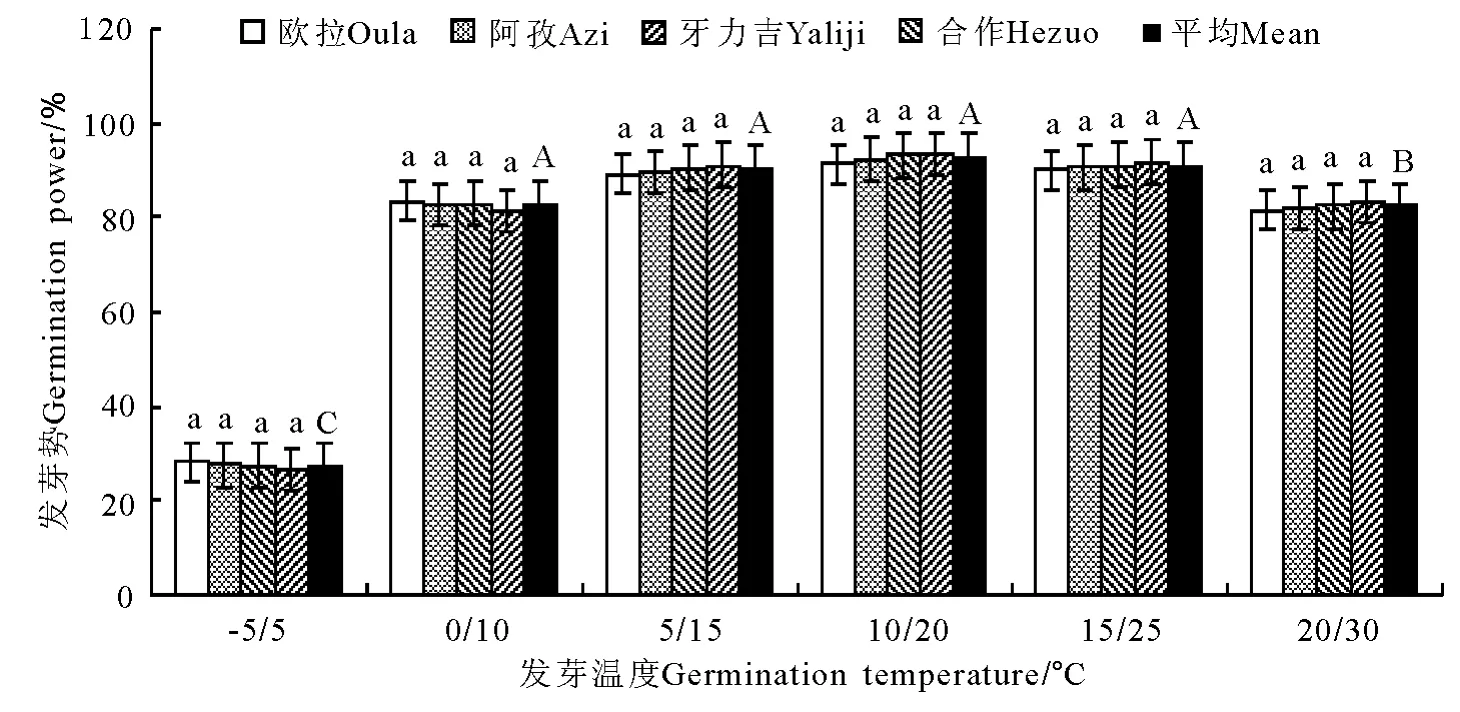
图6 变温条件下发芽势的变化Fig.6 Germination power under variable temperature conditions
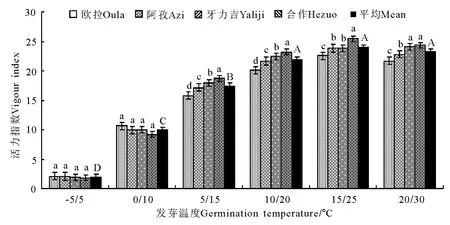
图7 变温条件下的活力指数的变化Fig.7 Vigour indexes under variable temperature conditions
2.3 光照处理 无论是发芽率、发芽势,还是活力指数和发芽周期,在光照处理条件下的表现均与变温处理条件下极其相似,略有不同的是,在变温处理下,种子的活力指数在中高温区段有随处理温度上升而加强的明显趋势,而在添加了光照处理之后,这种差异有所减小(图9-12)。这一结果总体上表明,光照对垂穗披碱草种子萌发的影响微乎其微。
2.4 综合比较 在不考虑温度梯度和不同采集地因素,仅以不同温度处理方式为比较基准的条件下,垂穗披碱草种子发芽情况如图13、14所示。
发芽率在恒温条件下均显著低于(P<0.05)变温和光照处理,而变温和光照处理间差异不显著(P>0.05)(图13)。种子活力在变温处理下虽明显高于恒温处理,但差异不显著(P>0.05),而光照处理则显著高于(P<0.05)恒温处理,变温与光照处理间差异不显著(图14)。发芽周期在恒温处理下平均为16.4d,显著高于(P<0.05)变温(14.6 d)和光照(14.0d)处理,而变温与光照处理间差异不显著(P>0.05)。
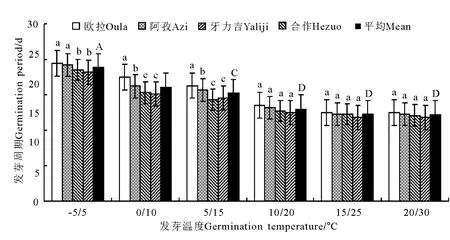
图8 变温条件下的发芽周期变化Fig.8 Germination periods under variable temperature conditions
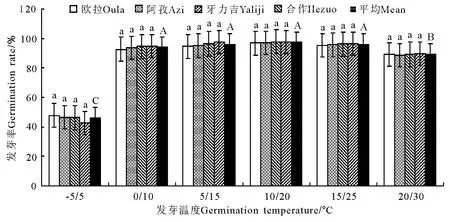
图9 光照条件下的发芽率变化Fig.9 Germination rate under illumination conditions
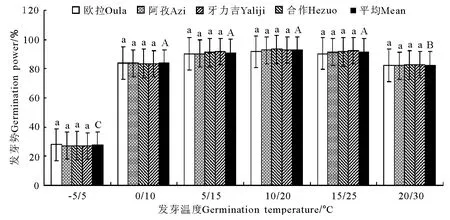
图10 光照条件下的发芽势变化Fig.10 Germination power under illumination conditions
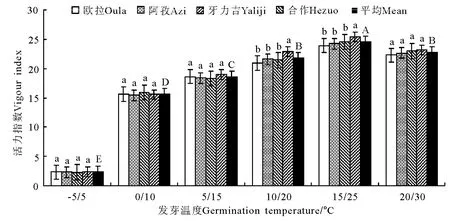
图11 光照条件下的活力指数的变化Fig.11 Vigour indexes under illumination conditions
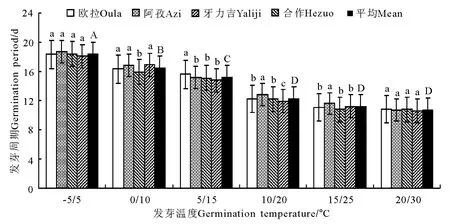
图12 光照条件下的发芽周期变化Fig.12 Germination periods under illumination conditions
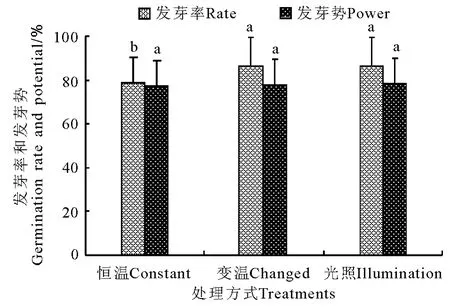
图13 不同处理条件下发芽率和发芽势的比较Fig.13 Comparison of germination rate and power under different treatments
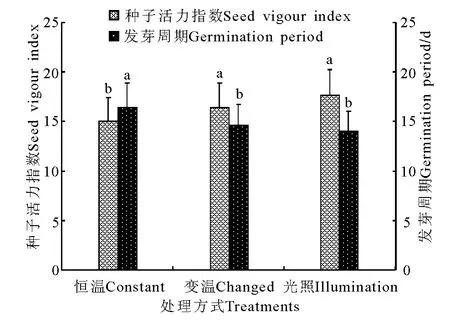
图14 不同处理条件下活力指数和发芽周期的比较Fig.14 Comparison of seed vigour indexes and germination periods under different treatments
3 讨论
垂穗披碱草作为高寒地区的本土草种,近年来在栽培草地建植和天然草地恢复与改良中的应用日趋广泛,用量不断增加,并以其高度的适应性和优良的品质而成为高寒地区的当家草种[6]。然而,垂穗披碱草虽经20余年的栽培驯化,其基本的野生性状并未得到完全去除,因此,在应用实践中,难免因其不适合一般的栽培规范而在具体的技术操作上无法做到精准无误。比如,对于种子用量的确定,最重要的参数便是种子的发芽率,如果不能取得这一参数的准确数值就不能求得精准无误的理论用种量,因而就会影响到对草籽种用价值的估算,进而影响草地的建植质量和成本。对一般栽培草种而言,发芽率的测定均有一定的标准规程[7],但对于垂穗披碱草而言,应用常规的技术规程就很难准确测出其真实的发芽率,因为实验室所设定的条件并不代表其最佳的发芽条件,而野生环境条件可能更有利于其种子萌发[8]。在实践中,常会出现在实验室所测发芽率极低的种子在播种之后其出苗数远高于理论预测值的情况,也正因为如此,种子质量检测的委托方经常会对种子检测部门出具的检测报告提出质疑。这也是证明垂穗披碱草因其野生特性的存在而不应以栽培草种等同对待的实证。本研究的结果也充分证明,垂穗披碱草种子的适宜萌发条件更接近于其野生的自然环境,至少对温度条件的选择表现出了对原产地萌发温度特别是变温环境的高度适应性。
在本研究中,垂穗披碱草在恒温条件下发芽率的适宜温度为10~25℃,其间的4个梯度以20℃最高(82.4%),之后依次为10 ℃(80.9%)、25 ℃(80.9%)和15℃(80.3%),但4个梯度间差异均不显著。此外,即使在5℃的低温条件下,其发芽率亦可达到74.1%的较高水平。发芽势的结果进一步缩小了垂穗披碱草对种子最适发芽温度的选择范围,发芽势的最高值出现在15~20℃,且显著高于其他4个温度梯度。活力指数的峰值则偏向于稍高的温度条件,最高值分别出现在20和25℃,但仍以20℃时最高。从发芽周期来看,整体上随温度的上升而缩短,但15~30℃之间的4个梯度间差异却不显著(P>0.05)。这一结果说明,垂穗披碱草种子对发芽温度的要求相对较低,其上限为25℃,最适温度更接近于20℃左右,且生态变幅较大。在变温和光照处理中,发芽率和发芽势的最高值均出现在10℃/20℃的温度梯度上,尽管种子活力和发芽周期同样有倾向于较高温度的明显趋势,但总体上仍无法动摇垂穗披碱草种子的适宜发芽温度明显低于一般栽培草种的基本结论。
许多研究证明,对于野生植物而言,变温条件具有刺激种子萌发、提高种子发芽率的显著作用[9-10],此外,对于部分草种而言,一定的光照条件也具有类似的明显功效[11-12]。对此,学者们普遍认为,野生草种的种子具有较栽培草种更强的休眠特性,这是对野生环境的一种适应性,具有重要的生态学意义[13]。在本研究中,变温处理除-5℃/5℃梯度水平外,其余5个梯度水平的发芽率和发芽势均显著高于(P<0.05)其所对应的恒温处理,如果在统计中同时剔除变温处理-5℃/5℃及其对应的恒温处理5℃的数据,5个变温处理水平下的平均发芽率和发芽势分别可达94.6%和87.8%,而恒温处理则分别为79.5%和73.3%,差异均极显著(P<0.01)。这说明,垂穗披碱草种子的野生性状仍较为突出,在恒温条件下所获取的种子发芽率与其实际发芽率存在着很大的差异。然而,在本研究中,无论是发芽率、发芽势和种子活力,还是发芽周期,在昼夜交替式光照和昼夜全光照两种处理间差异均不显著(P>0.05),这说明,垂穗披碱草种子的萌发对光照条件反应不敏感,但也有可能与人工光照条件和自然光照条件存在较大差异有关。
生活于不同海拔高度和纬度的同种植物由于生存环境的差异也可造成种子对萌发条件的适应性分异[14-15]。在本研究中,不同海拔高度种子的发芽能力因温度条件不同而表现出较为明显的差异。总体而言,在低温条件下,高海拔种子的综合发芽能力要优于低海拔种子,而在高温条件下则表现为低海拔优于高海拔的趋势,但在中温条件下,各海拔高度间却无明显差异。另外,如果单纯考虑海拔因素,这种差异的产生显然与海拔高度的差异程度密切相关,而且只有当海拔高度的差异达到一定程度时,种子发芽特性的差异才会有较为明显的表现[16]。在本研究中,这种可导致垂穗披碱草种子发芽特性发生明显分异的海拔高度差至少应在700m以上,而这种差异性表现是否来自于遗传基因的支配还有待做进一步研究。
4 结论
1)垂穗披碱草具有较强的野生特性,对种子萌发温度要求较低,在恒温条件下的适宜发芽温度范围为15~25℃。
2)昼夜变温条件可显著提高垂穗披碱草种子的发芽率和整齐度,最佳变温组合为10℃/20℃,发芽率和发芽势分别较20℃恒温条件提高20%。
3)垂穗披碱草种子萌发对光照条件反应不敏感。
4)不同海拔高度的垂穗披碱草种子在相同发芽条件下的发芽性状存在一定差异,但这种差异在不同发芽条件下却有着不同的表现。在高温条件下,低海拔种子的总体发芽能力要优于高海拔种子,而在低温条件下则表现为高海拔优于低海拔的趋势。
[1] 张卫国,江小雷.鼢鼠的造丘活动及不同休牧方式对草地植被生产力的影响[J].西北植物学报,2004,24(10):1882-1887.
[2] 刘蓉,张卫国,江小雷,等.垂穗披碱草群落退化演替的植被特性及其与土壤性状的相关性研究[J].草业科学,2010,27(10):96-103.
[3] 鱼小军,师尚礼,龙瑞军,等.生态条件对种子萌发影响的研究进展[J].草业科学,2006,23(10):44-49.
[4] Jack L B,Gary E L,Volker B.Germination Characteristics of Prairie Dropseed,Blanketflower,and Hairy Goldaster in Response to Prechill and Temperature Treatments[D].South Dakota:South Dakota State U-niversity,2011.
[5] 乔安海,韩建国.垂穗披碱草种子成熟过程中活力变化及适宜收获期研究[J].安徽农业科学,2010,38(22):11847-11850.
[6] 田福平,李锦华,张小甫,等.钾肥对西藏垂穗披碱草种子产量的影响[J].种子,2010,29(12):9-12.
[7] 唐怀建,陈威,王伶俐,等.GB/T 5520-2011粮油检验籽粒发芽试验[S].北京:中国标准出版社,2011.
[8] Raboteaux N N G,Anderson N O.Germination of Cleome hassleriana and Polanisia dodecandra seed lots in response to light,temperature and stratification[J].Research Journal of Seed Science,2010,3(1):1-17.
[9] Qu X X,Huang Z Y,Jerry M,et al.Effect of temperature,light and salinity on seed germination and radicle growth of the geographically widespread halophyte shrub halocnemum strobilaceum[J].Annals of Botany,2008,101:293-299.
[10] Meulebrouck K,Ameloot E,Van Assche J A,et al.Germination ecology of the holoparasite Cuscuta epithymum[J].Seed Science Research,2008,18:25-34.
[11] Nonogaki H.Seed germination-the biochemical and molecular mechanisms[J].Breeding Science,2006,56:93-105.
[12] Verma S K,Kumar b,Ram G,et al.Varietal effect on germination parameter at controlled and uncontrolled temperature in Palmarosa (Cymbopogon martinii)[J].Industrial Crops and Products,2010,32:696-699.
[13] Graae B J,Also I G,Ejrnaes R.The impact of temperature regimes on development,dormancy breaking and germination of dwarf shrub seeds from arctic,alpine and boreal sites[J].Plant Ecology,2008,198(2):275-284.
[14] 苗佳敏,张新全,陈智华,等.青藏高原和新疆地区垂穗披碱草种质的SRAP及RAPD分析[J].草地学报,2011,19(2):306-316.
[15] 马啸,周永红,于海清,等.野生垂穗披碱草种质的醇溶蛋白遗传多样性分析[J].遗传,2006,28(6):699-706.
[16] 邹华.高寒地区垂穗披碱草种子生产草地土壤种子库研究[J].草业与畜牧,2009,161(4):11-13.
猜你喜欢
草业学报(2022年11期)2022-11-23
美文(2022年18期)2022-10-20
现代畜牧科技(2021年7期)2021-07-28
西藏农业科技(2021年4期)2021-04-18
现代园艺(2020年19期)2020-10-02
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
中国盐业(2018年16期)2018-12-23
中国粮油学报(2016年5期)2016-01-23
