
4种病原弧菌外膜蛋白的提取及抗原性初步分析
2012-10-23 03:01阎斌伦张晓君秦国民毕可然
海洋科学 2012年5期
阎斌伦, 张晓君, 秦国民, 毕可然, 徐 静, 秦 蕾
(淮海工学院 江苏省海洋生物技术重点实验室, 江苏 连云港 222005)
4种病原弧菌外膜蛋白的提取及抗原性初步分析
阎斌伦, 张晓君, 秦国民, 毕可然, 徐 静, 秦 蕾
(淮海工学院 江苏省海洋生物技术重点实验室, 江苏 连云港 222005)
利用十二烷基磺酸钠抽提并结合超速离心的方法提取了鳗弧菌(Vibrio anguillarum)、河口弧菌(V.aestuarianus)、霍乱弧菌(V.cholerae)和副溶血弧菌(V.parahaemolyticus)4种致病性弧菌的主要外膜蛋白, 通过SDS-PAGE分析比较4种弧菌主要外膜蛋白的组分。结果表明: 4株弧菌的外膜蛋白电泳图谱一般可得到5~10条蛋白带, 分子量多数集中在26~40 kD和48~85 kD。对所提取的4种弧菌主要外膜蛋白进行SDS-PAGE后, 分别与自制的兔抗鳗弧菌血清、抗河口弧菌血清、抗副溶血弧菌血清进行Western-blotting印迹分析。结果显示每种全菌抗血清可与相应菌的外膜蛋白部分组分发生免疫反应,并与其他3种弧菌外膜蛋白的部分组分产生交叉免疫反应, 这些反应条带分子量主要集中于30~48 kD之间。本研究为进一步研究致病性弧菌外膜蛋白免疫学特性提供参考。
鳗弧菌(Vibrio anguillarum); 河口弧菌(V.aestuarianus); 霍乱弧菌(V.cholerae); 副溶血弧菌(V.parahaemolyticus); 外膜蛋白; Western-blotting印迹
由弧菌属(Vibrio)细菌引起的“弧菌病(Vibriosis)”是在世界各地海、淡水养殖鱼, 虾, 蟹,贝类等水产动物中普遍流行且危害最大的细菌性疾病, 给水产养殖业造成了极大的经济损失。因此, 弧菌病的有效预防与控制已成为目前水产动物的健康养殖和可持续发展的重要保障。外膜蛋白(Outer Membrane Protein, OMP)位于革兰氏阴性菌表面, 产量丰富, 具有良好的免疫原性, 不仅可激发机体的体液免疫, 而且还可引起细胞免疫并可与其他相关病原菌及同种不同血清型菌株产生免疫交叉反应,可认为是一种潜在的共同免疫保护性抗原。因此主要外膜蛋白是病原细菌疫苗的重要材料, 是制备免疫保护特异性强的亚单位疫苗的候选成分, 分离出参与血清学反应的主要外膜蛋白并制备相应免疫制剂, 将能有效保证其相应的免疫保护效果, 近年来的研究也证实了病原细菌外膜蛋白在作为保护性抗原方面的作用。
几年来, 作者在对水产动物细菌性疾病及相应病原细菌的研究中, 分离到具有强致病性的鳗弧菌(V.anguillarum)、副溶血弧菌(V.parahaemolyticus)、霍乱弧菌(V.cholerae)及河口弧菌(V.aestuarianus)[1-4]。本研究以此 4种病原弧菌为研究材料, 提取并比较了4种病原弧菌外膜蛋白成分, 并通过Western-blot初步分析了 4种病原弧菌外膜蛋白的抗原性。本研究旨在探索 4种病原弧菌外膜蛋白组分及抗原性,为致病性弧菌免疫学研究提供参考。
1 材料和方法
1.1 供试菌株
供试鳗弧菌(BH1株)及河口弧菌(TS1株)分离自江苏省赣榆县某工厂化养殖病(死)半滑舌鳎(Cynoglossus semilaevis L.); 副溶血弧菌(JGB080708-1株)分离自江苏省赣榆县池塘养殖发病凡纳滨对虾幼虾(Litopenaeus vannamei L.); 霍乱弧菌(LD081008B-1株)分离自江苏省赣榆县池塘养殖发病泥鳅(Misgurnus anguillicaudatus L.), 均为本实验室分离鉴定并保存。
1.2 免疫血清的制备
1.2.1 菌体抗原的制备
4株供试菌接种于普通营养肉汤培养基, 置 28℃培养18 h后, 加入福尔马林溶液0.5 mL(0.5%)后于37 ℃作用24 h灭活(菌检合格供用), 以6 000 r/min离心15 min后弃上清, 沉淀菌体经无菌生理盐水洗涤3次后, 再用无菌生理盐水16 mL悬浮即为全菌(OK)免疫原。
1.2.2 免疫血清的制备
取上述OK免疫原, 经耳静脉注射接种体质量2 kg左右的健康家兔。接种前取家兔血少许分离血清后, 以上述菌株为抗原, 对所用家兔进行相应的血清玻片凝集试验, 阴性的用于免疫接种。免疫剂量为2.0 mL/次, 间隔6 d免疫1次, 共4次, 末次后的7 d耳静脉采血少许分离血清测定其凝集效价, 合格后心脏采血分离血清, 置-20℃冻结保存供用。
1.3 外膜蛋白的提取
分别将4种弧菌接种于150 mL的营养肉汤培养基中, 28℃过夜培养 18 h后低速离心细菌培养物(4℃、4 000 r/min、30min), 收集菌体; 用20 mmol/L Tris-HCl (pH 7.4, 含10 mmol/L EDTA) 缓冲液洗涤3次后, 溶于适量缓冲液中, 置冰浴中超声波破碎细胞 10 min(振幅50%); 以4℃、6 000 r/min离心30 min,收集上清液; 经4℃、15 000 r/min离心1 h, 弃上清;沉淀溶于含0.5%(w/v)SDS的上述缓冲液中, 置37℃作用30 min后, 以4℃、15 000 r/min离心1 h, 弃上清; 然后用20 mmol/L Tris-HCl缓冲液洗涤沉淀1~2次, 溶于双蒸水, 置-20℃保存备用。
1.4 外膜蛋白SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)
采用解离非连续缓冲系统垂直板电泳。将所提取的外膜蛋白与电泳上样缓冲液(0.5 mol/L Tris-HCl;pH=6.8; 10% SDS; 1% β-巯基乙醇; 10%甘油; 0.05%溴酚蓝)按照4:1体积混匀, 加热煮沸5 min, 冷却后低速离心 1 min, 用微量加样器加样, 每孔加入 15 μL样品。采用 Tris-甘氨酸(Gly)电泳缓冲液(0.025 mol/L Tris-Base; 0.25 mol/L甘氨酸; 0.1% SDS; pH 8.3), 4℃条件下电泳。丙烯酰胺浓度: 分离胶10%(v/v), 浓缩胶 4.75%(v/v)。浓缩胶部分恒定电流为40 mA, 分离胶部分恒定电流为60 mA。电泳结束后, 取出凝胶经考马斯亮蓝R250(Sigma)染色后用全自动凝胶成像系统扫描, Gel-Pro Analyzer软件分析各蛋白的分子量。
1.5 Western-blotting印迹试验
SDS-PAGE结束后, 利用半干法对膜蛋白进行转印, 即将凝胶均匀转移至硝酸纤维素(NC)膜上, 40 mA转移1 h。含有5%牛血清白蛋白(BSA)的PBS缓冲液4 ℃封闭过夜, PBST (含有0.05 % Tween20的PBS) 洗涤 3次, 每次5 min。然后分别将不同的膜浸在稀释50倍的兔抗鳗弧菌、河口弧菌、副溶血弧菌全菌血清中37 ℃温育1 h。PBST洗膜3次, 每次5 min。于碱性磷酸酶(AP)标记的羊抗兔IgG(1:1000,Sigma)中37 ℃温育1 h。PBST洗膜3次, 每次5 min。最后将膜用含有66 μL NBT贮液(0.5 g NBT溶于10 mL70%的二甲基亚砜中)和 33 μL BCIP贮液(0.5 g BCIP溶于10 mL 100%二甲基亚砜中)的NBT/BCIP 缓冲液(10 mL)中发色 15~20 min, 双蒸水水洗终止反应、晾干。
2 结果
2.1 SDS-PAGE图谱
鳗弧菌、河口弧菌、副溶血弧菌和霍乱弧菌由SDS法制备的外膜蛋白经SDS-PAGE电泳显示如图1。副溶血弧菌主要有 5条蛋白带, 分子量大约在28~65 kD, 其中分子量大约在 29 kD的条带产物量较多; 鳗弧菌主要有 5条蛋白带, 分子量在 40~26 kD, 其中分子量大约在 28 kD的条带产物量较多;河口弧菌主要有 8条蛋白带, 分子量大约在 24~65 kD, 其中分子量大约在24 kD和26 kD的条带产物量较多; 霍乱弧菌主要有 10条蛋白带, 分子量大约在19~60 kD, 其中分子量大约在30 kD的条带产物量较多。电泳结果显示, 鳗弧菌、河口弧菌、副溶血弧菌和霍乱弧菌外膜蛋白组分不同, 但每种弧菌都具有一条产物量较多的条带, 鳗弧菌大约在 28 kD,河口弧菌大约在 26 kD, 副溶血弧菌大约在 30 kD,霍乱弧菌大约在32 kD。
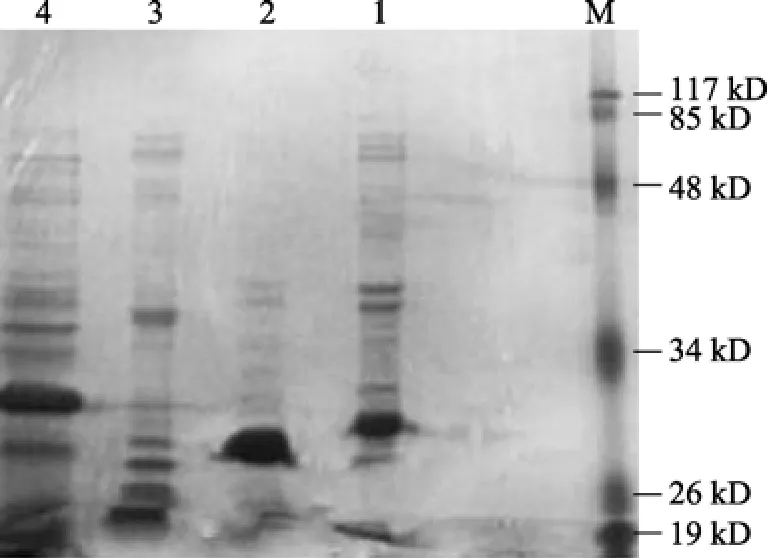
图1 4种病原弧菌外膜蛋白的SDS-PAGE图谱Fig. 1 SDS-PAGE profiles of outer membrane proteins of 4 pathogenic Vibrio sp.1. 副溶血弧菌; 2. 鳗弧菌; 3. 河口弧菌; 4. 霍乱弧菌; M.Marker(下同)1. V.anguillarum; 2. V.aestuarianus; 3. V.cholerae; 4. V.parahaemolyticus; M.Marker
2.2 Western-blotting结果
兔抗鳗弧菌、抗副溶血弧菌、抗河口弧菌血清与各外膜蛋白的 Western-blotting印迹表明, 兔抗鳗弧菌血清与霍乱弧菌和鳗弧菌菌株自身有1条免疫反应带,与副溶血弧菌有 1条免疫反应带, 而与河口弧菌没有出现免疫反应带(图 2); 兔抗河口弧菌血清与鳗弧菌出现3条免疫反应条带, 与河口弧菌自身菌株出现2条免疫反应条带, 与霍乱弧菌出现1条免疫反应条带, 与副溶血弧菌则无免疫反应条带出现(图3); 兔抗副溶血弧菌血清与鳗弧菌菌株出现 5条免疫反应条带, 与河口弧菌出现 4条主要的免疫反应带, 与副溶血弧菌自身菌株出现1条免疫反应条带(图4)。

图2 兔抗鳗弧菌血清作为一抗的免疫图谱Fig. 2 Western-blot profile with rabbit serum against V.anguillarum as the first antibody
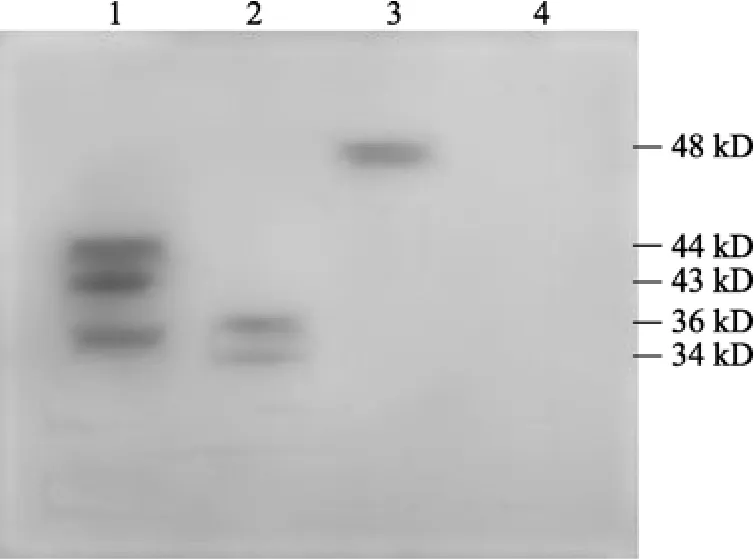
图3 兔抗河口弧菌血清作为一抗的免疫图谱Fig. 3 Western-blot profile with rabbit serum against V.aestuarianus as the first antibody

图4 兔抗副溶血弧菌血清作为一抗的免疫图谱Fig. 4 Western-blot profile with rabbit serum against V.parahaemolyticus as the first antibody
3 讨论
国内外在鱼类病原弧菌的外膜蛋白的免疫性状研究方面已多有报道, 吴灶和等[5]报道溶藻弧菌外膜蛋白对凡纳滨对虾具有较好的免疫原性; 黄志坚等[6]及段翠兰等[7]报道溶藻酸弧菌的主要外膜蛋白的免疫原性很强, 是重要的保护性抗原; 毛芝娟等[8]从浙江省象山港网箱养殖大黄鱼分离的一株副溶血弧菌(V.parahaemolyticus)中克隆了两种铁调外膜蛋白psuA和pvuA的基因, 获得大量表达后免疫大黄鱼, 结果显示该两种铁调外膜蛋白具有良好的免疫原性, 有可能作为高效疫苗成分。张崇文等[9]从哈维氏弧菌的总 DNA 中成功克隆了外膜蛋白 OmpK基因并在大肠杆菌中实现了高效表达, 用纯化的融合重组蛋白免疫新西兰白兔获得了高效价的抗血清,免疫印迹显示抗血清和哈氏弧菌外膜蛋白中分子质量约为 27 kD 的蛋白产生了一条特异反应带, 提示哈氏弧菌外膜蛋白 OmpK是它的保护性抗原之一,有望作为其疫苗的有效成分在大黄鱼哈氏弧菌病的防治中发挥作用。周丽等[10]分离鳗弧菌和溶藻胶弧菌(V.alginolyticus)的外膜蛋白并通过免疫印迹对其特性进行分析, 发现分子量大小为 51 kD的外膜蛋白可能是鳗弧菌和溶藻胶弧菌共有的抗原; Chart等[11]从鳗弧菌分离的 2个小分子量的外膜蛋白能有效的刺激机体的免疫反应, 说明外膜蛋白也是鳗弧菌很好的保护性抗原; 董传甫等[12]发现分子量为36 kD的主要外膜蛋白是副溶血弧菌和溶藻弧菌具有强免疫性的主要抗原。
外膜蛋白是革兰氏阴性菌细胞壁的特有成分, 位于细菌的表层, 在维持外膜结构, 物质转运方面发挥着重要作用, 具有直接与机体免疫系统接触的机会。本研究表明鳗弧菌、河口弧菌、副溶血弧菌和霍乱弧菌具有不同的外膜蛋白组分, 但每种弧菌都具有一条产物量较多的条带, 有望作为同种不同血清型菌株间的共同免疫保护抗原; Western-blotting印迹分析表明, 每种供试菌的全菌抗血清可与自身的外膜蛋白部分组分发生免疫反应, 并与其他 3种弧菌外膜蛋白的部分组分产生交叉免疫反应。该研究结果可为弧菌免疫学深入研究提供参考。
[1] 张晓君, 秦国民, 阎斌伦, 等. 半滑舌鳎病原鳗利斯顿氏菌表型及分子特征研究[J]. 海洋学报, 2009, 31(5):112-122.
[2] 张晓君, 陈翠珍, 阎斌伦, 等. 凡纳滨对虾病原副溶血弧菌的表型及分子特征[J]. 海洋与湖沼, 2009, 40(5):654-661.
[3] 邴旭文, 阎斌伦, 张晓君, 等. 泥鳅病原霍乱弧菌的表型与分子鉴定[J]. 海洋与湖沼, 2009, 40(6): 692-698.
[4] Zhang X J, Qin G M, Bing X W, et al. Molecular and phenotypic characterization of Vibrio aestuarianus, a pathogen of the cultured tongue sole Cynoglossus semilaevis[J]. Journal of Fish Diseases, 2011, 34, 57-64.
[5] 吴灶和, 曹剑香, 简纪常, 等. 溶藻弧菌外膜蛋白对凡纳滨对虾免疫功能的影响[J]. 热带海洋学报, 2005,24(6): 1-5.
[6] 黄志坚, 何建国. 溶藻弧菌外膜蛋白(Va-OMP)的免疫原性及免疫保护性[J]. 水产学报, 2006, 30(4): 538-543.
[7] 段翠兰, 吴灶和, 陈辉, 等. 溶藻弧菌 0l株外膜蛋白的分离及其抗原性分析[J]. 水产学报, 2004,28(B12): 112-116.
[8] 毛芝娟, 由振强, 魏永伟, 等. 副溶血弧菌ZJ2003株两种铁调外膜蛋白的克隆、表达和免疫原性[J]. 中国水产科学, 2007, 14(4): 563-569.
[9] 张崇文, 于涟, 毛芝娟, 等. 哈维氏弧菌外膜蛋白OmpK基因的克隆及原核表达[J]. 水产学报, 2006,30(1): 9-14.
[10] 周丽, 刘洪明, 战文斌, 等. 鳗弧菌、溶藻胶弧菌外膜蛋白的分离及特性[J]. 中国水产科学, 2003, 10(1): 31-35.
[11] Chart H, Trust T J. Characterization of the surface antigens of the marine fish Pathogens, Vibrio anguillarum and Vibrio ordalii [J]. Canadian Journal of Microbiology, 1984,30: 703-710.
[12] 董传甫, 林天龙, 许斌福, 等. 电泳和免疫印迹分析副溶血弧菌和溶藻弧菌主要外膜蛋白和多糖抗原[J].中国人兽共患病杂志, 2004, 20(7): 619-623.
Isolation and antigenicity analysis of outer membrane proteins from 4 pathogenic Vibrio sp.
YAN Bin-lun, ZHANG Xiao-jun, QIN Guo-min, BI Ke-ran, XU Jing, QIN Lei
(Jiangsu Key Laboratory of Marine Biotechnology, Huaihai Institute of Technology, Lianyungang 222005, China)
Jul., 4, 2011
Vibrio anguillarum; Vibrio aestuarianus; Vibrio cholerae; Vibrio parahaemolyticus; outer membrane protein;Western-blotting
The outer membrane proteins of 4 pathogenic Vibrio sp. (V.anguillarum, V.aestuarianus, V.cholerae and V.parahaemolyticus) were extracted using sodium laurylsulfonate and ultracentrifuging methods, and the ingredient of the outer membrane proteins were analyzed using SDS-PAGE. The SDS-PAGE results showed that five to ten proteins bands could be obtained for each vibrio with molecular weights ranging from 26 to 40 kD and from 48 to 85 kD. Western-blotting analysis was conducted after SDS-PAGE, the results showed that some of the outer membrane protein bands have cell antigenicity in the Western-blotting with rabbit antiserum against whole cell of V.anguillarum, V.aestuarianus,V.cholerae and V.parahaemolyticus, respectively, the molecular weights were between 30 to 48 kD. This study can provide reference for immunologic research of outer membrane proteins of pathogenic Vibrios.
S94
A
1000-3096(2012)05-0071-04
2011-07-04;
2011-09-01
江苏省六大人才高峰项目(2009); 连云港市科技攻关资助项目(CG0907); 江苏省水产三项工程资助项目(P2009-18)
阎斌伦(1962-)男, 江苏赣榆人, 教授, 主要从事应用海洋生物技术与水产健康养殖研究, E-mail:yanbinlun@yahoo.com.cn
(本文编辑:谭雪静)
猜你喜欢
生物工程学报(2017年9期)2017-11-03
西南农业学报(2017年5期)2017-06-23
哈尔滨医药(2016年3期)2016-12-01
环境污染与防治(2016年9期)2016-03-13
微生物学杂志(2015年6期)2015-12-27
癌变·畸变·突变(2015年4期)2015-02-27
现代检验医学杂志(2015年2期)2015-02-06
现代检验医学杂志(2015年1期)2015-02-06
应用海洋学学报(2014年1期)2014-11-22
微生物学杂志(2013年5期)2013-10-25
