
三峡水库香溪河库湾夏季蓝藻水华成因研究
2012-12-25 02:19田泽斌刘德富杨正健方小凤姚绪姣方丽娟三峡大学水利与环境学院三峡库区生态环境教育部工程研究中心湖北宜昌443002湖北工业大学资源与环境学院湖北武汉430072武汉大学水利水电学院湖北武汉430072
中国环境科学 2012年11期
田泽斌,刘德富,2*,杨正健,3,方小凤,姚绪姣,方丽娟 (.三峡大学水利与环境学院,三峡库区生态环境教育部工程研究中心,湖北 宜昌 443002;2.湖北工业大学资源与环境学院,湖北 武汉 430072;3.武汉大学水利水电学院,湖北 武汉 430072)
三峡水库香溪河库湾夏季蓝藻水华成因研究
田泽斌1,刘德富1,2*,杨正健1,3,方小凤1,姚绪姣1,方丽娟1(1.三峡大学水利与环境学院,三峡库区生态环境教育部工程研究中心,湖北 宜昌 443002;2.湖北工业大学资源与环境学院,湖北 武汉 430072;3.武汉大学水利水电学院,湖北 武汉 430072)
为探讨香溪河夏季蓝藻水华发生过程及主要影响因素,于2010年7月19日~8月30日在蓝藻水华暴发区域开展持续监测,并对水华过程进行分析.结果表明,水华持续 25d,自 7月 23日暴发,8月 16日逐渐消退,藻密度最高达到 108.03×106cells/L,优势藻种为鱼腥藻(Anabaena);自身悬浮机制、固氮机制、能够产生藻毒素抑制其他藻类生长,是鱼腥藻成为优势藻种的主要原因;充足的营养物质、显著的水体分层是水华暴发的必要条件,水华期间蓝藻对硝氮利用显著;在具备充足的营养盐的稳定水体中,水温持续升高、混合层与真光层比值的降低是诱发蓝藻水华的关键因子,并且在混合层与真光层比值为0.5时对蓝藻增殖影响最大.
三峡水库;香溪河库湾;蓝藻水华;混合层
三峡水库成库后,藻类水华发生的频率和规模开始增加,优势藻种从河流型向湖泊型转变[1],尤其是2008年开始部分支流库湾夏季暴发的蓝藻水华不仅破坏了生态环境,而且威胁着人类健康及水源安全.目前,有关三峡水库支流有害蓝藻水华频繁出现现象的研究较多[1-3],主要依托野外观测,研究水华期间藻类对特定环境因子的响应关系,以及种群结构和演替特征等.李哲等[4]认为,温度升高、真光层深度下降是诱发夏季蓝藻水华的重要因子,但没能建立复杂物理背景下蓝藻水华与水温的具体响应机制.张敏等[2]指出,库湾相对稳定的水体环境是蓝藻水华暴发的必要条件,但没有给出香溪河库湾具体的水体分层状态.目前对三峡水库支流蓝藻水华的研究成果尚难以应用于阐释深水河道型水库蓝藻水华形成机理.因此,本文以三峡水库典型支流香溪河为例,对2010年7~8月香溪河库湾水华过程进行持续监测,以该区域特殊水体分层背景下蓝藻水华的生态响应机制为研究重点,围绕水温、水体稳定系数、特殊的水体层化模式、营养盐等因子对蓝藻水华的影响机理展开探讨,探究蓝藻形成优势并暴发水华的诱导因子,以期为探讨三峡水库支流水华的机理提供参考,为有效防治支流水华提供理论依据.
1 研究区域及研究方法
1.1 研究区域及样点设置
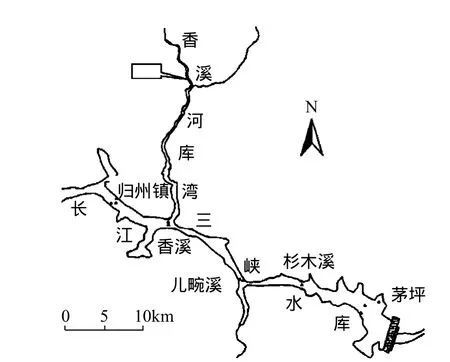
图1 香溪河采样点位置示意Fig.1 Sampling site in Xiangxi Bay
香溪河系长江三峡库区湖北省境内最大的支流,发源于神农架林区,河流由北向南,在秭归香溪汇入长江,其干流长度 94km,流域范围(110°25′~111°06′)E,(30°57′~31°34′)N,流域面积3099km2,拥有南阳河、古夫河、高岚河3条主要支流.地处亚热带大陆性季风气候区,由于地势高差大,地形复杂,气温垂直变化明显.年平均气温为16.6℃,流域多年平均流量为40.18m3/s,年降雨量 1015.6mm.香溪河库湾属深水型回水库湾,由于干支流的水温差异及颗粒物浓度含量不同导致分层异重流现象长期存在,同时库湾存在显著的水温分层现象[5].两种密度相差不大、可以相混的流体,因为密度的差异而发生的相对运动称为异重流[6].三峡水库调度时,长江干流水体与香溪河库湾水体通过异重流方式进行水体交换.依托三峡大学水生态与环境野外观测站,2010年7月-8月在香溪河库湾峡口附近选取香溪河水华暴发严重的区域设置监测点(图1)进行监测.
1.2 监测频率与指标
监测时间为2010年7~8月,每天上午10:00监测1次,主要监测指标有:叶绿素a浓度、藻种及其密度、水温、透明度、流速、水下光强、营养盐(N-N、PO43--P、SiO2-Si)等.在采样点每次取350mL表层水,用GF/C滤膜过滤并10℃保存带回实验室分析,用90%的丙酮提取 24h,3500r/min离心 10min,收集上清液,用分光光度法[7]测定表层叶绿素a浓度.根据《水和废水监测分析方法》[7]测定水中 NO3--N、PO43--P浓度,用硅钼黄分光光度法[7]测定溶解性硅酸盐浓度SiO2-Si;水温由Hydrolab DS5多参仪现场测定;流速采用Vector三维点式流速仪(挪威)测定;现场同步采用手持式风速风向仪现场记录观测点风速风向;用PVC塑料瓶取1200mL水样,加鲁哥固定,带回实验室沉降浓缩成75 mL,在显微镜(10×40倍)下对浮游植物细胞进行计数,藻类根据《中国淡水藻类》[8]和《淡水微型生物图谱》[9]进行鉴定.
1.3 数据处理
本文忽略水体中泥沙对水体密度的影响,水温对应的水体密度按Lawson等[10]的公式(1)计算:
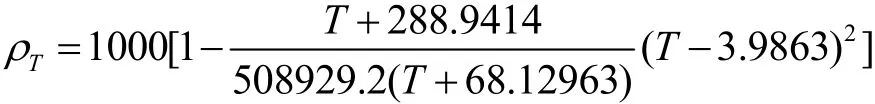
式中: ρT为水体密度,单位 kg/m3;T为水体温度,单位℃.
水体稳定系数(RWCS)是一个无量纲的参数,该参数通过比较底部(Db)和表面(Ds)水的密度差异和纯水在 4℃(D4)和 5℃(D5)时的密度差异,根据公式(2)计算[11]:
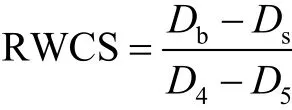
自然条件下的水体真光层深度的计算采用透明度的 2.7倍[12],混合层深度采用与表层水温相差不大于0.2℃的深度范围[13].
2 结果与分析
2.1 叶绿素a浓度及藻密度
研究表明[13],香溪河库湾蓝藻水华暴发时叶绿素a浓度的阀值为40mg/m3.由图2可知,监测期间,香溪河库湾叶绿素a浓度先增大后减小,变幅为0.47~87.48mg/m3.香溪河库湾在7月23日~8月16日暴发了一次蓝藻水华,持续25d.7月19日叶绿素a浓度为31.70mg/m3,随后叶绿素a浓度波动明显,总体呈增大趋势,至8月5日达峰值87.48mg/m3,之后叶绿素 a浓度维持在较高水平波动,8月17日回落至7.23mg/m3,水华消退.此后叶绿素 a浓度出现短期小幅升降过程,并未形成蓝藻水华,8月 25日以后叶绿素 a浓度降至2mg/m3以下.
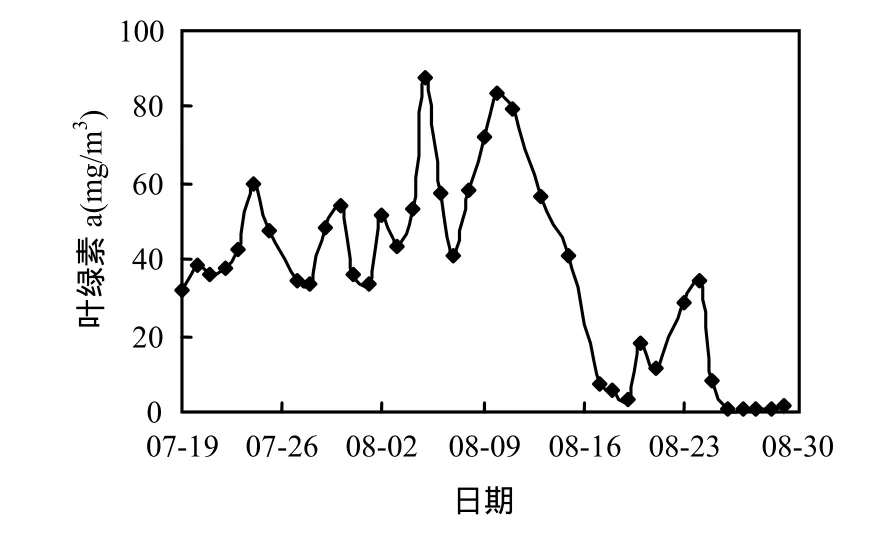
图2 叶绿素a浓度随时间的变化Fig.2 Variation of chlorophyll a concentration in Xiangxi Bay during the summer
藻细胞密度能很直观的表明水体中藻类的多少和水华暴发强度[13],富营养化水体发生水华期间,藻类大量繁殖,藻密度不断增加,当水体发生重度水华时,浮游植物的细胞密度一般大于20×106cells/L[13-14].从图3可知,香溪河库湾藻类总密度整体先增大后减小,与叶绿素 a浓度变化趋势大体一致,镜检得出优势藻种为鱼腥藻(Anabaena).7月19日藻密度为29.46×106cells/L,之后呈上升趋势,7月 28日达峰值 138.54×106cells/L后,呈现先下降后上升的过程,8月5日藻密度增至127.84×106cells/L,此后逐渐回落,8月25日降至0.48×106cells/L后维持较低水平.监测期间,蓝藻密度占总密度的百分比最高达94%,与总密度变化趋势基本一致.
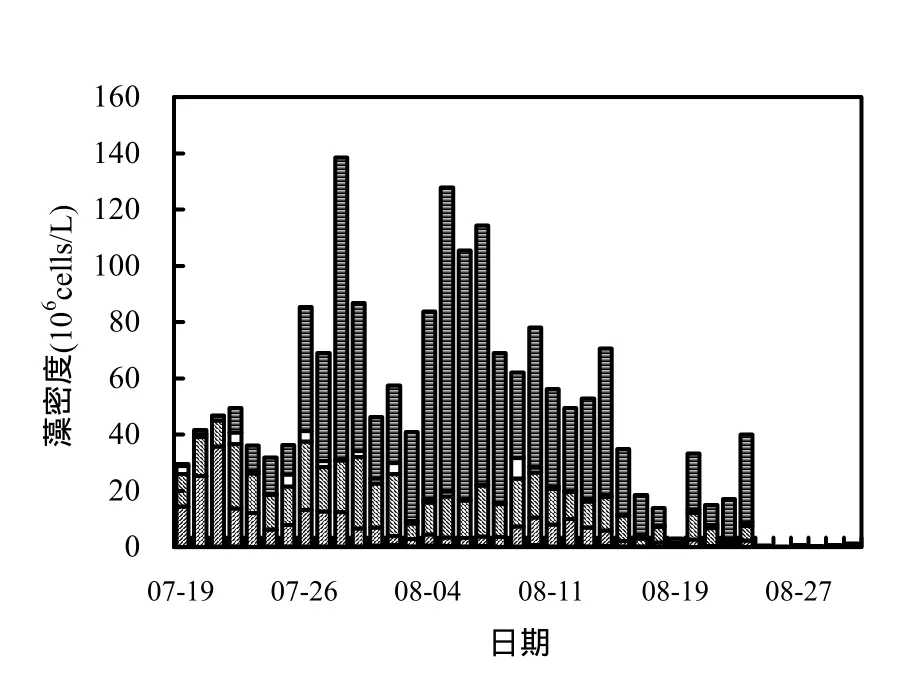
图3 藻密度及藻种组成随时间的变化Fig.3 Variation of cell density and composition in Xiangxi Bay during the summer
2.2 水温

图4 表层水温随时间的变化Fig.4 Variation of surface temperature in Xiangxi Bay during the summer
水温是藻类生长的重要影响因子.它通过控制光合作用、酶促反应或呼吸作用强度,直接影响藻类细胞生长增殖相应的一系列过程[4,15],同时适宜的水温是蓝藻暴发的必要条件[16].由图 4知,香溪河库湾表层水温先增大后减小,在26.31~31.11℃之间,符合多数蓝藻生长增殖最适温度(25~35℃[17]).监测期间,由于受气温降低及降雨的影响,7月19日库湾表层水温较低,随着太阳辐射及气温的显著升高,表层水温升温较快,7月28日达峰值31.11℃,此后在 30℃上下波动,维持蓝藻生长所需的较高温度,8月16日后气温逐渐降低,降雨较为频繁,加之来流量明显增大,加快了水体交换,表层水温呈降低的趋势,至 8月 25日表层水温降至27.66℃,此后呈变幅较小的波动趋势.
2.3 水体稳定系数
水体稳定系数是评价水体垂向混合程度及水体分层状态的重要参数,水体稳定系数越大,水体垂向越稳定,分层现象越显著;水体稳定系数越小,越易形成混合水体[18].水体的稳定性是影响蓝藻生长及分布的重要物理因素,垂直稳定分层越明显的水体,越易暴发高强度的藻类水华[19].
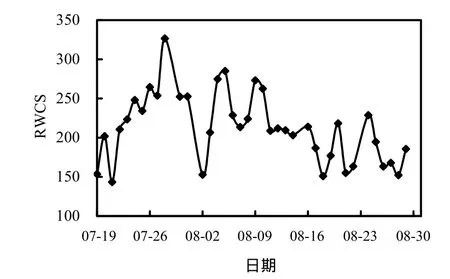
图5 水体稳定系数随时间的变化Fig.5 Variation of RWCS in Xiangxi Bay during the summer
由图5知,监测期间香溪河库湾水体稳定系数先增大后减小,在 143.40~326.66范围变化,与表层水温(图4)变化趋势基本一致.由于表层水温显著升高,表底温差增大,水温分层现象明显,之后随着气温的降低,加之降雨频繁,水体垂向掺混剧烈,表底温差逐渐减小,分层特性减弱,从而使水体稳定系数降低,峰值出现在 7月 28日,达326.66.监测期间水体稳定系数较高,热分层现象明显,蓝藻可以通过控制伪空胞的数量来调控细胞密度,利用其悬浮机制在获取更多的营养和光照竞争中取得优势[20],而水体的分层加速了蓝藻这种喜好在稳定水体中生长的种类大量的繁殖和聚集,致使蓝藻在水体中更具有优势[21].
2.4 真光层、混合层及其比值
真光层是浮游植物接受有效光照进行光合作用的最大深度[22],它直接影响水柱中藻类的生长增殖过程.由图6知,香溪河库湾真光层深度变幅较小,变化范围在 0.8~3.4m之间.混合层是藻类可以迁移悬浮的水层范围,香溪河库湾混合层深度呈先降低后升高趋势,在 0.8~6m范围内变化.8月中下旬开始,混合层深度显著增加,表层水体呈混合状态.在研究中,可以采用混合层和真光层的比值(Zmix/Zeu)来反映藻类在真光层内生长的潜势[23].由图 6知,监测期间 Zmix/Zeu与藻密度变化(图3)趋势相反,在0.3~3.1之间变化,7月19日之后Zmix/Zeu明显减低,随着Zmix/Zeu的降低,真光层范围内的藻类能够利用充足的光照,提高增长率,表现为藻密度相应增加,8月16以前Zmix/Zeu随时间变幅较小,相应藻密度维持较高水平,此后Zmix/Zeu总体呈增加趋势,混合层深度明显大于真光层深度,表现为混合层内的藻类受到光照的限制,生长率降低,从而对藻类增殖起到抑制作用.

图6 真光层、混合层深度及其比值随时间变化Fig.6 Variations of Zeu, Zmix and Zmix/Zeu in Xiangxi Bay during the summer
2.5 营养盐
溶解性营养盐可以被浮游植物直接吸收利用进行自身生长增殖.香溪河库湾氮磷营养含量丰富[24],对蓝藻水华的暴发不存在绝对的营养盐限制.
由图7可知,监测期间香溪河库湾NO3--N浓度大致呈先降低后增加的趋势,结合图2可知,蓝藻水华期间NO3--N浓度显著降低,水华消退后NO3--N浓度恢复到较高水平,说明蓝藻的增殖需要消耗大量的无机氮营养,同时由于反硝化作用也会消耗大量的氮营养.监测期间正磷酸盐PO43--P浓度整体呈波动降低的趋势,变幅较大.SiO2-Si浓度在监测期间变化幅度不大,说明蓝藻增殖期间对硅营养消耗较少.
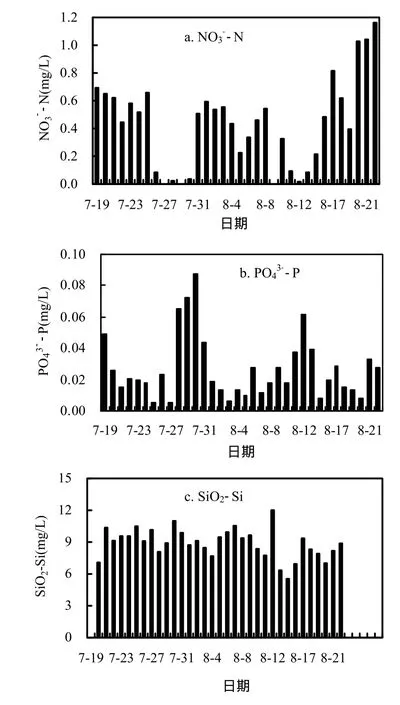
图7 表层NO3--N, PO43--P、SiO2-Si浓度随时间的变化Fig.7 NO3--N, PO43--P and SiO2-Si concentrations of surface water in Xiangxi Bay
3 讨论
3.1 蓝藻水华暴发相关分析
影响蓝藻水华暴发的环境因子很多,且不同因子在水华暴发过程中所起的作用大不相同,其中温度、水体光学特性、水动力特性、营养盐浓度被大多数研究认为是直接促使蓝藻形成水华的重要条件[25].水华的发生不是瞬时的,而是浮游植物生物量在水体中不断增加的过程[26].一般认为水华的暴发是适宜的水温、光照、水动力条件和充足的营养共同作用的结果.监测过程中香溪河水流流速缓慢,营养盐含量丰富[24],总氮、总磷含量均大于富营养化阀值,因此可以推测水温、Zmix/Zeu是诱发蓝藻水华的主要控制因子.
相关分析表明,整个监测期间蓝藻密度与水温、水体稳定系数呈显著正相关关系,与混合层/真光层(Zmix/Zeu)、混合层、硝酸盐浓度、TN/TP呈显著负相关关系.蓝藻生长的主要控制因子是水温、Zmix/Zeu、水体稳定系数.
3.2 蓝藻水华影响因子
水温与蓝藻密度具有显著的正相关关系,研究表明[27],在营养物质充足、水体较为稳定等条件具备时,适宜的高水温是蓝藻生长旺盛的外界诱发因子.监测发现,蓝藻密度对水温有很好的响应,随着温度的升高,蓝藻光合作用速率加快,丰度增加趋势明显,在高水温的诱发下蓝藻大量聚集在表层形成明显的水华现象.绿藻在温度较高时虽然可以形成水华[2],但当温度维持较高水平时,鱼腥藻可以产生藻毒素来抑制绿藻生长增殖[28].同时其他藻类在 25℃以上时生长速率缓慢[29],耐受不住高水温,因此高温条件下蓝藻更具有竞争优势暴发水华.可以推测,蓝藻增殖成为优势种群和蓝藻水华发生是高水温的诱发结果.
香溪河库湾夏季的持续高温,营养物质充足,藻类的生长更多的取决于接受有效光照的几率,这与混合层与真光层之比密切相关;由于香溪河库湾光照强度较高,加之水体上下混合作用微弱,减小了混合层(藻类可以悬浮分布的水层范围)和真光层(藻类维持宏观净增长的水层范围)的比值,有助于具有悬浮机制的鱼腥藻在真光层中自由迁移,以便接受充足光照、营养盐迅速繁殖而形成水华[30].随着水体掺混剧烈,混合层厚度逐渐增大,分层不明显,表层混合水体易将藻类携带至真光层以下,降低藻类接受充足光照的几率,增大藻类在真光层下的死亡率[31],则浮游植物生长增殖受到限制[32].对比图3、图6可知,Zmix/Zeu与蓝藻密度变化呈相反趋势,水华期间Zmix/Zeu小于 1,水柱垂向混合的范围在真光层深度以上,藻类进行充分光合作用并超过呼吸作用消耗,有利于水华的暴发,同时在Zmix/Zeu为0.5左右时,水华强度最大;水华后期Zmix/Zeu大于1,水体紊动剧烈,水柱混合均匀,混合层深度显著增加,在混合过程中,聚集在表层的藻类易被携带至下部水体,在混合层内的藻类会受到光照和营养盐的限制,降低水体初级生产力,水华消退.

表1 环境因子与蓝藻密度相关系数Table 1 Correlation coefficients between some environmental factors and cell density in Xiangxi Bay during the summer
鱼腥藻属固氮藻类,当周围氮源浓度降低时,鱼腥藻的部分营养细胞会转变为含固氮酶的异形胞,起到固氮作用,是较强的氮竞争者[28].监测期间,鱼腥藻对硝氮利用明显,而氮浓度的降低更加显现了鱼腥藻的竞争优势,从而更好的进行生长增殖.
研究表明,风对藻类的迁移有一定的影响,过大的风浪作用不利于藻类的富集.风的剪切、拖拽作用引起表层水体的扰动掺混,会一定程度加深Zmix深度.本研究中最大风速为4.2m/s,平均风速为1.1m/s,且风速大小变化不显著,风向以偏南风为主.香溪河库湾地形狭窄且曲折,吹程通常不超过2km[24],较之水温分层的变化,风对香溪河库湾水体混合层深度的影响有限.对于深水型水库,藻密度的分布受控于水动力扰动强度,水体垂向混合程度比风速对藻类垂向混合影响更为显著.监测期间,Zeu深度随时间变化较小, 因此,Zmix深度的变化直接决定着支流库湾水华的生消,极小的水体混合层深度导致了夏季蓝藻水华占优.异重流现象是导致 Zmix深度变化的主要影响因素,根据纪道斌等[33]的研究结果,可以通过三峡水库潮汐调度来改变水位波动方式,从而改变支流分层异重流的潜入方式,打破支流水温分层状态,进而迅速扩大支流库湾的Zmix深度,使Zmix深度远大于 Zeu深度,影响水下垂向光热条件,可促使蓝藻水华消退.
4 结论
4.1 夏季香溪河库湾暴发了严重的蓝藻水华,优势藻种为鱼腥藻,水华期间鱼腥藻通过自身悬浮机制、固氮机制增加获取更多光照、营养盐的几率,同时因其能够产生藻毒素,可以抑制其他藻类生长,增加其竞争优势,从而更好的进行生长增殖.
4.2 研究期间,香溪河库湾水体存在显著分层,氮磷营养含量充足,水华期间蓝藻对硝氮利用显著,在营养物质充足、库湾水体稳定等条件具备的情况下,适宜的水温是影响蓝藻生长的关键因子;水温的持续升高、Zmix/Zeu减小是蓝藻显著增殖并暴发水华的诱发因子并且在 Zmix/Zeu为 0.5左右时最有利于蓝藻的生长.水华后期水体表层紊动剧烈,打破库湾水温分层状态,迅速扩大 Zmix深度,导致 Zmix/Zeu增大从而限制藻类生长增殖,水华消退.
[1] 邱光胜,胡 圣,叶 丹,等.三峡库区支流富营养化及水华现状研究 [J]. 长江流域资源与环境, 2011,20(3):311-316.
[2] 张 敏,蔡庆华,王 岚,等.三峡水库香溪河库湾蓝藻水华生消过程初步研究 [J]. 湿地科学, 2009,7(3):230-236.
[3] 许秋瑾,郑丙辉,朱延忠,等.三峡水库支流营养状态评价方法[J]. 中国环境科学, 2010,30(4):453-457.
[4] 李 哲,郭劲松,方 芳,等.三峡小江回水区蓝藻季节变化及其与主要环境因素的相互关系 [J]. 环境科学, 2010,31(2):301-309
[5] 易仲强,刘德富,杨正健,等.三峡水库香溪河库湾水温结构及其对春季水华的影响 [J]. 水生态学杂志, 2009(005):6-11.
[6] 王光谦,方红卫.异重流运动基本方程 [J]. 科学通报, 1996, 41(018):1715-1720.
[7] 魏复盛,国家环境保护总局,水和废水监测分析方法编委会.水和废水监测分析方法 [M]. 北京:中国环境科学出版社, 2002.
[8] 胡鸿钧.中国淡水藻类 [M]. 上海:上海科学技术出版社, 1980.
[9] 周凤霞,陈剑虹.淡水微型生物图谱 [M]. 北京:化学工业出版社, 2005.
[10] Lawson R, Anderson M A. Stratification and mixing in Lake Elsinore, California: An assessment of axial flow pumps for improving water quality in a shallow eutrophic lake [J]. Water Research, 2007,41(19):4457-4467.
[11] 张书农.环境水力学 [M]. 南京:河海大学出版社, 1988.
[12] Lawrence I, Bormans M, Oliver R, et al. Physical and nutrient factors controlling algal succession and biomass in Burrinjuck Reservoir [Z]. Land and Water Resources Australia, 2000.
[13] 郑建军,钟成华,邓春光.试论水华的定义 [J]. 水资源保护, 2006,22(5):45-47.
[14] 金相灿,屠清瑛.环境科学. 湖泊富营养化调查规范 [M]. 北京:中国环境科学出版社, 1990.
[15] 王海云,程胜高,黄 磊.三峡水库 “藻类水华” 成因条件研究[J]. 人民长江, 2007,38(2):16-18.
[16] 陈宇炜,秦伯强,高锡云.太湖梅梁湾藻类及相关环境因子逐步回归统计和蓝藻水华的初步预测 [J]. 湖泊科学, 2001,13(1): 63-71.
[17] 高月香,张永春.水文气象因子对藻华爆发的影响 [J]. 水科学与工程技术, 2006(2):10-12.
[18] Zhang Y, Prepas E E. Regulation of the dominance of planktonic diatoms and cyanobacteria in four eutrophic hardwater lakes by nutrients, water column stability, and temperature [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1996,53(3):621-633.
[19] Steinberg C E W, Hartmann H M. Planktonic bloom‐forming Cyanobacteria and the eutrophication of lakes and rivers [J]. Freshwater Biology, 1988,20(2):279-287.
[20] Kawara O, Yura E, Fujii S, et al. A study on the role of hydraulic retention time in eutrophication of the Asahi River Dam reservoir [J]. Water Science and Technology, 1998,37(2):245-252.
[21] 江启明,侯 伟,顾继光,等.广州市典型中小型水库营养状态与蓝藻种群特征 [J]. 生态环境学报, 2010,19(010):2461-2467.
[22] Khanna D R, Bhutiani R, Chandra K S. Effect of the euphotic depth and mixing depth on phytoplanktonic growth mechanism [J]. International Journal of Environmental Research, 2009, 3(2):223-228.
[23] Davis J R, Koop K. Eutrophication in Australian rivers, reservoirs and estuaries–a southern hemisphere perspective on the science and its implications [J]. Hydrobiologia, 2006,559(1):23-76.
[24] Liu L, Liu D, Johnson D M. Effects of vertical mixing on phytoplankton blooms in Xiangxi Bay of Three Gorges Reservoir: Implications for management [J]. Water Research, 2012,1(1): 2121–2130
[25] Havens K E, James R T, East T L, et al. N: P ratios, light limitation, and cyanobacterial dominance in a subtropical lake impacted by non-point source nutrient pollution [J]. Environmental Pollution, 2003,122(3):379-390.
[26] 孔繁翔,高 光.大型浅水富营养化湖泊中蓝藻水华形成机理的思考 [J]. 生态学报, 2005,25(3):589-595.
[27] 王 炜,余卫东.千岛湖蓝藻生物量制约因素分析 [J]. 云南环境科学, 2002,21(4):44.
[28] 戴瑾瑾,陈德辉,高云芳,等.蓝藻毒素的研究概况 [J]. 武汉植物学研究, 2009,27(001):90-97.
[29] 史小丽,王凤平,蒋丽娟,等.扰动对外源性磷在模拟水生态系统中迁移的影响 [J]. 中国环境科学, 2002,22(6):537-541.
[30] Jones G J, Poplawski W. Understanding and management of cyanobacterial blooms in sub-tropical reservoirs of Queensland, Australia [J]. Water Science and Technology, 1998,37(2):161-168.
[31] Doyon P, Klein B, Ingram R G, et al. Influence of wind mixing and upper-layer stratification on phytoplankton biomass in the Gulf of St. Lawrence [J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2000,47(3/4):415-433.
[32] Whitton B A, Potts M. The ecology of cyanobacteria: their diversity in time and space [M]. Amsterdam. Kluwer Academic Publishers, 2000.
[33] 纪道斌,刘德富,杨正健,等.汛末蓄水期香溪河库湾倒灌异重流现象及其对水华的影响 [J]. 水利学报, 2010,6:691-702.
Cyanobacterial bloom in Xiangxi Bay, Three Gorges Reservoir.
TIAN Ze-bin1, LIU De-fu1,2*, YANG Zheng jian1,3, FANG Xiao-feng1, YAO Xu-jiao1, FANG Li-juan1,2(1.Engineering Research Center of Eco-environment in Three Gorges Reservoir Region, Ministry of Education, College of Hydroelectric and Environmental Engineering, China Three Gorges Univerxity, Yichang 443002, China;2.College of Resources and Environment Sciences, Hubei University of Technology, Wuhan 430072, China;3.College of Resources and Environment Sciences, Hubei University of Technology, Wuhan 430072, China). China Environmental Science, 2012,32(11):2083~2089
A cyanobacteria bloom was observed in Xiangxi Bay of Three Gorges Reservoir in summer of 2010. Based on field data collected from 19 July to 30 August 2010, this paper descripted the cyanobacteria bloom process and analyzed the influence factors. The bloom lasting for about 25 days, broke out on 23 July, and ended on 16 August, cell density summit reached 108.03×106cell/L. The dominant species was Anabaena, and the major reasons were the suspension and nitrogen fixation mechanism, along with the capacity to produce microcystin to inhibit the growth of other algal. Specific stratification model of water and adequate nutrients were necessary to the cyanobacteria bloom, and nitrate might be the major nitrogen source. Under the conditions of undisturbed water situation and adequate nutrients, the proliferation rate of cyanobacteria might be accelerated gradually with increasing water temperature and lowering Zmix/Zeu. Especially when Zmix/Zeuwas less than 0.5, it was more favorable to cyanobacteria growth.
Three Gorges Reservoir;Xiangxi Bay;cyanobacteria bloom;mixing depth
2012-02-25
国家水体污染控制与治理科技重大专项(2009ZX07104-001,2009ZX07104-004,2009ZX07104-003-03);“十一五”国家科技支撑计划(2008BAB29B09)
* 责任作者, 教授, dfliu@ctgu.edu.cn
X524
A
1000-6923(2012)11-2083-07
田泽斌(1989-),女,河北秦皇岛人,三峡大学硕士研究生,主要从事生态水工方面的研究.
猜你喜欢
当代水产(2021年8期)2021-11-04
环境科学导刊(2021年2期)2021-01-09
当代水产(2019年8期)2019-10-12
当代水产(2019年9期)2019-10-08
当代水产(2018年8期)2018-11-02
作文与考试·初中版(2017年20期)2017-07-08
中华诗词(2016年10期)2017-02-22
创新作文(1-2年级)(2016年4期)2016-05-14
幼儿智力世界(2015年5期)2015-08-20
小猕猴智力画刊(2013年1期)2013-03-15
