
短乳杆菌发酵生产尿苷磷酸化酶条件的优化
2013-02-13 08:16王伟洁李红梅邓龙华高露姣黄艳青
食品科学 2013年17期
王伟洁,李红梅,*,邓龙华,高露姣,黄艳青
(1.上海理工大学医疗器械与食品学院,上海 200093;2.中国水产科学研究院东海水产研究所,上海 200090)
尿苷磷酸化酶是嘧啶补救途径中的关键酶,具有可逆性催化尿苷转变为尿嘧啶的异化和磷酸化作用,广泛存在于生物有机体中。尿苷磷酸化酶在酶法合成抗肿瘤核苷类药物中具有重要作用[1],其中脱氧氟尿苷DFUR是一种良好的抗代谢类抗肿瘤药物,在肿瘤细胞内受肿瘤组织中嘧啶磷酸化酶的作用而转化为真正起作用的5-氟尿嘧啶;此外尿苷磷酸化酶在肿瘤诊断中具有一定医学价值,肿瘤组织细胞中尿苷磷酸化酶的含量比正常细胞高很多,可以作为细胞癌变的检测指标[2]。
目前通过传统筛选和诱变技术获得了一些具有较高酶活力的菌种如产气肠杆菌、乙酰短杆菌等应用于核苷类似物的酶法合成[3],但关于短乳杆菌产核苷磷酸化酶却鲜有报道,野生型短乳杆菌仍存在产酶量相对较低等缺点,因此如何提高短乳杆菌产核苷磷酸化酶具有重要的研究和应用价值。
现有筛选重要影响因子的方法中,以Plackett-Burman设计最具优势,它能快速有效地从众多因子中筛选出影响短乳杆菌产尿苷磷酸化酶显著的因子[4-5]。响应面法能快速对影响显著因子进行优化与评价,并确定各因素最佳组合和响应值的最优值[6]。目前响应面分析方法已大量应用于微生物工程领域的实验分析中[7-9]。因此,以短乳杆菌为出发菌,以葡萄糖酵母膏培养基为基础培养基,利用Plackett-Burman设计、最陡爬坡试验及响应面法对影响酶活的各因子水平及其交互作用进行优化与评价,并快速有效地确定多因子系统的最佳条件,旨在为高产尿苷磷酸化酶菌株的开发、应用以及产酶发酵条件优化等相关研究及后续研究提供一定的参考。
1 材料与方法
1.1 菌株
短乳杆菌 本实验室保存。
1.2 试剂
胸苷、肌苷、尿苷、尿嘧啶等 河南新乡拓新生化股份有限公司;其他化学试剂(均为分析纯) 国药集团药业公司。
1.3 培养基及培养条件
1.3.1 培养基
菌种活化液(g/L):酵母粉10.0,pH7.0~7.2;斜面培养基(g/L):葡萄糖20.0、酵母粉10.0、氯化钠5.0、琼脂20.0,pH7.0~7.2;初始发酵培养基(g/L):葡萄糖20.0、酵母粉10.0、氯化钠5.0,pH7.0~7.2;发酵培养基:根据不同实验设计要求,改变培养基的各组分。
1.3.2 培养条件
将保存于深冷冰箱的短乳杆菌接种在固体斜面培养基上36℃培养24h,4℃冰箱中保存备用。无菌环境下从斜面培养基上挑取单一菌落菌体接种到装液量为50mL的250mL锥形瓶活化液中,在36℃摇床转速为110r/min条件下活化10~12h,然后接种到发酵培养基中,同样条件下继续培养12h收集菌体。
1.4 酶活力测定
1.4.1 湿菌体的收集
将培养好的菌液4500r/min常温离心15min,去除上清液后,用无菌水和pH7.3无菌磷酸钾缓冲液各冲洗1次,离心去上清,收集湿菌体冷藏备用。
1.4.2 尿苷磷酸化酶活测定方法
据Saunders等[10]报道,采用紫外分光光度法测定反应生成的尿嘧啶来表征产酶量。标准酶反应混合液包含一定浓度的湿菌体50mg/mL反应液,25mmol/L尿苷,1mmol/L EDTA,pH7.3、50mmol/L的磷酸钾缓冲溶液。55℃条件下反应120min,反应结束后,反应液在沸水中煮沸5min终止反应,离心去除沉淀[11]。上清液用pH12的NaOH稀释100倍,以未添加菌体的反应液作为对照测定不同培养基及培养条件下反应液在290nm波长处的紫外光密度OD290nm的增值[12]。尿苷磷酸化酶酶活单位定义为:在上述条件下,1min内OD290nm变化0.01所需的湿菌体量定义为短乳杆菌的1个酶活力单位[3]。

式中:K为稀释倍数,取100;t为酶反应时间(120min);m为湿菌体质量(50mg)。
1.5 发酵培养基的优化
1.5.1 单因素试验
在基础发酵培养基中,分别添加适量的速效氮源蛋白胨、金属离子Mg2+(MgCl2·6H2O)、磷酸根(K2HPO4·3H2O)、底物诱导剂(胸苷、肌苷及尿苷),考察其对短乳杆菌产尿苷磷酸化酶的作用。
1.5.2 Plackett-Burman试验设计
根据文献查阅及前期单因素试验研究[13-14],选取发酵温度、发酵时间、摇床转速、接种量、葡萄糖、酵母粉、蛋白胨、磷酸根、尿苷和肌苷作为Plackett-Burman设计的10个因素,以尿苷磷酸化酶活作为响应值,每个因素取两个水平,高水平(+1)约是低水平(-1)的1.5~2倍,另设3个虚拟列:X3、X6、X10以考察试验误差。通过比较各因素的显著水平,筛选出对酶活力影响较为显著的因素。试验设计见表1。

表1 Plackett-Burman试验设计方案Table 1 Factors and levels used in Plackett-Burman design
1.5.3 最陡爬坡试验
根据Plackett-Burman试验结果分析确定显著影响因素,以拟合的回归方程模型的系数符号和大小来设定显著因素的步长和变化方向,即对显著因素进行浓度梯度设计,使响应值快速逼近最大响应区间。
1.5.4 Box-Behnken设计
根据Box-Behnken中心组合试验设计原理[15-16],以葡萄糖、磷酸根、尿苷、酵母粉4个因素为自变量,以尿苷磷酸化酶活为响应值,设计四因素三水平的响应面分析试验,并用Design-Export 8.0软件对试验数据进行回归分析。
2 结果与分析
2.1 单因素试验筛选
利用底物诱导(胸苷、肌苷和底物尿苷)、添加金属离子(MgCl2·6H2O)、速效氮源(蛋白胨)和磷酸盐(K2HPO4·3H2O)等手段提高短乳杆菌产尿苷磷酸化酶能力[13],结果如表2所示。可以看出,蛋白胨、磷酸根、肌苷和尿苷4个因素对短乳杆菌合成尿苷磷酸化酶有一定的促进作用。蛋白胨为菌体生长提供充足的氮源,提高菌体单位时间内的丰度;磷酸根是尿苷磷酸化酶催化尿苷可逆磷酸化反应的有效因子,可能会促进菌体产尿苷磷酸化酶;而核苷、核苷酸及碱基对核苷磷酸化酶有一定的诱导作用,但不同微生物对诱导剂的响应不同,E.coli中尿苷磷酸化酶可由胞苷诱导表达[14],Salmonella typhimurium中尿苷磷酸化酶由尿苷和胞苷诱导[17],本研究发现,尿苷和肌苷可诱导短乳杆菌产尿苷磷酸化酶。

表2 单因素试验设计方案及结果Table 2 Single factor design and corresponding results
2.2 Plackett-Burman设计及优化
选用Factors=10,Runs=20的Plackett-Burman设计,见表3。根据表3结果采用Minitab15.0软件进行方差分析,结果(表4)表明,在发酵过程中,肌苷、发酵时间、酵母粉、磷酸根、蛋白胨及底物尿苷6个因素对短乳杆菌合成尿苷磷酸化酶呈正效应关系,而摇床转速、接种量、发酵温度及葡萄糖添加量4个因素对短乳杆菌合成尿苷磷酸化酶呈负效应关系。

表3 Plackett-Burman试验设计及结果Table 3 Plackett-Burman design arrangement and corresponding results
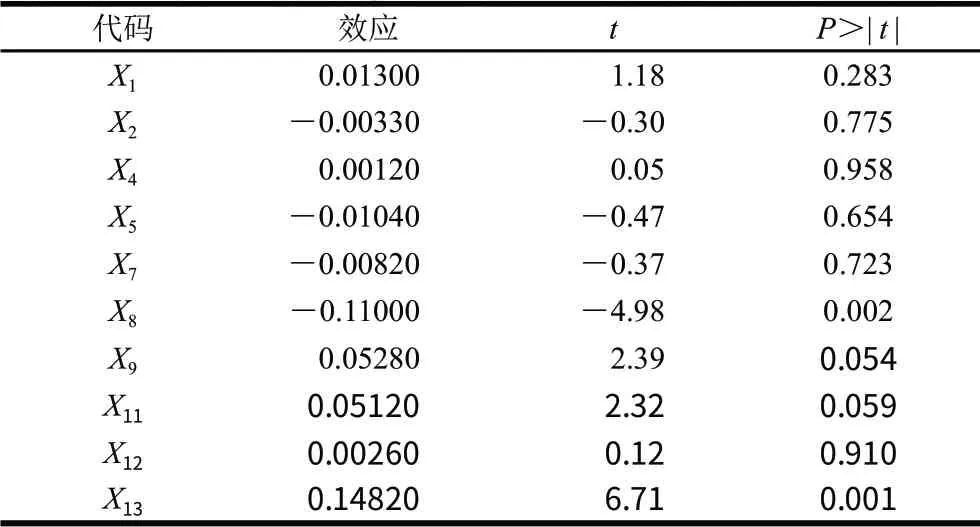
表4 Plackett-Burman试验结果的方差分析Table 4 Analysis of variance for the Plackett-Burman experimental design
在Plackett-Burman设计中,一般认为P<0.1的因素对系统的响应值影响较为显著,本研究中葡萄糖(P=0.002)、酵母粉(P=0.054)、磷酸根(P=0.059)和尿苷(P=0.001)对短乳杆菌合成尿苷磷酸化酶的影响较为显著,因此将这4个因素作为主要因素进行最陡爬坡试验和响应面试验。其他因素正效应取高水平,负效应取低水平。其中葡萄糖和尿苷浓度对短乳杆菌产尿苷磷酸化酶影响非常显著(P<0.01),葡萄糖添加量过低,导致营养不足,添加量过高,可能会出现葡萄糖限制或其产物的抑制,均不利于尿苷磷酸化酶的产生[13];尿苷可以促进尿苷磷酸化酶活的试验结果可以初步判断尿苷磷酸化酶为诱导酶,添加酶底物尿苷可以显著提高短乳杆菌产酶量,这与Salmonella typhimurium[17]、Klebsiellasp.[18]产尿苷磷酸化酶可被尿苷诱导表达研究一致。
2.3 最陡爬坡试验结果
Plackett-Burman试验结果显示葡萄糖为负效应,在后续设计中添加量应减小;酵母粉、磷酸根添加量和尿苷浓度3个因素均为正效应,在后续的设计中添加量应增加。因此最陡爬坡试验设计应根据各个因素效应大小比例,来设定它们的变化步长,如表5所示,短乳杆菌产尿苷磷酸化酶能力在第3组达到最大值(1.198U/mg),因此,以第3组作为响应面试验的中心点。

表5 最陡爬坡试验设计及结果Table 5 Steepest ascent design and corresponding results
2.4 Box-Behnken响应面试验结果及方差分析
Box-Behnken试验设计与结果见表6,对试验结果用Design Export 8.0响应面分析软件进行多元回归分析,结果见表7。经拟合得到二次回归数学模型,获得短乳杆菌产尿苷磷酸化酶能力对葡萄糖、磷酸根、尿苷及酵母粉添加量的二次多项回归方程为:
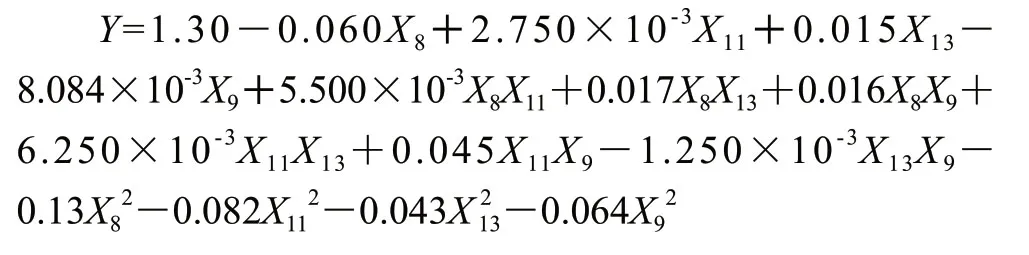
由该方程的方差分析可见,该模型极显著(P<0.0001),失拟度在α=0.05水平上不显著(P=0.0584>0.05),而且经分析计算,该模型的确定系数R2=0.9183,表明模型与实际情况拟合很好,因此该模型可用于预测短乳杆菌产尿苷磷酸化酶能力的实际情况[19]。由回归方程得到优化结果为葡萄糖18.30g/L、磷酸根3.99g/L、尿苷21.38mmol/L、酵母粉15.71g/L,预测响应面最大值为1.30887U/mg。
利用Design Export 8.0软件绘出方程的响应面图及等高线分析图,响应面分别代表两个独立变量之间对响应值尿苷磷酸化酶活力的影响,并可以直接地反映出最优值;等高线的形状可以反映两因素间交互作用的强弱大小,圆形表示交互作用不显著,椭圆形表示交互作用显著。由图1可知,磷酸根-酵母粉的交互作用等高线呈椭圆形,表明磷酸根-酵母粉交互作用对尿苷磷酸化酶活影响显著,其他因素间交互作用影响不显著。

表6 Box-Behnken试验设计与结果Table 6 Box-Behnken design arrangement and corresponding results

表7 响应面回归模型的方差分析Table 7 Analysis of variance for the response surface regression model
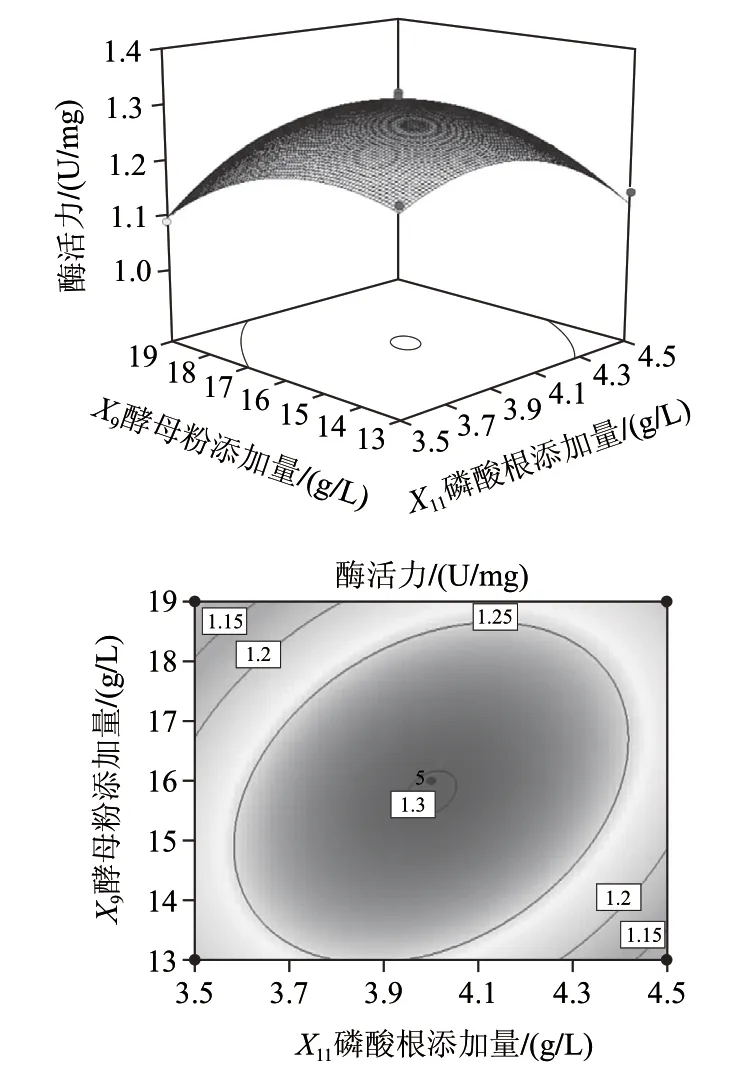
图1 磷酸根与酵母粉添加量相互作用对尿苷磷酸化酶产量的响应面立体分析图与等高线图Fig.1 Response surface and contour plots showing the interaction effects of various factors on the yield of uridine phosphorylase
2.5 模型方程的验证
响应面法分析得到的最优结果并没有在设计的试验组合中,为了进一步验证模型的可靠性,在最佳培养条件下,做5次重复实验,得到尿苷磷酸化酶活为1.325U/mg,基本与响应面预测的最大值1.30887U/mg符合,说明响应面法优化得到的数学模型与实验数据拟合较好。在基础发酵培养基发酵条件下,短乳杆菌产尿苷磷酸化酶能力为0.883U/mg,优化后提高了50.0%。
3 结 论
提高菌体产酶量最有效的方法主要是通过改良微生物产酶培养基。短乳杆菌产尿苷磷酸化酶过程中,影响产酶量的因素很多,且因素之间相互联系。本实验通过单因素试验确定了蛋白胨、磷酸根、尿苷及肌苷4个因素为促进短乳杆菌产尿苷磷酸化酶的有效因子,并通过Plackett-Burman试验筛选出影响尿苷磷酸化酶活的显著因子,进而利用最陡爬坡试验逼近最大响应区域,最后通过响应面法优化短乳杆菌产尿苷磷酸化酶发酵培养基配方为:NaCl 5g/L、葡萄糖18.30g/L、酵母粉15.71g/L、蛋白胨15g/L、磷酸根3.99g/L、肌苷20mmol/L、尿苷21.38mmol/L;发酵条件:pH8.0、摇床转速110r/min、发酵温度32℃、培养时间14h、接种量1.0%。在优化培养基的基础上进行验证实验,尿苷磷酸化酶活力达1.325U/mg,较优化前的酶活力0.883U/mg提高了50.0%。以上结果表明通过响应面法优化短乳杆菌产尿苷磷酸化酶发酵培养基及其发酵条件是可行有效的,这为进一步通过选育菌种、代谢调控等技术提高酶活提供了研究基础。
[1]徐渊, 应国情.通过大肠杆菌的核苷磷酸化酶合成核苷的研究[J].科技创新导报, 2008, 23(5): 22-23.
[2]丁庆豹.核苷磷酸化酶的克隆、表达与应用[D].上海: 华东理工大学, 2010: 15-17.
[3]谭黎, 欧阳立阳, 丁庆豹.大肠杆菌核苷磷酸化酶的重组表达和活性[J].华东理工大学学报, 2008, 24(5): 660-664.
[4]MILLER A, SITTER R R.Using the folded-over 12-Run Plackett-Burman design to consider interactions[J].Technometrics, 2001, 43(1):44-54.
[5]MUSSATTO S I, DRAGONE G, ROCHA J M, et al.Optimum operating conditions for brewer’s spent grain soda pulping[J].Carbohydrate Polymers, 2006, 64(1): 22-28.
[6]AMBATI P, AYYANNA C.Optimizing medium constituents and fermentation conditions for citric acid production from palmyra jaggery using response surface method[J].World Journal of Microbiology and Biotechnology, 2001, 17(4): 331-335.
[7]BARRINGTON S, KIM J W.Response surface optimization of medium components for citric acid production byAspergillus nigerNRRL 567 grown in peat moss[J].Bioresource Technology, 2008,99(2): 368-377.
[8]吴孔阳, 王学军, 周培华, 等.响应面法优化黑曲霉发酵产低聚异麦芽糖培养基[J].食品科学, 2012, 33(19): 216-220.
[9]刘松, 李祝, 周礼红, 等.响应面法优化黑曲霉产纤维素酶的发酵条件[J].食品科学, 2012, 33(5): 1-7.
[10]SAUNDERS P P, WILSON A B, SAUNDERS G F, et al.Purification and comparative properties of a pyrimidine nucleoside phosphorylase fromBacillus stearothermophilus[J].The Journal of Biological Chemistry, 1969, 244(13): 3691-3697.
[11]DING Qingbao, OU Ling, WEI Dongzhi, et al.Induction of recombinant uridine phosphorylase and its application in biosynthesis of pyrimidine nucleosides[J].Chinese Journal of Chemical Engineering, 2011, 19(1): 122-127.
[12]魏晓琨, 张春艳, 丁庆豹, 等.产气肠杆菌菌体内核苷磷酸化酶酶活测定条件的分析[J].中国临床医药研究杂志, 2007(13): 4-7.
[13]倪孟祥, 葛亚文, 吴梧桐.影响产气肠杆菌EAM-Z1中嘧啶核苷磷酸化酶活力的因素[J].中国医科大学学报, 2007, 38(6): 558-561.
[14]魏晓琨, 刘岩岩, 周丽娜, 等.产气肠杆菌体内核苷磷酸化酶的诱导表达[J].现代生物医学进展, 2008, 8(2): 255-259.
[15]GAWANDE N B, SONAWANE M A, JOGDAND V V, et al.Optimization of cyclodextrin glycosyltransferase production fromKlebsiella pneumoniaeAS-22 in batch, fed-batch and continuous cultures[J].Biotechnology Progress, 2003, 19(6): 1697-1702.
[16]ROSSO M A, FERRAROTTI A S, KRYMKIEWICZ N,et al.Optimization of batch culture conditions for cyclodextrin glucanotransferase production fromBacillus circulansDF 9R[J].Microbial Cell Factories, 2002, 52(9): 3-12.
[17]ROGERT M C, TRELLES J A, PORRO S, et al.Microbial synthesis of antiviral nucleosides usingEscherichia coliBL 21 as biocatalyst[J].Biocatalysis and Biotransformation, 2002, 20(5): 347-351.
[18]ZHANG Shaotan, NI Mengxiang, RUAN Qiping.Enzymatic synthesis of nucleoside drugs[J].Progress in Pharmaceutical Sciences, 2005, 2:56-62.
[19]吴孔阳, 王学军, 周培华, 等.响应面法优化黑曲霉发酵产低聚异麦芽糖培养基[J].食品科学, 2012, 33(19): 216-220.
猜你喜欢
阅读(快乐英语中年级)(2022年5期)2022-06-03
当代水产(2022年1期)2022-04-26
江西水产科技(2019年3期)2019-09-10
天津农业科学(2019年4期)2019-05-31
中成药(2017年5期)2017-06-13
中成药(2017年5期)2017-06-13
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
中国当代医药(2015年9期)2015-03-01
肝博士(2015年2期)2015-02-27
