
氮添加对高寒草甸植物花期物候和群落结构的影响
2013-03-14 04:44章志龙刘旭东杜国祯
草业科学 2013年5期
章志龙,刘旭东,贾 鹏,杜国祯,
(1.兰州大学草地农业科技学院,甘肃 兰州 730020; 2.兰州大学生命科学学院,甘肃 兰州 730000)
由于人类活动的影响,氮素进入陆地生态系统的方式和速率发生着极大变化,人为固定的氮素正在不断积累,并对生态系统的结构和功能产生显著影响[1-3]。氮素在陆地生态系统中的增加引起了土壤生物化学性质及生物多样性的变化[4],草地植物群落中氮素的增加可导致群落物种多样性和群落结构的变化[5-8]。近年来的研究表明,氮素的增加会引起植物物候的变化,尤其是植物花期的变化[9-11],以及对植物的繁殖策略产生影响[12-13]。植物繁殖的时间体现了营养生长与生殖生长的平衡[14-15],其中花期紧密地连接着营养生长和之后的生殖生长。植物的许多生理生态特征在花期集中体现出来,反映了植物在特定环境下的适合度[16-17],因此,花期的变化可视为植物应对环境变化、避免干扰的核心属性[18-19]。花期物候决定着后代群落的动态结构,影响着群落的进化过程[20]。尽管已有研究表明,环境变化对植物物候产生了较大影响,然而目前关于环境变化对花期物候的影响尚无定论[21-24]。
关于草地群落土壤环境因素对植物群落结构、生产力和多样性影响的研究已比较多[5-8],但涉及氮素添加对草地群落物候影响的研究则相对较少[11,25]。本研究以青藏高原东部高寒草甸植物为例,分析了高寒草甸植物群落中30个常见组分种的始花期,群落性状和结构对氮素添加的响应,以期确定高寒草甸群落组分种的始花期是否对氮素添加产生响应,揭示氮素添加导致始花期变化的原因,以及物候的变化对群落结构的影响。
1 材料与方法
1.1研究地概况和样地描述 研究地位于青藏高原东缘玛曲县境内兰州大学高寒草甸与湿地生态系统定位研究站(阿孜分站)(33°40′ N,101°52′ E),海拔 3 550 m。年均温1.2 ℃(1月均温-10 ℃,7月均温为11.7 ℃),无霜期小于90 d。过去30年年均降水量为670 mm,主要集中在短暂的夏季。主体土壤类型为亚高山草甸土,植被属于高寒草甸类。样地植被类型为莎草类+禾草类+双子叶杂草类群系,莎草类主要以线叶嵩草(Kobresiacapillifolia)为主;禾草类主要有冷地早熟禾(Poapoophagorum)、垂穗披碱草(Elymusnutans)、胡氏剪股颖(Agrostishugoniana)和草地早熟禾(P.pratensis)等;杂类草中常见的有毛茛科的钝裂银莲花(Anemoneobtusiloba)、条叶银莲花(A.trullifolia)、小花草玉梅(A.rivularis)以及丝叶毛茛(Ranunculustanguticus)。此外,还有豆科植物多枝黄耆(Astragaluspolycladus)、高山豆(Tibetiahimalaica)和黄花棘豆(Oxytropisochrocephala),菊科的长毛风毛菊(Saussureahieracioides)、瑞苓草(S.nigrescens)和重冠紫菀(Asterdiplostephioides)等。样地自2008年起实行夏季禁牧。
1.2试验设计 氮素添加试验采用随机区组设计,在围栏内选择一块地势平坦且匀质的样区,分成15个10 m×6 m的小样区,小样区四角用木桩标记,各小样区间距为2 m。在每个小样区内设置4个0.5 m×0.5 m的固定样方用于群落测定和物候观测。设置了3个氮素添加水平,分别为对照(CK,0 g·m-2)、低氮(LN,5 g·m-2)和高氮(HN,10 g·m-2),所用含氮试剂为硝酸铵(NH4NO3),3个氮素添加水平随机的布置在不同小样区内[26]。2009-2011年,于每年5月上旬在多云阴天时将硝酸铵按照以上设计的氮素添加水平均匀的撒在每个小样区内。
1.3物候观测和群落性状调查 植物群落物候观测于2009-2011年每年的5月上旬-9月下旬3个完整的生长季内进行。每次物候观测间隔为5~7 d,记录样方内出现的所有物种及其物候期,每次记录独立于上一次[27],并在花期测量各物种的高度。记录不同氮素添加小区内用于物候观测的样方中出现的每一个物种的始花期(First Flowering Date)。在每次观测中,当物候观测样方中的双子叶植物出现可辨认的花瓣、花药和花丝时,则认为该植物进入花期;当禾草类植物的小穗只要不再被叶鞘包裹时,则认为禾草进入花期,并以儒略历(Julian day,1月1日为1年中第1天)计数法计时[28]。在采样区对不同氮素添加下处于花期的各物种的植物个体进行采样,每一个物种采集20株完整的地上部分,从每一株上选取1~2片健康未破损的叶子,用扫描仪扫描叶片测定叶面积,测定完毕后在烘箱内烘干(80 ℃,48 h),计算比叶面积(SLA=叶面积/叶干重,cm2·g-1)。每年8月底在每个样方测定群落结构,记录每个0.5 m ×0.5 m样方内物种丰富度、物种的多度和分盖度。
1.4数据分析 用群落中各物种的相对多度计算辛普森多样性指数(Simpson’s Diversity Index,D):
式中,S为群落中的物种数目,Pi为群落内任意物种的相对多度。
用物种的相对多样性加权计算群落花期植株高度(CWM-H)和群落比叶面积(CWM-SLA)两个性状(Trait),按照Garnier等[29]的方法计算物种相对多样性加权的群落特征值(Community Weighted Means):
式中,n为物种数,traiti为物种i的特征值。
用单因素方差分析(One-way ANOVA)检验氮素添加对30个物种的始花期和花期高度的影响,以及氮素添加对群落特征CWM-H和CWM-SLA的影响,用多重比较中的最小显著差数法(LSD Method)比较3个氮素添加水平之间差异(P<0.05)。用非度量多维尺度分析(NMDS)检验氮素添加对开花物候加权的群落结构相似性的影响。统计分析和绘图使用SPSS 16.0和 R 2.8.1统计软件进行。
2 结果与分析
2.1氮素添加对主要组分种始花期和花期物候及植株高度的影响 莎草科线叶嵩草,禾本科冷地早熟禾、垂穗鹅观草(Roegnerianutans)、垂穗披碱草、草地早熟禾、羊茅(Festucaovina)、胡氏剪股颖、芒剪股颖(A.trinii)在高氮处理下的始花期显著迟于对照(P<0.05),而在低氮处理下,除羊茅与对照无显著差异外(P>0.05),其余物种的始花期也显著推迟(P<0.05)。具有固氮能力的豆科植物多枝黄耆、高山豆和黄花棘豆的始花期在氮素添加后均无显著差异。双子叶类菊科杂草长毛风毛菊、瑞苓草和重冠紫菀在高氮处理下始花期显著迟于对照(P<0.05),在低氮处理下,瑞苓草的始花期显著推迟(P<0.05)。与对照相比,湿生扁蕾(Gentianopsispaludosa)、椭圆叶花锚(Haleniaelliptica)、刺芒龙胆(Gentianaaristata)、甘肃马先蒿(Pediculariskansuensis)、短腺小米草(Euphrasiaregelii)、丝叶毛茛和条叶银莲花在高氮处理下的始花期显著提前(P<0.05),在低氮处理下的始花期亦提前但并不显著。其余的双子叶类杂草的始花期在氮素添加下与对照群落无显著差异(表1,表2)。在氮素添加下,30个常见物种在科间的始花期变化总体上表现为禾本科、莎草科和菊科的始花期推迟,而毛茛科、龙胆科和玄参科的始花期提前,且高氮处理下的变化大于低氮处理。此外,豆科始花期无变化(图1)。30个常见物种的花期植株高度随着氮素添加量的增加而增加(表2),莎草科和禾本科所有物种的花期植株高度在氮素添加下增加,双子叶类杂草中除刺芒龙胆和高山豆与对照无差异外,其余皆与对照差异显著 (P<0.05)。

表1 氮素添加对高寒草甸植物群落中30个常见物种始花期的影响Table 1 Effects of nitrogen addition on the first flowering date of 30 common species d
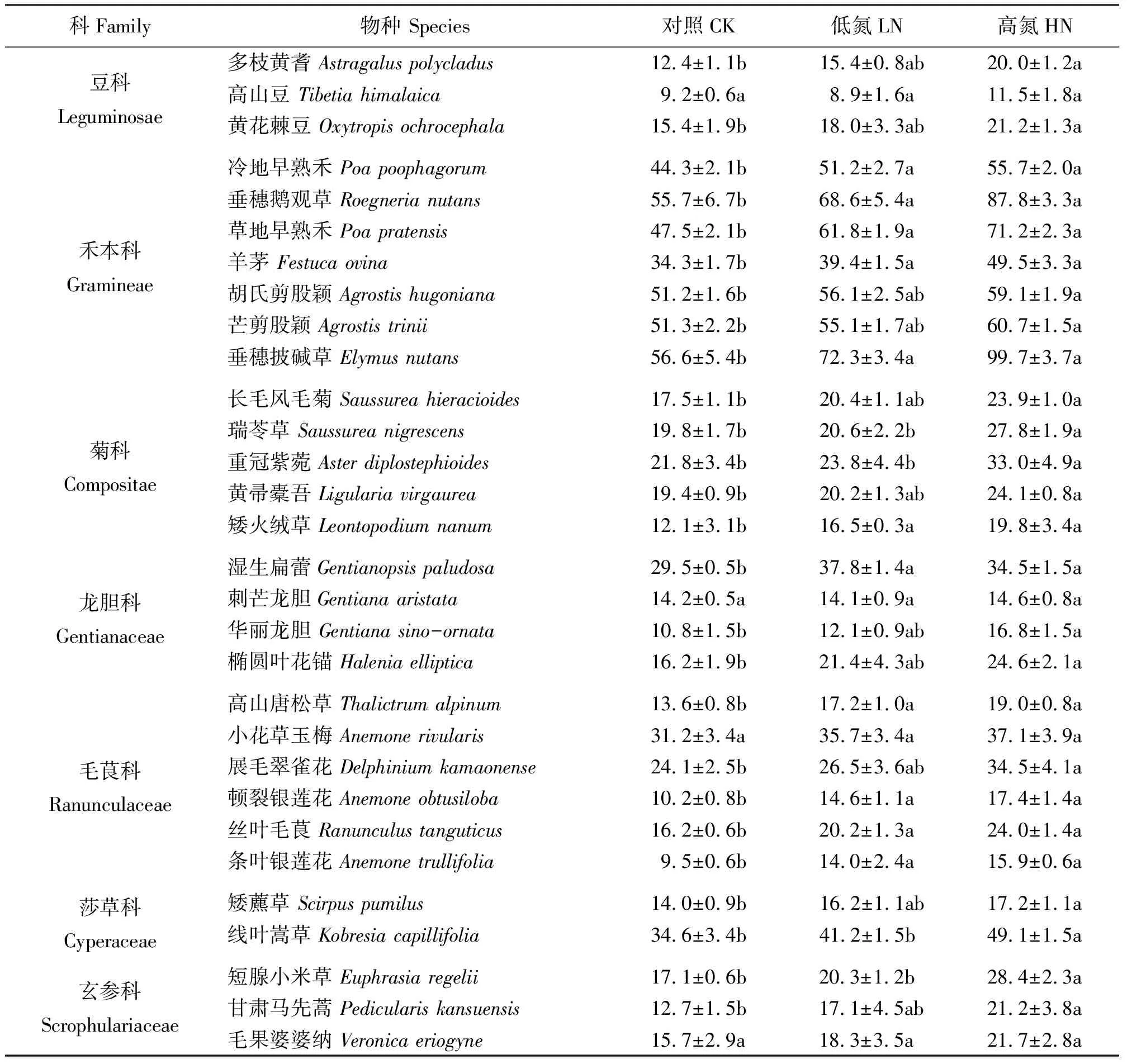
表2 氮素添加对高寒草甸植物群落中30个常见物种植株高度的影响Table 2 Effects of nitrogen addition on plant height of 30 common species cm
2.2氮素添加对群落性状和群落花期物候和群落结构的影响 氮素添加下的群落植株高度和群落比叶面积都显著高于对照(P<0.05)(图2)。随着氮素添加量的增加,多样性指数降低,高氮处理下降低显著(P<0.05) (图3)。自然群落与氮素添加下的群落花期物候相似性和群落结构的比较如图4所示,3条不同的曲线分别表示了非度量多维尺度分析(NMDS)排序呈现出的3个氮素处理(CK、LN和HN)下群落之间在群落花期物候和群落结构的相似性分离程度(Stress Value=0.08)。对照、低氮和高氮群落之间的相似性逐渐降低,对照群落与低氮群落的相似性大于对照群落与高氮群落的相似性,说明随着氮素添加量的增加,群落的物候和群落物种组成的变化越显著。
3 讨论
氮素是陆地生态系统植物生长的主要限制性因子,氮素的增加会影响植物的功能性状和植物的繁殖策略[12,30-31],最终降低草地群落物种多样性,改变群落结构[7-8]。植物性状指与植物形态、生理和物候有关的特性,是对植物的生长、繁殖、存活以及适合度具有重要影响的植物特性[32]。本研究中,所有观测物种的花期高度都在氮素添加后增加,禾草和莎草高度的增加最强烈。禾草和莎草的始花期显著推迟,氮素添加使植物对营养部分和繁殖部分投入量也会发生相应的变化[13,32]。从时间的角度来看,植物在生长季节内面临营养生长与生殖生长之间时间分配权衡,生长季节内如果营养生长期延长,则生殖生长期相应推迟[33],结果氮素添加使得禾草和莎草的始花期显著推迟。双子叶杂草类物种始花期变化不一致:具有固氮能力的豆科植物的始花期和高度与对照差异不显著,其余植物除了菊科的两个物种始花期推迟外,其余各科物种的始花期均提前。氮素添加后所有物种的高度和生物量都增加,但禾草类生物量比非禾本科显著增加[34],氮素添加后双子叶类物种花期提前可能是由于双子叶植物的开花时间受内部因素的调控大于其受外界环境变化的影响[11]。本研究中氮素添加显著增加了CWM-H和CWM-SLA,它们高度代表了对光资源的竞争能力[35-36],而比叶面积则代表了资源的投入与产出[37],两个特征反映了群落对资源的竞争能力和生长速率。由于氮素增加后群落中禾草类的生长速率和高度显著高于非禾本科物种[38-39],在群落中双子叶植物的个体相对较小,为了避免与禾草在资源上(主要是光竞争)的竞争,提前开花可以有效地确保繁殖部分的生长,保证其未来在群落中的生存,这可能是氮素添加后双子叶植物比对照群落的始花期提前的一个原因,但是潜在的机理还有待于进一步深入研究。2006年Cleland等[11]研究了模拟氮沉降对一年生草地群落中9个物种花期物候的影响,发现氮素量为7 g·m-2时杂草类花期提前2~4 d,禾草类花期推迟2~6 d,与本研究结果基本一致。

图1 氮素添加处理下科间植物始花期的变化Fig.1 Changes of first flowering day between families among nitrogen addition treatments
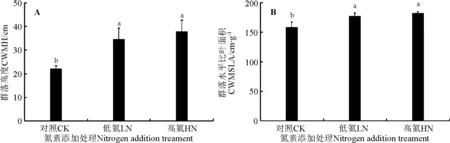
图2 氮素添加对植物群落高度(A)和群落比叶面积(B)的影响Fig.2 Effects of nitrogen addition on CWM-H (A) and CWM-SLA (B)

图3 氮素添加对辛普森多样性指数的影响Fig.3 Effects of nitrogen addition on Simpson’s diversity index

图4 植物开花物候加权的群落结构相似性在氮素添加梯度上的非度量多维尺度分析(NMDS)排序(stress value=0.08) Fig.4 Nonmetric multidimensional scaling [NMDS] ordination of phenologically-weighed community similarity among nitrogen addition gradient(stress value=0.08)
功能特征在群落构建中具有非常重要的作用,花期物候作为一个影响繁殖的关键因素,同样能够影响群落构建。花期物候对环境因素变化反应敏感,花期物候变动对生态系统结构与功能产生深远影响[40]。非度量多维尺度分析(NMDS)排序的结果表明,在不同的氮素添加下群落花期物候和群落结构有显著差异,随着氮素梯度的升高开花物候加权的群落结构相似性变低,群落每次物候观测之间的相似性也变低。总体上,氮素添加后莎草类和禾草类植物始花期推迟,而双子叶类杂草始花期提前,这种反方向的变化使得氮素添加的群落与自然群落在物候和群落结构上产生了分离。氮素添加后群落物候偏移促进非相似性群落形成,施肥后花期物候偏移所导致的群落生态位变化,降低群落物候补偿,增强了功能群间对限制性资源的竞争,群落中出现的生态位空洞为物种入侵或对群落结构变化提供生态位间隙[24]。群落中各物种对环境变化的响应会对物种之间竞争的相互作用产生潜在的影响,群落花期物候偏移潜在影响到更高营养级物种组成,可能会引起植物在繁殖及相互竞争共存上的新格局,最终改变群落的组成[20,27]。
[1] Holland E A,Dentener F J,Braswell B H,etal.Contemporary and pre-industrial global reactive nitrogen budgets[J].Biogeochemistry,1999,46:7-43.
[2] Galloway J N,Dentener F J,Capone D G,etal.Nitrogen cycles:past,present,and future[J].Biogeochemistry,2004,70:153-226.
[3] Galloway J N,Townsend A R,Erisman J W,etal.Transformation of the nitrogen cycle:Recent trends,questions,and potential solutions[J].Science,2008,320:889-892.
[4] Bowman W D,Cleveland C C,Baron J S,etal.Negative impact of nitrogen deposition on soil buffering capacity[J].Nature Geoscience,2008,1:767-770.
[5] Tilman D.Secondary succession and the pattern of plant dominance along experimental nitrogen gradients[J].Ecological Monographs,1987,57:189-214.
[6] Zavaleta E S,Shaw M R,Chiarellio N R,etal.Grassland responses to three years of elevated temperature,CO2,percipitation and N deposition[J].Ecological Monographs,2003,73:585-604.
[7] Stevens C J,Dise N B,Mountford J O.Impact of nitrogen deposition on the species richness of grasslands[J].Science,2004,303:1876-1879.
[8] Clark C M,Tilman D.Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands[J].Nature,2008,451:712-715.
[9] Vitousek P M,Aber J D,Howarth R W,etal.Technical report:human alteration of the global nitrogen cycle:Hources and consequences[J].Ecological Applications,1997,7:737-750.
[10] Diekmann M,Falkengren-Grerup U.Prediction of species response to atmospheric nitrogen deposition by means of ecological measures and life history traits[J].Journal of Ecology,2002,90:108-120.
[11] Cleland E E,Chiariello N R,Laorie S R,etal.Diverse responses of phenology to global changes in a grassland ecosystem[J].PNAS,2006,103:13740-13744.
[12] Obeso J R.Costs of reproduction inIlexaquifolium:Effects at tree,branch and leaf levels[J].Journal of Ecology,1997,85:159-166.
[13] Niu K C,Luo Y J,Philippe C L,etal.The role of biomass allocation strategy in diversity loss due to fertilization[J].Basic and Applied Ecology,2008,9:485-493.
[14] Elzinga J A,Atlan A,Biere A,etal.Time after time:Flowering phenology and biotic interactions[J].Trends in Ecology & Evolution,2007,22:432-439.
[15] Bolmgren K,Cowan P.Time-size tradeoffs:A phylogenetic comparative study of flowering time,plant height and seed mass in a north-temperate flora[J].Oikos,2008,117:424-429.
[16] Walther G R,Post E,Convey P,etal.Ecological responses to recent climate change[J].Nature,2002,416:389-395.
[17] Tooke F,Battey N H.Temperate flowering phenology[J].Journal of Experimental Botany,2010,61:2853-2862.
[18] Weiher E,Van der Werf A,Thompson K,etal.Challenging Theophrastus:A common core list of plant traits for functional ecology[J].Journal of Vegetation Science,1999,10:609-620.
[19] 刘志民,闫巧玲,马君玲,等.生殖物候与草甸草地多年生植物的消长[J].生态学报,2006,26(3):773-779.
[20] Sherry R A,Zhou X H, Gu S L,etal.Divergence of reproductive phenology under climate warming[J].PNAS,2007,104:198-202.
[21] Menzel A.Phenology:Its importance to the global change community[J].Climatic Change,2002,54:379-385.
[22] Menzel A,Fabian P.Growing season extended in Europe[J].Nature,1999,397:659-659.
[23] Fitter A H,Fitter R S R.Rapid changes in flowering time in British plants[J].Science,2002,296:1689-1691.
[24] Cleland E E,Chuine I,Menzel A,etal.Shifting plant phenology in response to global change[J].Trends in Ecology & Evolution,2007,22:357-365.
[25] 巴雅尔塔,贾鹏,杨晓,等.青藏高原高寒草甸组分种花期物候对施肥响应[J].草业学报,2010,19(3):233-239.
[26] Wilson S D,Tilman D.Plant competition and resource availability in response to disturbance and fertilization[J].Ecology,1993,74:599-611.
[27] Martinkova J,Smilaur P,Mihulka S.Phenological pattern of grassland species:Relation to the ecological and morphological traits[J].Flora,2002,197,290-302.
[28] Hovenden M J,Wills K E,Vander Schoor J K,etal.Flowering phenology in a species-rich temperate grassland is sensitive to warming but not elevated CO2[J].New Phytologist,2008,178:815-822.
[29] Garnier E,Cortez J,Billes G,etal.Plant functional markers capture ecosystem properties during secondary succession[J].Ecology,2004,85:2630-2637.
[30] Obeso J R.Costs of reproduction in plant[J].New Physiologist,2002,155:321-348.
[31] Niu K C,Cholar P,Zhao B B,etal.The allometry of reproductive biomass in response to land use in Tibetan alpine grasslands[J].Functional Ecology,2009,23:274-283.
[32] Violle C,Navas M L,Vile D,etal.Let the concept of trait be functional[J].Oikos,2007,116:882-892.
[33] Bloom A J,Chapin F S,Mooney H A,etal.Resource limitation in plants an economic analogy[J].Annual Review of Ecology and Systematics,2003,16:363-392.
[34] Ren Z W,Li Q,Chu C J,etal.Effects of resource additions on species richness and ANPP in an alpine meadow community[J].Journal of Plant Ecology,2010,3:25-31.
[35] Falster D S,Westoby M.Plant height and evolutionary games[J].Trends in Ecology & Evolution,2003,18:337-343.
[36] Moles A T,Warton D I,Warman L,etal.Global patterns in plant height[J].Journal of Ecology, 2009,97:923-932.
[37] Reich P B,Ellsworth D S.Generality of leaf trait relationships:A test across six biomes[J].Ecology,1999,80:1955-1969.
[38] Li W,Wen S J,Du G Z,etal.Root-shoot competition interactions cause diversity loss after fertilization:A field experiment in an alpine meadow on the Tibetan Plateau[J].Journal of Plant Ecology,2011,4:138-146.
[39] 葛庆征,魏斌,江小雷,等.草地恢复措施对高寒草甸植物群落的影响[J].草业科学,2012,29(10):1517-1520.
[40] Smith M M,Knapp A K,Collins S L.A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change[J].Ecology,2009,90:3279-3289.
猜你喜欢
数学物理学报(2022年5期)2022-10-09
热带作物学报(2022年6期)2022-07-08
草地学报(2021年7期)2021-08-03
河北画报(2020年8期)2020-10-27
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
西南农业学报(2017年5期)2017-06-23
河北林业科技(2016年5期)2016-11-08
浙江大学学报(工学版)(2016年2期)2016-06-05
西南农业学报(2016年5期)2016-05-17
