
杜仲EuACOTS基因家族的鉴定及生物信息学分析
2013-04-03 09:15乌云塔娜叶生晶
经济林研究 2013年4期
乌云塔娜,王 淋,叶生晶
(1.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;2.国家林业局杜仲工程技术研究中心,河南 郑州 450003;3.中南林业科技大学a.经济林育种与栽培国家林业局重点实验室;b.林学院, 湖南 长沙410004;4.国家林业局中南林业调查规划设计院,湖南 长沙 410014)
杜仲Eucommia ulmoidesOliv.为杜仲科杜仲属植物,是仅存的孑遗植物[1]。杜仲为落叶乔木,是我国特有的经济树种[2]。杜仲在我国的栽培面积约占世界的90%以上,我国的中亚热带到暖温带地区均有分布,多达16个省、市[3]。 由于杜仲具有药用价值,含有大量的京尼平苷酸、绿原酸等,具有镇痛、保肝等功效[4],而且杜仲的树皮、果实以及叶子中均有含有杜仲胶,可代替天然橡胶,具有很高的商业价值[5-6]。因此杜仲被称为世界上适应范围最广的重要胶源树种[7-8],杜仲胶被誉为基因橡塑二重性的高分子合金材料[9]。
杜仲天然橡胶(杜仲胶)又名古塔波胶(Gutta percha),其化学结构为反式聚异戊二烯,而普通天然橡胶巴西橡胶为顺式聚异戊二烯,两者互为同分异构体[10]。杜仲胶是一种多萜类化合物,萜类物质(Terpenoids)是植物中重要的次生代谢产物,1958年,Lynen等[11-12]提出了萜类物质合成是主要由位于细胞质的甲羟戊酸(Mevalonate,MVA)途径完成。随后日本学者用同位素标定法标记萜类物质合成MVA 途径代谢的中间产物发现MVA代谢途径存在于杜仲的含胶细胞中[13]。因此,目前推测杜仲胶有可能在细胞质途径中发生。
乙酰COA酰基转移酶(acetoacetylCoA thiolase,ACOT)是萜类物质合成MVA途径起始酶[10],对众多蛋白的酰基化及去酰基化起催化作用[14-15]。在MVA途径中,2个乙酰CoA分子(acetylCoA)在乙酰乙酰CoA硫解酶(ACOT)作用下,合成乙酰乙酰CoA (acetoacetylCoA),是合成萜类物质必须前体异戊烯焦磷酸(IsopentenylPP, IPP)的第一步[16-17]。
文中通过杜仲转录组以及基因组序列得到了杜仲ACOTS基因家族序列,对其进行了生物信息学分析,从植物功能基因转录水平的角度探讨了EuACOTS基因家组的调控机制,旨在为生物合成杜仲胶方面的研究奠定理论基础。
1 材料与方法
1.1 数据来源
对杜仲胶合成时期转录组数据组装及基因功能注释和杜仲全基因组测序数据为依据进行序列相关生物信息学分析[18-19]。
1.2 方 法
利用 ExPASyProtParam(http: / /cn.expasy.org)程序与ScanProsite程序对EuACOTS家族基因氨基酸残基数、氨基酸组成、蛋白质相对分子量、理论等电点、蛋白质的亲疏水性等性质以及蛋白的结果保守域进行了预测与分析;利用TMHMM程序 (http://www.cbs.dtu.dk/services/TMHMM2.0/)预测蛋白质跨膜结构域;利用PSIPRED程序(http: / /bioinf.cs.ucl.ac.uk /psipred)预测蛋白质的二级结构;利用Swiss model (http://swissmodel.expasy.org/)程序,根据基因氨基酸序列进行建模,预测蛋白质的三级结构;利用在线软件MEME(http://meme.nbcr.net/meme/)预测蛋白的结构保守域;利用软件GSDS(http://gsds.cbi.pku.edu.cn/index.php)对基因的内含子和外显子进行了分析;利用PLACE(http://www.dna.affrc.go.jp/PLACE/signalscan.html)和 PLANT CARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线软件预测EuACOTS家族启动子序列中可能存在的顺式元件进行分析。
2 结果与分析
2.1 ACOTS基因家族序列分析
杜 仲EuACOTS(EuACOT8、EuACOT9、EuACOT10)基因家族氨基酸序列与拟南芥Arabidopsis thaliana、 葡 萄Viteus vitifoliae的ACOT基因家族的基因序列相似性达到80%以上(见图1),因此命名为EuACOTS,EuACOTS基因在杜仲中至少包含3个家族成员EuACOT8、EuACOT9、EuACOT10[20]。通过Prosite数据库(http://prosite.expasy.org/)分析EuACOTS蛋白质的结构位点,结果表明,EuACOT8的第354~370氨基酸残基、EuACOT9的350~366氨基酸残基、EuACOT10的354~370氨基酸残基均存在保守序列 NvhGGaVSlGHPlGcSG;EuACOT8的 第 389~402氨基酸残基、EuACOT9的385~398氨基酸残基、EuACOT10的389~402氨基酸残基存在保守序列GVGGICNGgGgAsA,2个保守序列均为硫解酶特异序列(见图1),EuACOTS属于硫解酶。
2.2 ACOTS基因家族聚类分析
利用MEGA软件的邻结法(neighbor joining, NJ),构建了杜仲ACOTS的系统进化树(见图2)。选择单子叶植物玉米Zea mays、双子叶植物拟南芥Arabidopsis thaliana、葡萄Viteus vitifoliae、苔藓类小立碗藓Physcomitrella patens的ACOT基因家族进行进化分析。从进化树中可见,共分为3个主要的分支,其中EuACOTS位于1个分支,且于双子叶植物拟南芥和葡萄的遗传距离较近,与单子叶植物玉米以及藓类遗传距离较远,这与传统的分类结果相一致。

图1 全长ACOTS蛋白多序列对比Fig.1 Multiple sequence alignment of the full length ACOTS proteins
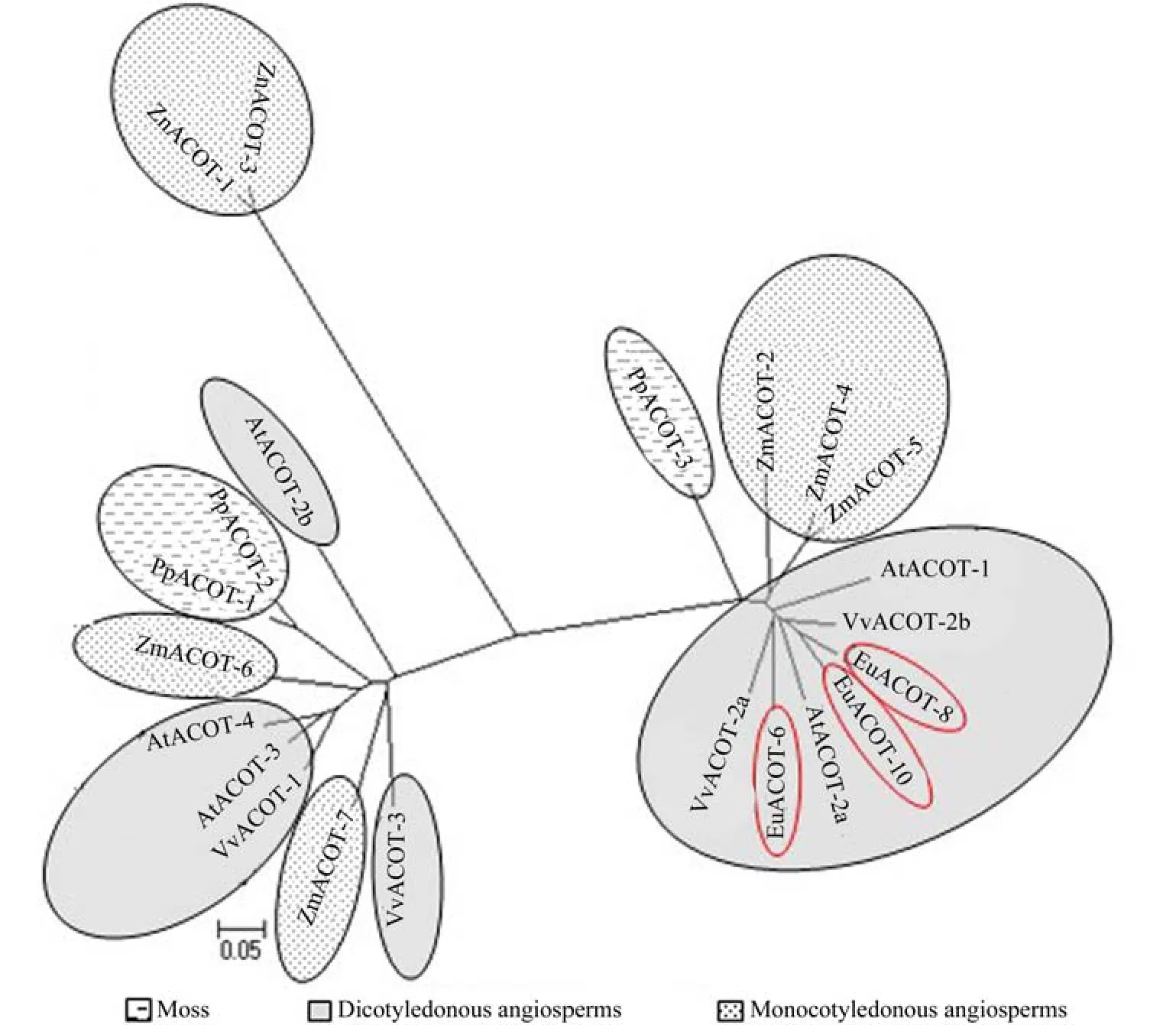
图2 ACOTS蛋白的聚类分析Fig.2 Cluster analysis of ACOTS proteins
2.3 ACOTS基因家族的蛋白质理化性质分析
利用生物信息学在线软件,对ACOTS基因家族的蛋白质理化性质进行预测,如表1所示。EuACOTS编码氨基酸分别为406 aa、314 aa、406 aa;理论等电点分别为 5.66、8.92、5.80;EuACOTS均属于稳定的疏水性蛋白,与其亲缘关系较为相近的VvACOT2a、VvACOT2b以及AtACOT2a属于稳定的疏水性蛋白相一致。
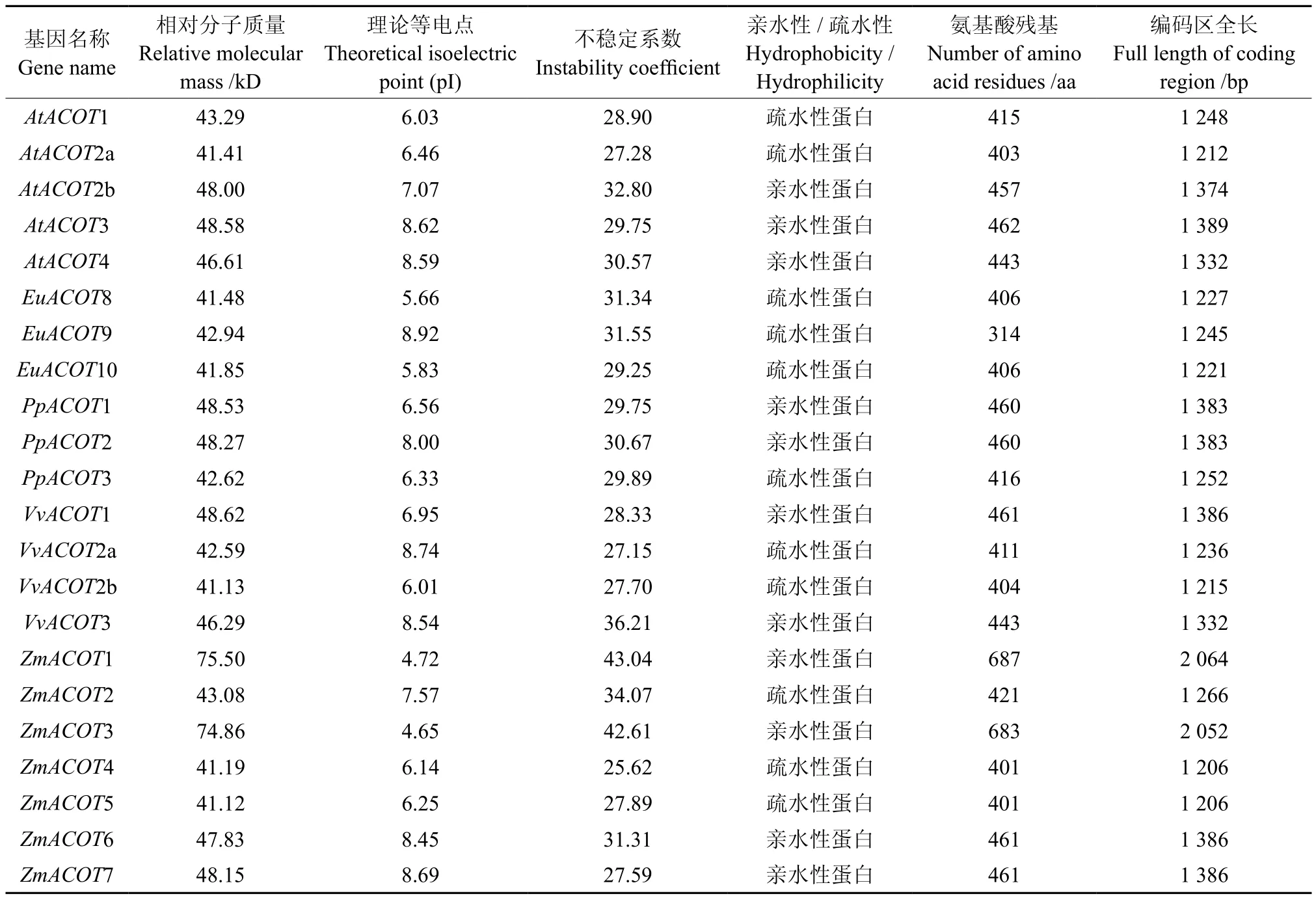
表1 EuACOTS编码氨基酸的基本理化参数Table 1 Some basis physical and chemical parameters of EuACOTS
2.4 EuACOTS基因蛋白结构保守域预测
利用MEME在线软件预测EuACOTS基因家族蛋白结构保守域(见图3)。在ACOTS基因家族中,除了VvACOT2b、ZmACOT1、ZmACOT3、ZmACOT7之外,其它基因均含有3个结构保守域,且其分布具有一定的规律性,第2个和第3个保守域均为硫解酶特异保守序列。
2.5 EuACOTS基因家族的蛋白质基因跨膜结构域的预测
利用在线工具TMHMM2.0对EuACOTS家族氨基酸序列的跨膜结构域进行预测(见图4)。EuACOTS家族蛋白无明显跨膜区,蛋白全部在膜外。
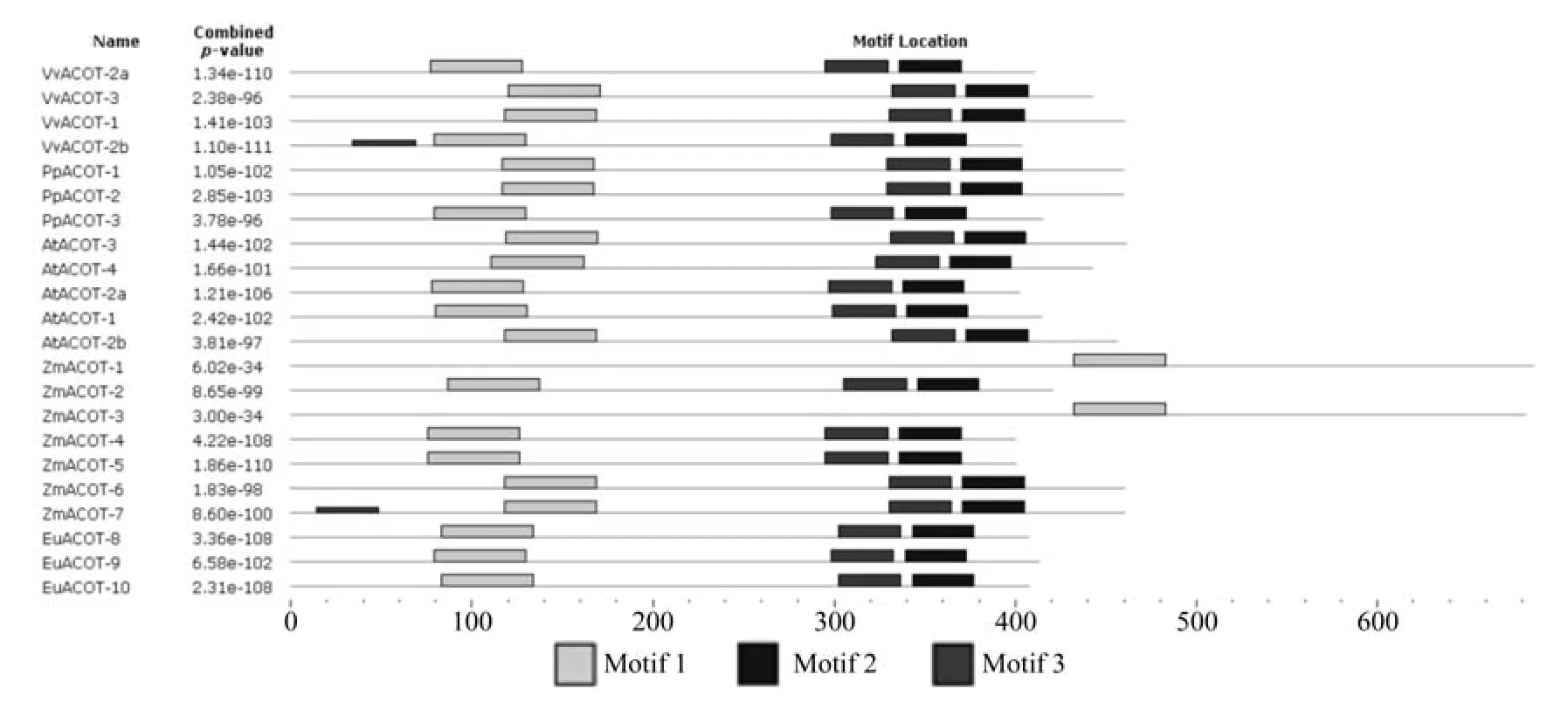
图3 ACOTS基因的保守结构域Fig.3 Some conserved domains in ACOTS gene

图4 EuACOTS蛋白的跨膜区预测Fig.4 Predicted transmembrane region of the deduced EuACOTS protein
2.6 EuACOTS基因家族蛋白的二级结构的预测
根据EuACOTS家族的氨基酸序列,预测家族蛋白质二级结构(见图5)。其中EuACOT8蛋白二级结构中α螺旋占40.69%,β折叠占15.44%,螺环结构占43.87%,属于混合型结构的蛋白质;EuACOT9蛋白二级结构中α螺旋占41.55%,β折叠占14.01%,螺环结构占44.44%,属于混合型结构的蛋白质;EuACOT10蛋白二级结构中α螺旋占39.95%,β折叠占14.46%,螺环结构占45.59%,属于混合型结构的蛋白质。

图5 EuACOTS蛋白二级结构预测Fig.5 Predicted secondary structure of the deduced EuACOTS protein
2.7 EuACOTS基因家族蛋白的三级结构的预测
采用auto mode建模方式以蛋白模板(2ib8D)对EuACOTS蛋白进行同源建模,得到EuACOTS蛋白的三维空间模型(见图6)。EuACOT8蛋白模型QMEAN 6 得分为0.6,蛋白序列的相似性为49.62%;EuACOT9蛋白模型QMEAN 6 得分为0.58,蛋白序列的相似性为48.09%;EuACOT10蛋白模型QMEAN 6得分为0.57,蛋白序列的相似性为48.23%。EuACOTS家族基因的蛋白质三级结构极为相似。

图6 EuACOTS蛋白三级结构预测Fig.6 Predicted tertiary structure of the deduced EuACOTS protein
2.8 EuACOTS基因的内含子和外显子预测及分析
利用在线软件GSDS,将杜仲ACOT基因组序列及其对应的完整编码序列(coding sequence CDS)提交到GSDS网站系统,获得了EuACOTS基因结构示意图以及外显子、内含子分布图(见图7、表2)。EuACOTS基因家族基因组序列含有相同的外显子和内含子数量,均为13个外显子,12个内含子,其中内含子/外显子的边界均符合GTAG 剪切模式,EuACOTS其中7个内含子位于0相位,3个内含子位于1相位,2个内含子位于2相位。
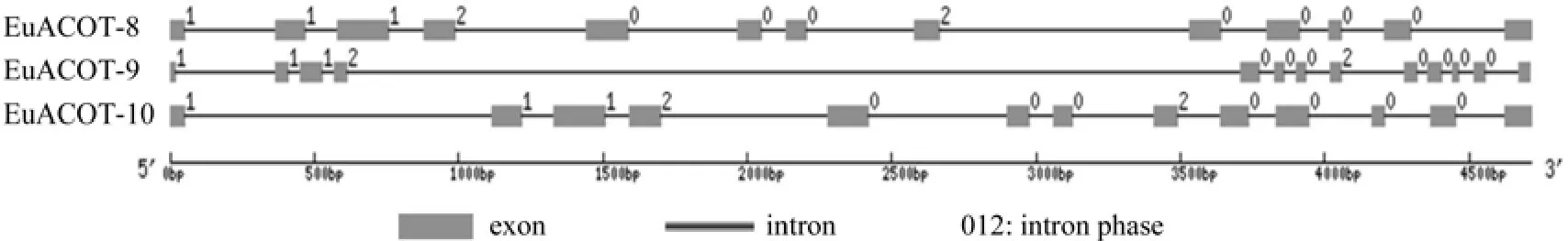
图7 EuACOTS基因内含子和外显子预测Fig.7 Prediction of the introns and exons in EuACOTS
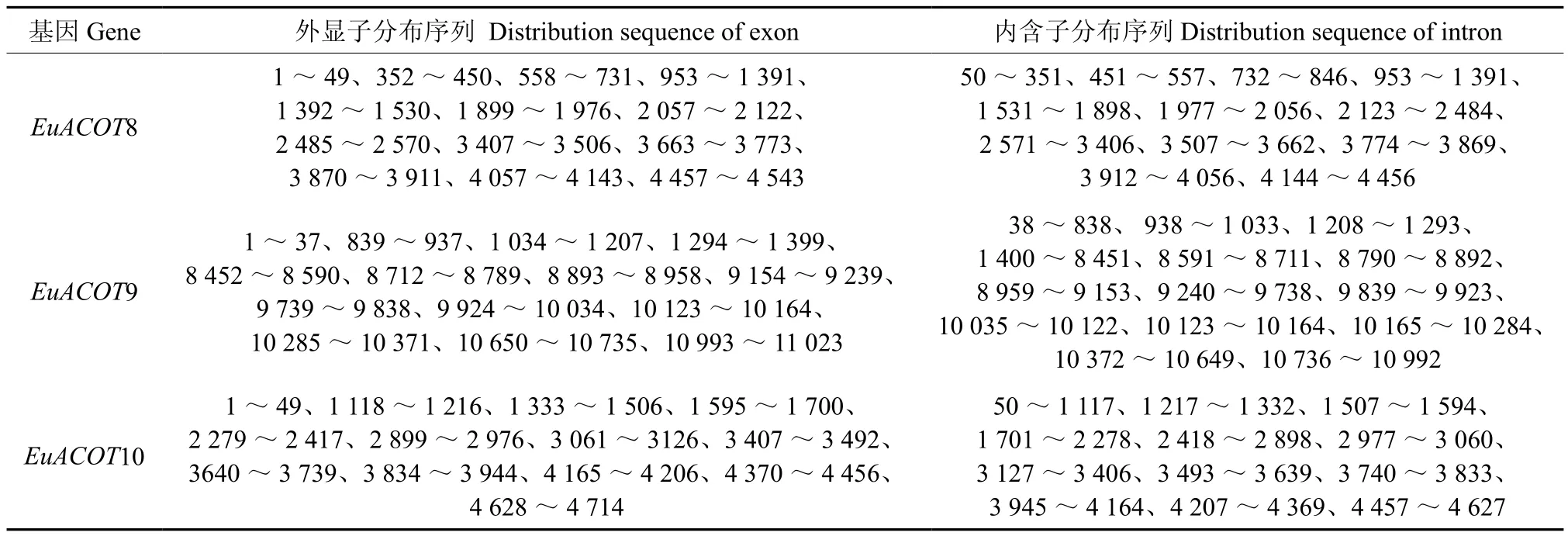
表2 EuACOTS外显子和内含子分布序列Table 2 Distribution sequence of the introns and exons in EuACOTS bp
2.9 EuACOTS基因家族启动子的顺式元件作用分析
根据杜仲基因组测序结果,得出EuACOTS家族基因组序列,应用PLANCE以及PLANT CARE软件分析其家族启动子序列(5′UTR 2 000 bp)主要顺式元件潜在的分布以及功能。EuACOTS家族基因启动子主要顺式元件潜在的分布以及功能见表3。EuACOTS家族启动子含有大量基本顺式作用元件TATAbox、CAATbox,除此之外还存在光、乙烯、真菌诱导和脱落酸响应元件等多个顺式调控元件。其中EuACOT8特有赤霉素和热胁迫响应顺式作用元件,EuACOT8可能参与赤霉素诱导以及热胁迫响应基因的调控;EuACOT9特有茉莉酸响应元件,EuACOT9可能受茉莉酸热胁迫响应基因的调控;而EuACOT10特有生长素响应元件,可能参与生长素诱导响应基因的调控。
3 结论与讨论
乙酰CoA酰基转移酶(AOCT)属于硫解酶,能催化蛋白质的酰基化和去酰基化,调节蛋白质活性、基因表达等功能[15,21]。ACOT基因已经从大多数植物中克隆并进行了相关分析,但目前对于杜仲ACOT基因家族的研究并未见报道,文中主要对杜仲ACOT基因家族进行了生物信息学分析,EuACTOS基因家族编码氨基酸为314~406 aa,EuACTOS基因家族编码氨基酸与其它植物的ACOT基因具有较高的同源性,与其遗传距离较近的为双子叶植物葡萄(VvACOT2a、VvACOT2b)以及拟南芥(AtACOT2a),这些基因在结构及特性上相符合,表明EuACTOS基因家族是双子叶植物ACOT基因,属于萜类物质合成MVA途径的起始酶。Iván Ahumada[22]提出拟南芥中有2种不同的乙酰基转移酶AtACOT1和AtACOT2,对其进行亚细胞定位得知AtACOT1主要定位过氧化物酶中,而AtACOT2主要定位在和细胞质和细胞核中,且AtACOT2的在拟南芥中的相对表达量较高,得出AtACOT2为萜类物质合成关键酶,对异戊二烯的生物合成有关键作用。聚类分析结果表明,EuACTOS基因家族的基因与AtACOT2a及AtACOT2b的遗传距离较近,更加说明EuACTOS基因家族应为萜类物质合成的关键酶,对杜仲胶的合成应有一定的作用。
通过生物信息学方法分别从核苷酸和蛋白质序列水平上,分析了EuACTOS基因家族的序列特征,发现EuACTO基因编码蛋白均属于稳定的疏
水蛋白质,其二级结构均为螺旋和折叠结构并存的混合结构蛋白质,均含外显子13个内含子12个,启动子序列部分含有大量的基本顺式作用元件TATAbox,CAATbox以及光、乙烯、真菌诱导和脱落酸响应元件等多个顺式调控元件。
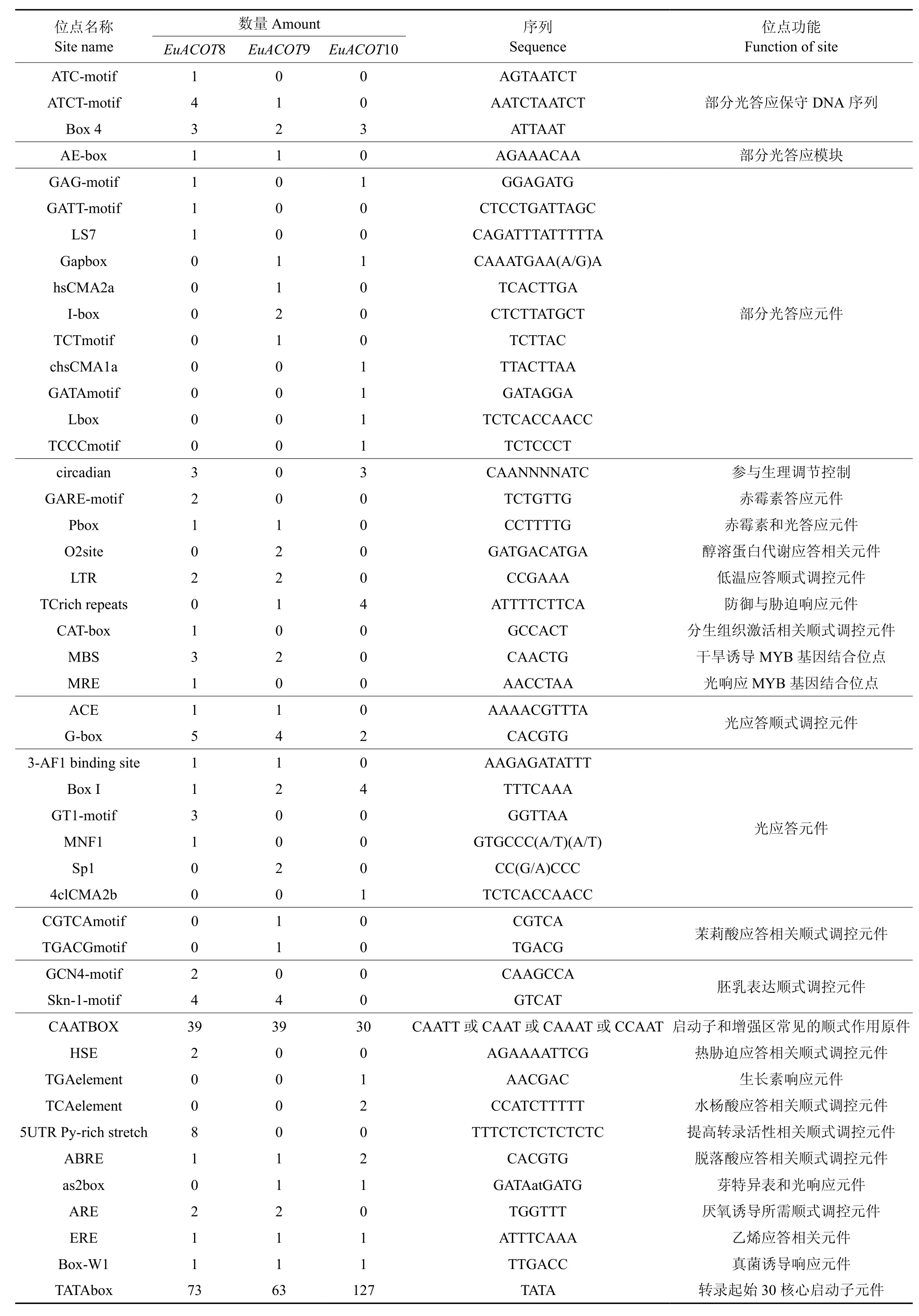
表3 EuACOTS基因启动子区顺式元件作用预测Table 3 Function prediction of the cis-elements of promoter in EuACOTS
参考文献:
[1] 高 超,闫文德,田大伦,等.杜仲光合速率日变化及其与环境因子的关系[J].中南林业科技大学学报,2011, 31(5):100-104.
[2] 胡宋其.杜仲丰产栽培技术[J].农业科技,2006, 27:151.
[3] 何 方,张康健,王承南,等.杜仲产区划分[J].经济林研究,2010, 28 (2):15-17.
[4] 杜红岩,刘昌勇,李 钦,等.杜仲叶中3种主要活性成分含量的季节变化[J].中南林业科技大学学报,2011, 31(8): 6-9.
[5] 卢 敏,胡正海,田兰馨.杜仲茎韧皮部超微结构的初步研究[J].浙江林学院学报, 1990,7(4):316-321.
[6] 杜红岩,杜兰英,谢碧霞,等.杜仲叶内杜仲胶的形成积累规律[J].中南林学院学报, 2006, 26(2):1-6.
[7] 杜红岩,胡文臻,俞 锐.杜仲产业绿皮书:中国杜仲橡胶资源与产业发展报告[M].北京:社会科学文献出版社,2013:2.
[8] 杜红岩,赵 戈,卢绪奎.论我国杜仲产业化与培育技术的发展[J].林业科学研究, 2000, 13 (5) : 554-561.
[9] 杜红岩.我国的杜仲胶资源及其开发潜力与产业发展思路[J].经济林研究,2010,28(3):1-4.
[10] 严瑞芳.杜仲胶研究进展及发展前景[J].化学进展,1995,1(7): 65-71.
[11] Chaykin S, Law J, Phillips A H,et al.Phosphorylated Intermediates in the Synthesis of Squalene[J].Proc N A S, 1958,44 (10):998-1004.
[12] Lynen F, Efferer H, Henning U,et al.Farnesyl-pyrophosphatund 3-Methyl-Δ3-butenyl-1-pyrophosphat, die biologischen Vorstufendes Squalens.Zur Biosynthese der Terpene, III [J].Angewandte Chemie, 1958 ,70(24):738-742.
[13] Takeshi B, Michiko M, Koichirou G,et al.Contribution of mevalonate and methylerythritol phosphate pathways to polyisoprenoid biosynthesis in the rubberproducing plantEucommia ulmoidesOliver[J].Z Naturforsch C, 2010, 65(5/6):363-372.
[14] Lichtenthaler HK.The 1-deoxydxylulose-5-phosphate (DOXP)pathway for the of isoprenoid biosynthesis in plants[J].Annu RevPlant Physiol Plant MolBiol, 1999, 50: 47-65.
[15] 张 琳,谭晓风,胡 娇,等.油茶乙酰CoA酰基转移酶基因cDNA克隆及序列特征分析[J].中南林业科技大学学报,2011, 31(8):108-112 .
[16] 王计平,史华凭,李润植.植物种子油合成的调控与遗传修饰[J].植物遗传资源学报, 2007, 7(4):488-493.
[17] 兰闻智,余龙江,蔡永军,等.类异戊二烯非甲羟戊酸代谢途径的分子生物学研究进展[J].西北植物学报, 2001,21(5):1039-1047.
[18] 李铁柱,杜红岩,刘慧敏,等.杜仲幼果和成熟果实转录组数据组装及基因功能注释[J].中南林业科技大学学报, 2012,32(10):9-17.
[19] 李铁柱,杜红岩,刘慧敏,等.杜仲果实和叶片转录组数据组装及基因功能注释[J].中南林业科技大学学报, 2012,32(11):122-130.
[20] 叶生晶.杜仲MVA途径相关基因表达差异及全长cDNA序列特征[D].长沙:中南林业科技大学,2012.
[21] 崔光红,王学勇,冯 华,等.丹参乙酰CoA酰基转移酶基因全长克隆和SNP分析[J].药学学报, 2012, 45(6):785-790.
[22] Iván Ahumada, Albert Cairó, Andréa Hemmerlin.Characterisation of the gene family encoding acetoacetyl CoA thiolase inArabidopsis[J].Functional Plant Biology, 35(11):1100-1111.
猜你喜欢
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
小哥白尼(军事科学)(2019年9期)2019-12-21
生物工程学报(2019年6期)2019-07-10
电影(2019年3期)2019-04-04
生物学通报(2019年1期)2019-02-15
生物学通报(2018年12期)2018-10-10
中成药(2018年8期)2018-08-29
少儿科学周刊·少年版(2017年3期)2017-06-29
陕西画报(2016年1期)2016-12-01
小雪花·小学生快乐作文(2015年9期)2015-09-29
