
三沙湾浮游动物生态类群演替特征
2013-09-11 08:38徐佳奕徐兆礼
生态学报 2013年5期
徐佳奕 ,徐兆礼
(1.厦门大学海洋学系,厦门 361005;2.中国水产科学研究院东海水产研究所,农业部海洋与河口渔业重点开放实验室,上海 200090)
三沙湾位于福建省东北部沿海,具有典型的亚热带气候特征,四季交替明显。每年的10月至翌年4月盛行东北季风,6月至8月盛行西南季风,5月和9月为转风期[1]。冬季海域受随着东北季风流南下的浙闽沿岸流影响;夏季,西南季风流携带南海高温水团以及台湾暖流的共同作用,高温高盐水侵入湾内[2]。根据福建省志(气象志)的描述,5—6月为每年的雨季,大量大陆径流淡水注入湾内[3],并伴随转风期南北水团的共同作用,使得海区水文环境较为复杂。因此,三沙湾海洋环境因子也具有明显的季节交替。三沙湾浮游动物季节变化对于反映我国亚热带海湾浮游动物的一般特征具有一定的代表性。
我国对近海、河口和海湾等的浮游动物研究已有许多报道[4-7],但关于浮游动物生物群落生态类群组成结构时空变化特征及其成因的专题研究尚未见报道。对三沙湾浮游动物的研究虽有较多报道,如林景宏等[8]列举了浮游动物的3种生态类群,并对海区中浮游动物生物量、种类数分布等做了详尽描述。另外,陈国斌等[9]也有过类似研究。但这些论著所涉生态类群的研究也仅限于对生物量、种类组成等的描述,而并未具体阐述生态类群转变与季节交替之间的重要联系。
鉴于浮游动物的最适温度、盐度等生物学属性,在海水温度、盐度和海流季节变化的影响和胁迫下,导致群落结构特征发生变动。本文通过对浮游动物生态类群组成结构特征季节变化的分析,以期揭示浮游动物群落结构变化与季节交替变化的生态关系。
1 调查海域与方法
2010 年6 月、8 月、10 月和2011 年4 月、5 月,在三沙湾海域(26°25'N—26°48'N、119°38'E—120°07'E)进行5个航次的海洋综合调查,共布设10个站位,见图1。
浮游动物样品采集和室内分析均按照《海洋调查规范》规定进行。用浅水Ⅰ型浮游生物网(口径50 cm、筛绢CQ14、孔径0.505 mm),自海底至水面垂直拖曳获得浮游动物样品,采样站位平均水深42 m。网采样品经5%福尔马林溶液固定,在室内,剔除样品杂质后,进行称重、分类鉴定和计数。
参照相关文献[10],将浮游动物生态类型中温度界限定为热带种(≥25℃)、亚热带种(20—25℃)和暖温带种(15—20℃)。盐度界限定为近海种(28—32)、外海种(32—34)和大洋种(>34)。另充分考虑种类分布的地理和季节特征,确定物种的生态类型[11-19]。
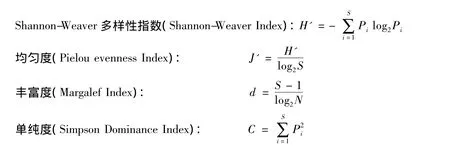
式中,Pi为第i种的丰度与该样方总丰度之比值,S为种类数,N为样方总丰度。
物种更替率:
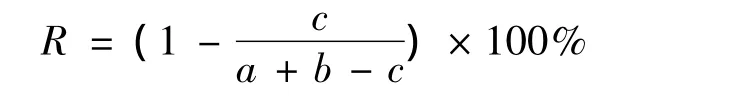
式中,a、b分别为相邻月份的物种数,c为相邻月份共同的物种数[20]。
运用PRIMER 5.0软件,以各月份不同生态类群浮游动物“种类数×丰度”矩阵计算Bray-Curtis距离,建立“月份×月份”距离系数Q型矩阵,采用组平均法作出聚类谱系图,进行聚类分析。

图1 采样站位图Fig.1 Sampling stations in Sansha Bay
2 结果
2.1 浮游动物种类组成
三沙湾共鉴定浮游动物71种(不包含28种浮游幼体),包括腔肠动物门9种,栉水母门2种,节肢动物门52种,软体动物门1种,毛颚动物门4种,尾索动物门3种。其中4月出现17种,5月19种,6月45种,8月43种,10月33种(表1)。
种类各季节的物种更替率R值统计分析结果:R值在4—5月为56.00%,5—6月最高69.39%,6—8月为55.74%,8—10月为50.98%。其中5—6月变动率明显最大,表明5月和6月间的种类交替最明显,8—10月R值最小,表明8月和10月间的种类交替率相对较低。

表1 浮游动物生态类群及其丰度Table1 Abundance and ecological groups of zooplankton species

续表
2.2 生态类群组成分析
三沙湾海域浮游动物主要的几个生态类群组成见表1。
暖温带近海种:主要包括中华哲水蚤(Calanus sinicus)、近缘大眼水蚤(Corycaeus affinis)、五角水母(Muggiaea atlantica)和纳嘎带箭虫(Zonosagitta nagae)等。为三沙湾4月最常见的种类,另外,在5月亦有较多种类出现,而6月、8月和10月,暖温带近海种所占比例逐月下降。
亚热带近海种:如背针胸刺水蚤(Centropages dorsispinatus)、太平洋纺锤水蚤(Acartia pacifica)、真刺唇角水蚤(Labidocera euchaeta)、球型侧腕水母(Pleurobrachia globosa)、百陶带箭虫(Z.bedoti)和中华假磷虾(Pseudeuphausia sinica)等。该种生态类群在除4月外的其他4个月都有较多出现,证明三沙湾海域属于较典型的亚热带海湾。其中6月种类数最多,全部5个月均为第二大生态类群。
亚热带外海种:包括驼背隆哲水蚤(Acrocalanus gibber)、精致真刺水蚤(Euchaeta concinna)和微刺哲水蚤(Canthocalanus pauper)等。4、5月几乎没有出现,但在6、8和10月丰度明显上升,尤其是8月和10月,在研究海区中占很高比例,远远超过暖温带近海种和亚热带近海种的种类数。
另外,三沙湾海域8月还有半口壮丽水母(Aglaura hemistoma)1种热带大洋种出现,因其丰度占总丰度比例极小,下文略去讨论。
综上所述,本次调查中浮游动物生态类群以亚热带外海种和亚热带近海种居多,反映出三沙湾海域的亚热带属性。
2.3 生态类群种类组成及其月变化
从各月生态类群种类变化(表2,图2)可以看出,4月总种类数最少,仅17种。其中以暖温带近海种最多,为13种,占4月总种数76.47%的绝对优势。亚热带近海种为3种和亚热带外海种仅为1种分别占该月总种数百分比的17.65%和5.88%。5月,暖温带近海种8种占42.11%,比例明显下降,亚热带近海种7种占31.58%,开始上升,亚热带外海种数4种也有所增加。6月总种类数共出现45种,达到最大。亚热带外海种升至22种占48.89%,迅速上升并位居第一。亚热带近海种数17种占37.78%,也明显上升。暖温带近海种所占比例13.33%继续降低。8月和10月,亚热带外海种分别为24种和19种,比例为57.14%和57.58%较6月进一步升高,为海区种类组成优势类群。同时,亚热带近海种比例分别为33.33%和39.39%也较高,而暖温带近海种仅有9.52%和3.03%进一步下降。
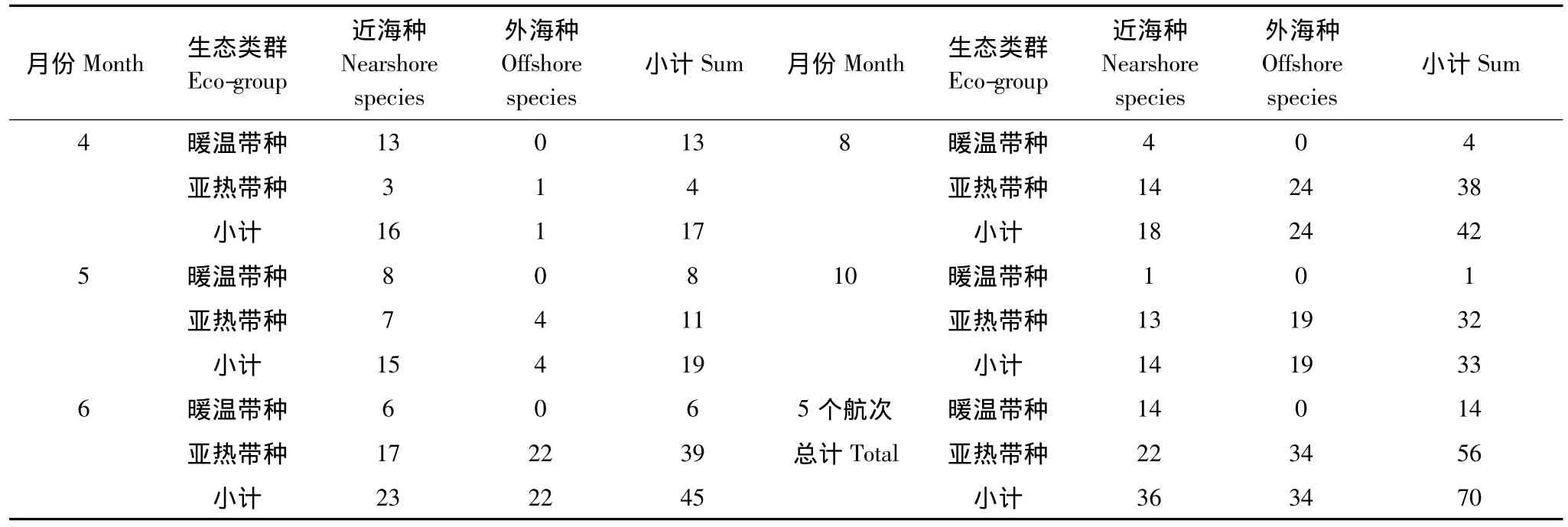
表2 各月份不同生态类群种类数Table 2 The seasonal distribution of species numbers of each eco-group
综上,4月以暖温带近海种占优势,5—8月随温度上升,亚热带近海种和亚热带外海种上升,至8月(全年月均温最高)达到高峰,10月温度回落,亚热带种也随之回落。
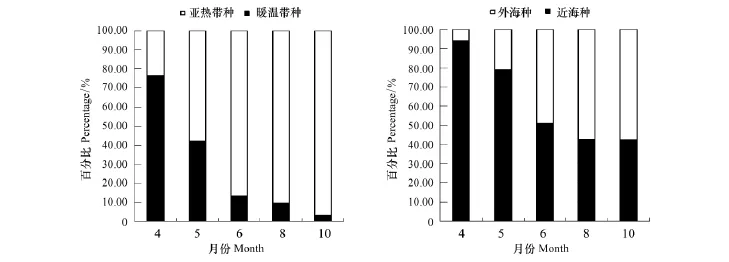
图2 浮游动物各月份不同生态类群种类数的变化Fig.2 The variation of each eco-group species numbers of zooplankton in different seasons
2.4 不同生态类群生物丰度组成及月变化
从表3,图3可看出,4月暖温带近海种丰度为22.52个/m3,占该月浮游动物总丰度的96.28%,其中贡献最大的种为中华哲水蚤,丰度达到15.21个/m3。5月,浮游动物总丰度为调查期间的最低值,仅7.83个/m3,其中最主要的生态类群为亚热带近海种和暖温带近海种,丰度分别达3.86个/m3和3.78个/m3,两者占总丰度的比例较接近,分别为49.30%和48.28%。6月,浮游动物丰度最高的生态类群为亚热带近海种11.56个/m3,占总丰度的73.03%;暖温带近海种和亚热带外海种丰度分别为1.56个/m3和2.71个/m3,仅分别占总丰度的9.85%和17.12%,较低;而亚热带外海种比例为17.12%,较4、5月的1.54%和2.43%有明显上升。8月,浮游动物总丰度出现最高值,达182.23个/m3,主要生态类群为亚热带外海种和亚热带近海种,丰度分别为109.22个/m3和70.84个/m3,分别占总丰度的59.94%和38.87%,而暖温带近海种丰度仅1.19%迅速下降。10月,各生态类群丰度又转为亚热带近海种最高为7.62个/m3,亚热带外海种略低为6.78个/m3,分别占总丰度的52.55%和46.76%,暖温带近海种丰度比例0.69%进一步下降。
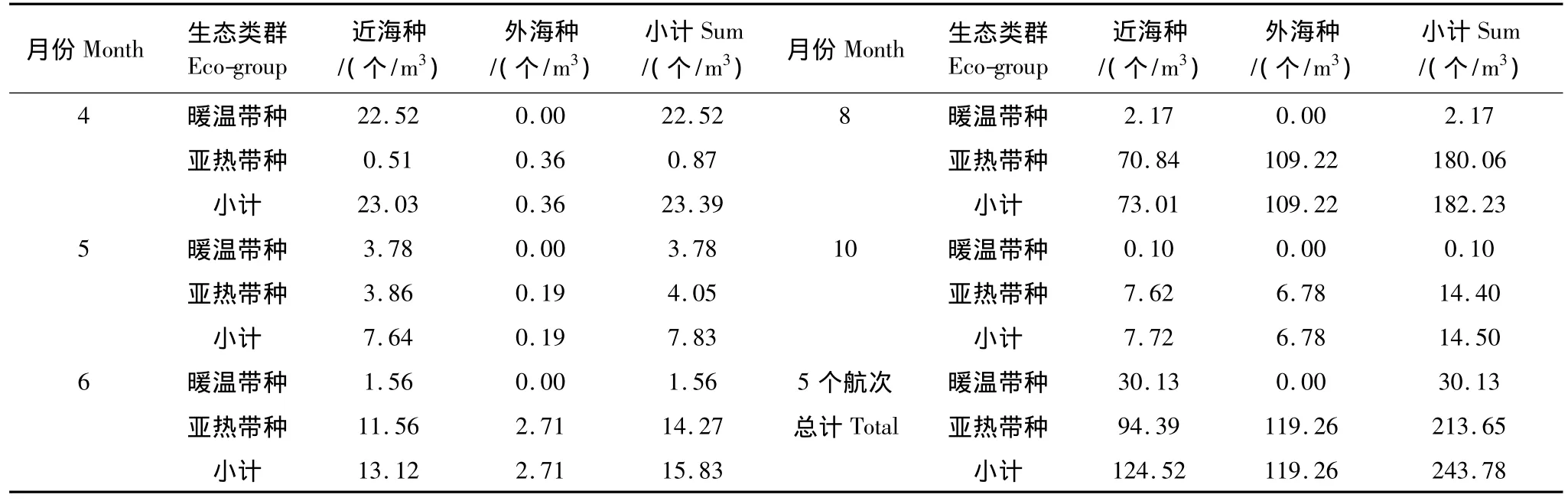
表3 各月份不同生态类群丰度Table 3 The seasonal distribution of abundance of each eco-group
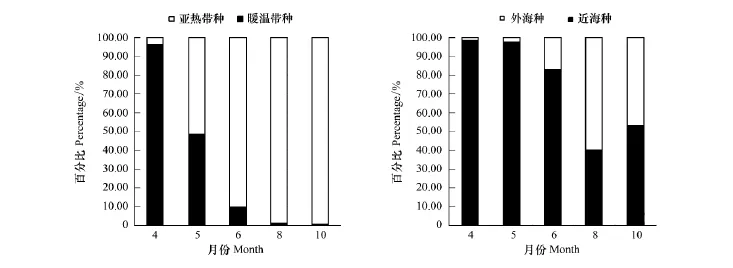
图3 浮游动物各月份不同生态类群丰度的变化Fig.3 The variation of each eco-group abundance of zooplankton in different seasons
2.5 各月份群落生态类群种类及其数量组成的聚类分析
从图4可看出,在 14.1距离系数水平上,6、8、10月聚为A组,表明这3个月份群落生态类群种类和丰度组成情况较相似,其中8月和10月以更小的Bray-Curtis距离聚合,表明其生态类群结构更相似;在21.5距离系数水平上,4、5月聚合为B组,表明这两个月的群落生态类群种类和丰度组成较相似;A、B两组在较大的距离系数上再聚合,表明两组相似性明显较低。
结果可将5个月分为3类。其中4月、5月为Ⅰ类,6月单独为Ⅱ类,8月、10月归为Ⅲ类。
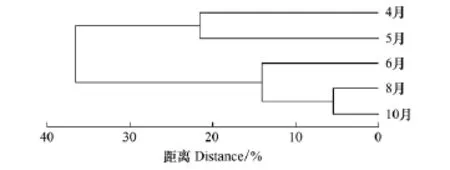
图4 月份聚类谱系树状图Fig.4 The result of cluster analysis
Ⅰ类:以暖温带近海种为最重要生态类群,4月在种类数和丰度比例上均占绝对优势。5月亚热带近海种比例开始上升,但在种类数比例上仍以暖温带近海种占优势。
Ⅱ类:6月为生态类群组成结构的过渡时期,暖温带近海种重要性迅速下降。在丰度百分比上亚热带近海种仍具有较明显优势,而在种类数上亚热带外海种比例已跃居第一。
Ⅲ类:暖温带近海种几乎消失,主要为亚热带外海种和亚热带近海种。8月又以亚热带外海种种类数和丰度所占比例最高,10月亚热带外海种虽在丰度上较亚热带近海种低,但种类数仍居第一高位。
另外,在较高距离水平上B组以亚热带种为主,与A组以暖温带种为主的特征存在明显差异。且在总种类数上,B组显著高于A组。
浮游动物物种更替率分析表明,浮游动物种类组成由4月、5月(Ⅰ类)向6月(Ⅱ类)和8、10月(Ⅲ类)过渡时物种更替率较高,表明Ⅰ类和Ⅱ、Ⅲ类间差异较大。调查同期海水平均温度变化也有类似趋势:4月(15.58℃)和5月(21.78℃)海水温度较低,属于暖温带种适温范围;6月(24.56℃)海水温度升高,较适宜亚热带种生长;8月(27.72℃)和10月(27.42℃)海水温度进一步升高并保持在27℃以上,使得偏热带海域的亚热带种能够大量出现聚集。
2.6 生物多样性分析
由图5可知,6月调查海域生物多样性指数H'和丰富度d平均值分别为3.12和4.52,最高,单纯度C较低。由此推测,经5月丰水期后,霍童溪、白马河等大陆径流大量注入三沙湾,同时台湾暖流向湾内侵入,致使海域温盐梯度增大,不同生态类群浮游动物均有分布,故而6月调查海域生物多样性全年最高,各站位间变化幅度最大。8月生物多样性指数平均值为3.09略低于6月;4、5月生物多样性指数平均值分别为2.03和2.02相近且最低。
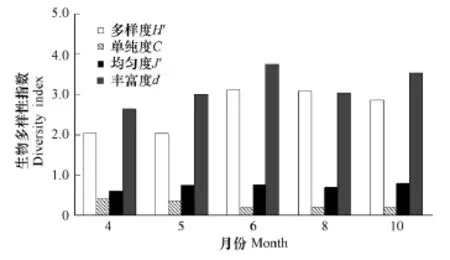
图5 三沙湾浮游动物的多样性指数Fig.5 Diversity index of zooplankton in Sansha Bay
3 讨论
3.1 生态类群种类组成结构的季节交替特征
三沙湾浮游动物种类组成的季节交替变化明显。尤其是5—6月物种更替率最高。
从物种更替率分析来看,4、5月之间的浮游动物种类组成季节交替较不明显,物种更替率为56.00%。从优势种看(表1),4月和5月,中华哲水蚤和纳嘎带箭虫均为三沙湾浮游动物的优势种。其中,中华哲水蚤广泛分布于我国北方海区,在黄海是冬春秋三季的最常见桡足类,在渤海,夏季中华哲水蚤也是主要的优势种[21],为西北太平洋典型的暖温带近海种[22]。另外一些我国暖温带近海的优势种,如:小拟哲水蚤(Paracalanus parvus)、平滑真刺水蚤(E.plana)和拟长腹剑水蚤(Oithona similis)等也是这一时期三沙湾海域常见种。这一时期,三沙湾海域浮游动物种类组成与长江以北我国暖温带一些海湾浮游动物组成相似。例如,在大连湾,浮游桡足类全年以中华哲水蚤、小拟哲水蚤、拟长腹剑水蚤和短角长腹剑水蚤(O.brevicornis)等偏暖温带近海种类为优势种[23];在胶州湾,中华哲水蚤、拟长腹剑水蚤、近缘大眼水蚤是冬春季(12月、1—5月)的优势种[24]。在苏北沿岸,真刺唇角水蚤、小拟哲水蚤、强壮滨箭虫(Aidanosagitta crassa)、中华哲水蚤和黑褐新糠虾(Neomysis awatschensis)为优势种[25];在东海长江口近海海域,春季,浮游动物结构较单一,中华哲水蚤和五角水母是主要优势种[26]。以上分析可见,三沙湾海域4、5月,浮游动物的种类组成,特别是优势种的种类组成,与我国暖温带海域浮游动物种类相似,具有暖温带浮游动物区系的种类特征。而与南海北部等典型的亚热带浮游动物区系不同。将5月和4月结果相比,5月的水温高于4月,虽然中华哲水蚤和纳嘎带箭虫仍为重要优势种,但中华哲水蚤优势度有所下降,五角水母和近缘大眼水蚤数量明显下降,退出优势种的地位。更多的亚热带近海种,包括球形侧腕水母和百陶带箭虫等成为了优势种。
5月到6月该海域浮游动物种类发生了明显的季节更替变化,物种更替率为69.39%。6月三沙湾海域浮游动物优势种类逐渐以亚热带近海种太平洋纺锤水蚤、瘦尾胸刺水蚤(C.tenuiremis)和中华假磷虾等为主,亚热带外海种也开始大量出现,例如精致真刺水蚤,微刺哲水蚤,驼背隆哲水蚤等,种类结构较为丰富(表1)。
相对5—6月种类组成的变化,8月、10月与6月之间的浮游动物物种更替率略低。6—8月物种更替率为55.74%,8—10月物种更替率为50.98%,均明显低于5—6月物种更替率。6—10月,三沙湾浮游动物优势种几乎都以亚热带外海种和亚热带近海种为主(表1),主要有中华假磷虾、亚强次真哲水蚤(Subeucalanus subcrassus)、太平洋纺锤水蚤、背针胸刺水蚤、微刺哲水蚤和百陶带箭虫等,其中,中华假磷虾为我国东南沿岸低盐海域特有种,主要分布在东海、南黄海小于33‰的海域,向南可扩布到厦门[27],为亚热带近海种。亚强次真哲水蚤则广泛分布于东海、台湾海峡和南海北部海域,夏秋季数量较大[27],为典型的亚热带外海种。
6月到10月之间种类组成变化比较(表1),6月优势种主要是太平洋纺锤水蚤和瘦尾胸刺水蚤等近海种,而8月,除了太平洋纺锤水蚤和背针胸刺水蚤等近海种外,更多的优势种为微刺哲水蚤、精致真刺水蚤和驼背隆哲水蚤等亚热带外海种,亚热带外海种无论在优势度还是丰度上均已经远远高于近海种。10月进一步形成了以肥胖软箭虫(Flaccisagitta enflata)、亚强次真哲水蚤等典型外海种为优势种的群落结构。该浮游动物群落结构特征与南方海域浮游动物组成极为相似。三门湾浮游动物优势种春季主要以中华哲水蚤和真刺唇角水蚤等暖温带近海种为主,夏季,则出现较多以驼背隆哲水蚤和太平洋纺锤水蚤为代表的亚热带种[28]。与之地理位置较为接近的罗源湾在5—7月间也主要以亚热带近海种为主,太平洋纺锤水蚤和中华假磷虾等有大量分布,夏秋季则出现较多亚热带高盐种类[29]。大亚湾等南部海湾也存在着湾内为高温低盐种,湾口为高温高盐种,且湾内群落结构较为复杂的特性[30]。
在三沙湾(表2,图2),从4月到5月,浮游动物呈现出暖温带种逐渐减少,亚热带种逐渐增加的过程,到了6月,优势种几乎为亚热带种构成。从6月到10月,浮游动物主要优势种虽都是亚热带种,但呈现出在外海种逐渐增加,近海种逐渐减少的过程。上述现象产生,与影响三沙湾海域水团的季节变化有关。
3.2 生态类群组成结构季节交替与水团变化的关系
4月,三沙湾外海域盛行东北季风,受到浙闽沿岸流的影响,水温较低,因此,浮游动物优势种以暖温带近海种为主(表2,图2),与春季瓯江口[31],三门湾[28]和长江口水域[26]同期浮游动物组成基本相似,显示出这些海域处于同一水团控制。5月,随着沿岸流减弱,海水温度升高,一些亚热带种成为优势种,直至6月几乎都以亚热带种构成。同时5—6月闽东北地区是雨季,导致大量淡水随径流注入湾内,海水盐度较低,导致5月和6月近海种成为主要优势种。随着6月西南季风逐渐强盛,自南携带南海高温水团和台湾暖流影响三沙湾海域,这一时期外海种开始随潮流进入湾内,成为6月种类组成的重要部分。到了8月,受副热带高压影响,当地降水减少[3],同时还因为台湾暖流加强,成为影响三沙湾的主要水团[32],高温高盐水团随潮流进入湾内,带入大量亚热带外海种,并迅速成为优势种的主要部分,暖温带种已经基本消失。10月,三沙湾海域持续受到台湾暖流的影响,台湾暖流影响累积的结果,使得近海种和暖温带种进一步减少,外海种优势度加强。
4、5月,近海种丰度比例高达97%左右,种类数比例均高于78%,而6月,这一比例明显下降并趋于相对稳定(表2,图2)。在适温性上虽表现为暖温带种比例逐月下降的趋势,但也能发现其中6、8、10月间的变化程度较小,与4、5月区分开。这一结论与聚类分析的结果相一致(图4),将4、5月并为A组,6、8、10月并为B组。说明,三沙湾浮游动物生态类群种类数季节变化较为明显,其中5月至6月是关键的转折点。
综上所述,三沙湾浮游动物生态类群组成结构的季节交替与东海南部近海和台湾海峡水团变化密切相关,4—5月是一个沿岸水团减弱,暖温带种减少的过程;5—6月沿岸水团影响已经几乎消失,然而浮游动物生态类群对水团环境变化响应是一个逐渐适应和调整的过程,反映在种类组成上,暖温带种继续减少,但未完全消失。6—10月是一个台湾暖流加强,近海种减少,外海种增加的过程。
3.3 生态类群种类数季节交替与水团变化的关系
通过对种类数季节变化(图2)和各月份不同生态类群种类数比例变化(图3)的分析,结果表明,4、5月与6、8、10月之间差别较大。4月,控制三沙湾海域的水团主要是暖温性沿岸流的水团,徐兆礼[33]指出,暖温带浮游动物种类数往往少于亚热带种类数,沿岸水团的种类数往往少于外海水团种类数。由于4月三沙湾浮游动物主要由暖温带种和近海种构成,亚热带种和外海种很少,这是4月总种类数最少的原因。5月,沿岸流水团消退,水温升高,加上当地径流量较大,亚热带近海种从上月的3种迅速增加到了7种,暖温带近海种从13种减少到8种,因此,总种类数略有增加,但仍保持较低水平。6月,台湾暖流开始影响湾外海域,亚热带外海种通过潮流携带进入湾内,同时径流量仍然较大,暖温带种尚未完全消失,因此,不同生态类群种类均出现在这个季节,导致总种类数急剧上升至全年最高值。在总种类数变化中,亚热带种贡献最多,其中亚热带外海种种类数增加最大,由4、5月的1种和4种增至22种,显示出台湾暖流对三沙湾浮游动物种类组成的影响明显。8月,台湾暖流占了主导地位,亚热带外海种略增至24种,亚热带近海种有所下降,暖温带近海种仍保持较低水平,以至于8月总种类数略低于6月。10月各个生态类群种类数均较6、8月有所下降,暖温带近海种下降,是因为台湾暖流影响持续了整个夏季,影响效应累积的结果,使得暖温带近海种种类数量降至最低。同时,由于台湾暖流影响开始减弱,导致亚热带外海种种类数减少,这也是10月总种类数略低于6月和8月的原因。
3.4 生态类群数量季节变化与优势种生态类群的关系
优势种是具有控制群落和反映群落特征的种类,是数量占比最多的关键种[10]。因此生态类群丰度特征,某种程度上可由优势种的丰度来决定。4月三沙湾浮游动物丰度较为明显的特征即存在中华哲水蚤单一优势种的现象,其丰度占该月总丰度比例的65%以上(表1),由此可见,4月之所以暖温带种丰度占主导地位(表3,图3),主要原因是中华哲水蚤作为暖温带种,具有较高的总丰度比例所致。5月优势种生态类群种类增加,中华哲水蚤丰度明显降低,优势种中有球型侧腕水母和百陶带箭虫等亚热带近海种加入补充,使得暖温带种丰度和亚热带种丰度接近平衡,亚热带种丰度略微占优的格局出现(表3,图3)。6月中华哲水蚤已不成为优势种,由亚热带近海种真刺唇角水蚤、太平洋纺锤水蚤、瘦尾胸刺水蚤和中华假磷虾等代替成为重要的优势种,占了很大丰度比例(表3,图3),因此,6月浮游动物总丰度主要由亚热带近海种构成。此外,6月起,优势种种类开始多元化,不再是较为单一优势种,而是多优势种均衡分布格局(表1),即存在多个优势种且单个优势种丰度较低的现象(表1)。
物种多样性指数H'值是反映浮游动物群落结构稳定性的指标,与种类数多少和丰度在种间分布均匀度有关。在暖温带种为优势种的群落结构,往往存在单一优势种,群落的丰度主要由该优势种构成,反映在多样性指数上,H'值往往较低,例如,4月生物多样性指数最低(图5)就是这个原因。在亚热带种为优势种的群落结构中,由于优势种种类数较多,丰度在不同优势种之间分布往往比较均匀,其多样性指数H'值明显高于暖温带种为优势种群落的H'值。同为亚热带种为优势种的群落结构,多样性指数H'值与种类数有一定的关系,6月种类数最多,使得该月多样性指数H'值最高。
可见,优势种的生态类群属性,决定了该群落总丰度生态类群的构成,决定了群落的丰度在不同种间分布格局,从而影响到该群落物种多样性大小,是决定该群落结构稳定性的重要因素。
从以上分析可见,三沙湾浮游动物总丰度主要由优势种丰度的贡献决定的。优势种的不同生态属性决定了浮游动物群落结构,暖温带种为优势种的群落结构显示出优势种种类数较少,单一优势种丰度较大的浮游动物群落结构格局;而亚热带种为优势种的群落结构则显示出优势种种类数较多,各个优势种丰度较小的结构格局。
致谢:刘守海、叶金清等对样品采集给予帮助,沈晓民老师、周时强老师帮助写作,特此致谢。
[1] Zhang C Y,Shang S L,Chen D W,Shang S P.Short-term variability of the distribution of Zhe-Min coastal water and wind forcing during winter monsoon in the Taiwan Strait.Journal of Remote Sensing,2005,9(4):452-458.
[2] Lin J H,Wang X P,Chen R X.Distribution of planktonic copepods in Sansha Bay,Fujian.Marine Science Bulletin,1997,16(6):13-19.
[3] The Chinese local chronicles compilation committee in Fujian province.The Chinese Local Chron icles of the Regional Meteorological Affairs in Fujian province.Beijing:Fangzhi Press,1996.
[4] Xu Z L,Chao M,Chen Y Q.Distribution characteristics of zooplankton biomass in the East China Sea.Acta Oceanologica Sinica,2004,26(3):93-101.
[5] Xu Z L,Wang Y L,Bai X M,Chen Y Q.An ecological study on zooplankton in the Changjiang estuary.Journal of Fishery Sciences of China,1999,6(5):55-58.
[6] Gao Q,Xu Z L.Species composition and diversity of zooplankton in Oujiang River estuary in summer and autumn.Chinese Journal of Ecology,2009,28(10):2048-2055.
[7] Lin J H,Chen R X,Dai Y Y,Lin M,Huang Y J.Ecological studies of zooplankton in Dongshan Bay.Journal of Oceanography in Taiwan Strait,1991,10(3):205-212.
[8] Lin J H,Chen R X,Lin M,Dai Y Y.Distribution of zooplankton in Sansha Bay and its comparison with that in Xinghua Bay and Dongshan Bay.Journal of Oceanography in Taiwan Strait,1998,17(4):426-432.
[9] Chen G B,Dai H,Xia Y J.Distributive characteristics of zooplankton in ecology monitoring area of Sandu Bay(part in shore).Marine Environmental Science,2007,26(5):475-478.
[10] Shen G Y,Shi B Z.Marine ecology.Beijing:Science Press,2002.
[11] Xu Z L.Analysis on the indicator species and ecological groups of pelagic ostracods in the East China Sea.Acta Oceanologica Sinica,2008,27(6):83-93.
[12] Xu Z L.Water environment adaptability and ecological groups of Hydroidomedusae in East China Sea.Chinese Journal of Applied Ecology,2009,20(1):177-184.
[13] Xu Z L.Mathematical analysis on adaptation of Heteropoda to different temperature and salinity in the East China Sea.Journal of Fishery Sciences of China,2007,14(6):932-938.
[14] Xu Z L,Gao Q,Chen H,Chen J J,Cai M.Ecological adaptation of pelagic Cladocera and Cumacea in East China Sea.Chinese Journal of Ecology,2007,26(11):1782-1787.
[15] Xu Z L,Chen Y Q.Relationships between dominant species of Chaetognatha and environmental factors in the East China Sea.Journal of Fishery Sciences of China,2005,12(1):76-82.
[16] Xu Z L,Shen A L.Pelagic Mysidacea in East China Sea and its significance as the indicator of water mass.Chinese Journal of Applied Ecology,2007,18(10):2347-2353.
[17] Xu Z L,Sun J,Lin M.Statistical analysis to ecological group of Thaliacea in the East China Sea.Acta Ecologica Snica,2008,28(11):5698-5705.
[18] Xu Z L.Determining optimal temperature and salinity of Lucifer(Dendrobranchiata:Sergestoidea:Luciferidae)based on field data from the East China Sea.Plankton& Benthos Research,2010,5(4):136-143.
[19] Xu Z L.Study on adaptation and ecotype of pelagic Ostracoda in the East China Sea.Acta Oceanologica Sinica,2007,29(5):123-131.
[20] Zhao Z M,Zhou X Y.Introduction to ecology.Chongqing:Chongqing Branch of Scientific and Technical Documents Publishing House,1984.
[21] Meng F,Qiu J W,Wu B L.Zooplankton of the Yellow Sea Large Marine Ecosystem.Journal of Oceanography of Huanghai& Bohai Seas,1993,11(3):30-37.
[22] Uye S.Why does Calanus sinicus prosper in the shelf ecosystem of the Northwest Pacific Ocean?Journal of Marine Science,2000,57(6):1850-1855.
[23] Wang Z L.Distribution of pelagic copepods in Dalian Bay.Journal of Oceanography of Huanghai& Bohai Seas,1995,13(1):47-54.
[24] Zhou K.Species composition and spatio-temporal distribution of dominant species of zooplankton in Jiaozhou Bay.Graduate school,Chinese Academy of Sciences,2006.
[25] Tian F G,Xu Z L.Ecological characters of zooplankton in middle area of Subei Shoal in spring and summer.Marine Environmental Science,2011,30(3):316-320.
[26] Xu Z L.Relationship between red tide occurrence and zooplankton communities structure in the coastal sea of East China Sea in spring.China Environmental Science,2004,24(3):257-260.
[27] Zhen Z,Li S J,Xu Z Z.Marine planktology.Beijing:Ocean Press,1984.
[28] Liu Z S,Wang C S,Zhang Z N,Liu C G,Yang G M.Seasonal dynamics of zooplankton and microzooplankton grazing impact in Sanmen Bay,China.Acta Ecologica Sinica,2006,26(12):3931-3941.
[29] Hung J Q,Li S J,Chen Y P.On the species composition and distribution of zooplankton at Luoyuan Bay of Fujian,China.Journal of Xiamen University(Natural Science),1989,28(sup.):85-95.
[30] Lian X P,Tan Y H,Huang L M,Chen Q C,Li K Z,Liu Y H.Space-time variations and impact factors of macro-meso zooplankton in Daya Bay.Marine Environmental Science,2011,30(5):640-645.
[31] Chen L,Xu Z L,Yao W M,Zhang Z,Chen S,Cai S W,Bao X P.The distribution of nutrients,phytoplankton and zooplankton in spring of the Oujang River estuary.Acta Ecologica Sinica,2009,29(3):1571-1577.
[32] Jan S,Wang J,Chern C S,Chao S Y.Seasonal variation of the circulation in the Taiwan Strait.Journal of Marine Systems,2002,35:249-268.
[33] Xu Z L.The past and the future of zooplankton diversity studies in China seas.Biodiversity Science,2011,19(6):635-645.
参考文献:
[1] 张彩云,商少凌,陈德文,商少平.冬季浙闽沿岸水分布的短期变动与风的关系初探.遥感学报,2005,9(4):452-458.
[2] 林景宏,王小平,陈瑞祥.福建三沙湾浮游桡足类的分布.海洋通报,1997,16(6):13-19.
[3] 福建省地方志编纂委员会.中华人民共和国地方志:福建省志·气象志.北京:方志出版社,1996.
[4] 徐兆礼,晁敏,陈亚瞿.东海浮游动物生物量分布特征.海洋学报,2004,26(3):93-101.
[5] 徐兆礼,王云龙,白雪梅,陈亚瞿.长江口浮游动物生态研究.中国水产科学,1999,6(5):55-58.
[6] 高倩,徐兆礼.瓯江口夏、秋季浮游动物种类组成及其多样性.生态学杂志,2009,28(10):2048-2055.
[7] 林景宏,陈瑞祥,戴燕玉,林茂,黄依金.东山湾浮游动物的生态.台湾海峡,1991,10(3):205-212.
[8] 林景宏,陈瑞祥,林茂,戴燕玉.三沙湾浮游动物的分布及其与兴化湾、东山湾的比较.台湾海峡,1998,17(4):426-432.
[9] 陈国斌,戴红,夏永健.三都湾生态监控区(近海部分)浮游动物的分布特征.海洋环境科学,2007,26(5):475-478.
[10] 沈国英,施并章.海洋生态学.北京:科学出版社,2002.
[12] 徐兆礼.东海水螅水母环境适应与生态类群.应用生态学报,2009,20(1):177-184.
[13] 徐兆礼.东海浮游异足类环境适应分析.中国水产科学,2007,14(6):932-938.
[14] 徐兆礼,高倩,陈华,陈佳杰,蔡萌.东海浮游枝角类和涟虫类生态适应性.生态学杂志,2007,26(11):1782-1787.
[15] 徐兆礼,陈亚瞿.东海毛颚类优势种及与环境的关系.中国水产科学,2005,12(1):76-82.
[16] 徐兆礼,沈盎绿.东海浮游糠虾类生态类型划分及其对水团的指示作用.应用生态学报,2007,18(10):2347-2353.
[17] 徐兆礼,孙军,林茂.东海海樽类生态类群统计分析.生态学报,2008,28(11):5698-5705.
[19] 徐兆礼.东海浮游介形类生态适应分析.海洋学报,2007,29(5):123-131.
[20] 赵志模,周新远.生态学引论.重庆:科学技术文献出版社重庆分社,1984.
[21] 孟凡,丘建文,吴宝铃.黄海大海洋生态系的浮游动物.黄渤海海洋,1993,11(3):30-37.
[23] 王真良.大连湾水域浮游桡足类的分布.黄渤海海洋,1995,13(1):47-54.
[24] 周克.胶州湾浮游动物的物种组成与优势种时空分布特征.中国科学院研究生院,2006.
[25] 田丰歌,徐兆礼.春夏季苏北浅滩大丰水域浮游动物生态特征.海洋环境科学,2011,30(3):316-320.
[26] 徐兆礼.东海近海春季赤潮发生与浮游动物群落结构的关系.中国环境科学,2004,24(3):257-260.
[27] 郑重,李少菁,许振祖.海洋浮游生物学.北京:海洋出版社,1984.
[28] 刘镇盛,王春生,张志南,刘诚刚,杨关铭.三门湾浮游动物的季节变动及微型浮游动物摄食影响.生态学报,2006,26(12):3931-3941.
[29] 黄加祺,李少菁,陈亚萍.福建罗源湾浮游动物的种类组成和数量分布.厦门大学学报(自然科学版),1989,28(增刊):85-95.
[30] 连喜平,谭烨辉,黄良民,陈清潮,李开枝,刘永宏.大亚湾大中型浮游动物的时空变化及其影响因素.海洋环境科学,2011,30(5):640-645.
[31] 陈雷,徐兆礼,姚炜民,张钊,陈 胜,蔡圣伟,鲍旭平.瓯江口春季营养盐、浮游植物和浮游动物的分布.生态学报,2009,29(3):1571-1577.
[33] 徐兆礼.中国近海浮游动物多样性研究的过去和未来.生物多样性,2011,19(6):635-645.
猜你喜欢
生物学通报(2021年9期)2021-07-01
辽宁化工(2021年5期)2021-06-03
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30
亚热带农业研究(2019年2期)2019-09-05
江西建材(2018年1期)2018-04-04
星星·诗歌原创(2017年5期)2017-08-13
湖南农业(2015年7期)2015-03-20
中国果菜(2015年2期)2015-03-11
亚热带资源与环境学报(2015年4期)2015-01-22
断块油气田(2014年5期)2014-03-11
