
云南4个地区高山姬鼠线粒体细胞色素b遗传分化的研究
2013-12-03 07:23朱万龙姜文秀王政昆
生物学杂志 2013年4期
朱万龙,姜文秀,王政昆
(云南师范大学生命科学学院,昆明650500)
哺乳动物的分布往往会受到环境变化、气候波动等因素的影响,而这些影响往往表现为动物的遗传变异[1]。分子标记作为种群遗传结构和地理变异的重要研究手段,被广泛地应用于遗传变异问题的探讨[2]。线粒体DNA是一种广泛使用的分子标记,其特点是进化速度快、母系遗传和非重组变异等[3],而其包括的细胞色素b(Cyt b)基因适合于种群水平差异的检测,普遍地用于动物的系统进化和分类研究[4]。
高山姬鼠(Apodemus chevrieri)属于姬鼠属,典型的古北界种类,主要分布于中国西南的横断山脉及其周围地区[5]。中国的西南地区是古北界寒带物种南迁,中南半岛热带物种北移的交汇地,是生物多样性研究的热点地区之一;地处该区的云南省,西为横断山区,东为云贵高原,地势西高东低、海拔变化剧烈、自然环境的地带性和非地带性变化明显,这些可能导致丰富的物种多样性,也可能存在生态物种的形成。栖息横断山和云贵高原的高山姬鼠,可能为了适应不同的生存环境,出现种间水平的差异。因此,本研究利用细胞色素b基因对云南4个地区高山姬鼠的遗传变异进行研究,为高山姬鼠遗传学研究提供实验依据。
1 材料和方法
1.1 实验动物
高山姬鼠捕自云南省中甸(3 305~3 296 m;N27°83'~26°84',E99°68'~ 99°69')、丽江(2 556 ~ 2 590 m;N26°71'~26°96',E100°20'~ 100°26')、剑川(2 556~2 590 m;N26°15'~ 26°45',E99°40'~ 99°55')、昆明(2 125 ~ 2 139 m;N25°05 ~ 25°15',E102°31' ~ 102°32')的农田及灌木丛中,用冷冻保存(-70℃)的肝脏标本提取线粒体DNA。
1.2 DNA的提取
基因组DNA参照王文和施立明的碱变性法[6]进行提取。
1.3 PCR扩增
Cyt b 的 PCR 引物采用 Pääbo 等[7]和 Irwin 等[8]报道的哺乳动物Cyt b通用引物,引物序列为L14724 5'-CGA AGCTTG ATA TGA AAA ACC ATC GTT G-3',H15915 5'-CGG AAT TCC ATT TTT GGT TTA CAA GAC-3',引物覆盖长度为1 140 bp。
扩增体系为50 μL,其中含有10 μmol/L引物各1 μL、50 ng左右的 DNA 模板、10 × Buffer 5 μL、2 U Taq酶、25 mmol/L MgCl22 μL 和 10 mmol/L dNTPs 1 μL。扩增条件:94℃预变性3 min;94℃变性30 s,51℃退火30 s,72℃延伸1 min,30 个循环;72℃延伸5 min;4℃终止反应。实验过程中使用空白对照来检测PCR是否污染。
1.4 PCR产物纯化和测序
扩增产物经琼脂糖凝胶电泳检测后,使用Glassmilk DNA纯化回收试剂盒(博大泰克)进行割胶回收和纯化,回收产物直接送大连宝生物工程有限公司测序。
1.5 序列分析
Cyt b序列使用ClustalX软件[9]进行比对。利用MEGA4.0[10]软件包分析序列特征、碱基组成、转换与颠换值,计算遗传距离;通过DnaSP 4.0[11]软件计算单倍型多样性(H)、单倍型的平均核苷酸差异数(K)和核苷酸多样性(Pi);计算种群间的平均核苷酸差异数(Kxy)、遗传分化指数(Fst)、核苷酸的分歧度(Dxy)、核苷酸净遗传距离(Da)和基因流(Nm)[12]。在MEGA 4.0软件中,使用Kimura两参数,以非加权组平均法(unweighted pair-group method with arithmetic means,UPGMA)构建系统发生关系,系统树各分支的自举检验值由1 000次重复检验所得,使用黑线姬鼠(Gene Bank号:JL1105)的同源区序列作为外群。在ARLEQUIN3.10[13]软件中进行Fst和种群间序列平均差异的分析以及分子变异分析 (AMOVA),用Network 4.5软件构建进化网络关系图。
2 结果
2.1 DNA的PCR扩增
扩增结果经琼脂糖凝胶电泳检测未发现非特异性条带,空白对照组未出现扩增产物,没有出现亮带,表明未受外源DNA的污染(图1)。

图1 高山姬鼠mtDNA细胞色素b序列扩增结果Fig 1 The amplification on result of Cyt b gene sequence in A.chevrieri
2.2 Cyt b基因序列变异
本研究共获得4个种群43只高山姬鼠的线粒体Cyt b基因全序列(1 140 bp)中共有50个核苷酸变异位点(占全部碱基数4.39%),包含简约信息位点21个,29个单变异位点,转换和颠换(TS/TV)比为7.58,其A、T、G、C序列平均碱基组成为:30.8%、29.0%、12.8%和27.5%,其中A+T的比例显著高于C+G。29个变异位点共定义22种单倍型,其中种群间的共享单倍型仅有1个(Hap3),且均为3个横断山种群所共享。单倍型的分布信息见表1。

表1 高山姬鼠的单倍型分布Table 1 Haplotypes of the population of in A.chevrieri
2.3 遗传多样性和种群间遗传差异
在4个种群中,昆明种群的Hd最高,丽江种群最低;昆明种群 Pi最高,剑川地区最低(表2)。Kxy、Dxy、Da均是昆明种群比横断山3个种群高。中甸、丽江、剑川3个种群间Nm>1,表明群体间的基因流的水平较高,群体间遗传分化较小;昆明种群与其它3个种群间Nm<1,说明群体可能由于遗传漂变而发生了分化(表3)。
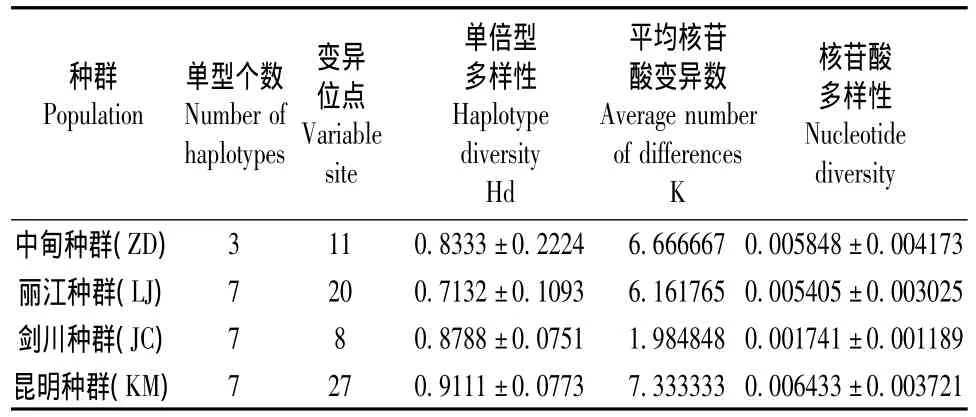
表2 高山姬鼠Cyt b基因序列的变异位点及遗传多样性Table 2 Variable sites and genetic diversity based on Cyt b gene sequence in A.chevrieri
在4个种群中,除中甸和丽江种群间Fst不显著外,其余各种群间的遗传差异显著;同时,种群序列平均差异除中甸和丽江种群差异不显著外,其余各种群间的遗传差异显著(表4)。分子变异分析(AMOVA)表明,遗传变异在种群内为60.80%,而种群间为39.20%,种群内变异大于种群间变异(表5)。
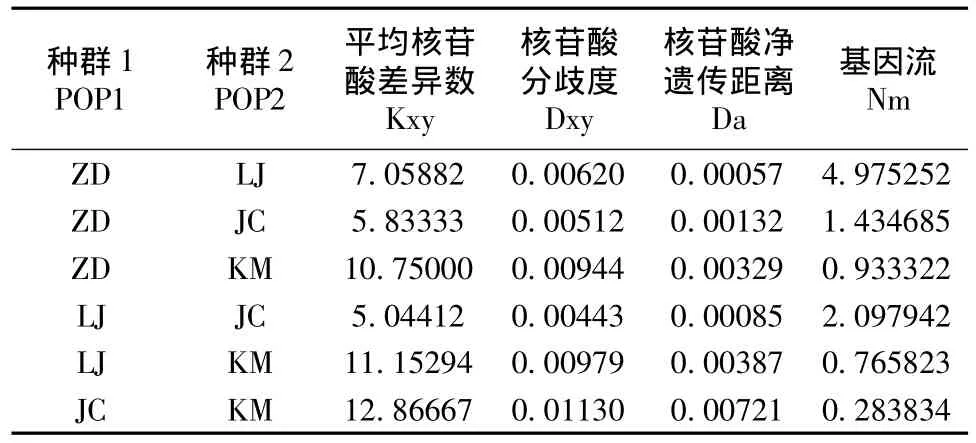
表3 高山姬鼠各种群遗传分化及基因流Table 3 Genetic differentiation and Nm values among different populations in A.chevrieri

表4 高山姬鼠4个种群之间遗传差异(Fst)及其显著性(对角线下)和修正后的种群间序列平均差异(PiXY-(PiX+PiY)/2)及其显著性(对角线上)Table 4 F-statistics test for pairwise population(below the diagonal)and P value of F-statistics test and corrected average pairwise difference(PiXY-(PiX+PiY)/2)(above the diagonal)between four populations in A.chevrieri
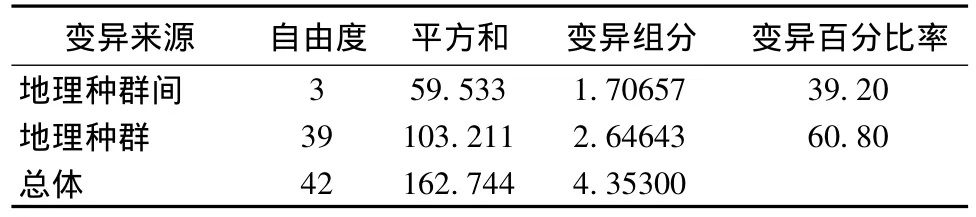
表5 高山姬鼠细胞色素b基因的分子变异分析Table 5 AMOVA of mitochondrial DNA Cyt b gene in A.chevrieri
2.4 单倍型系统发生分析
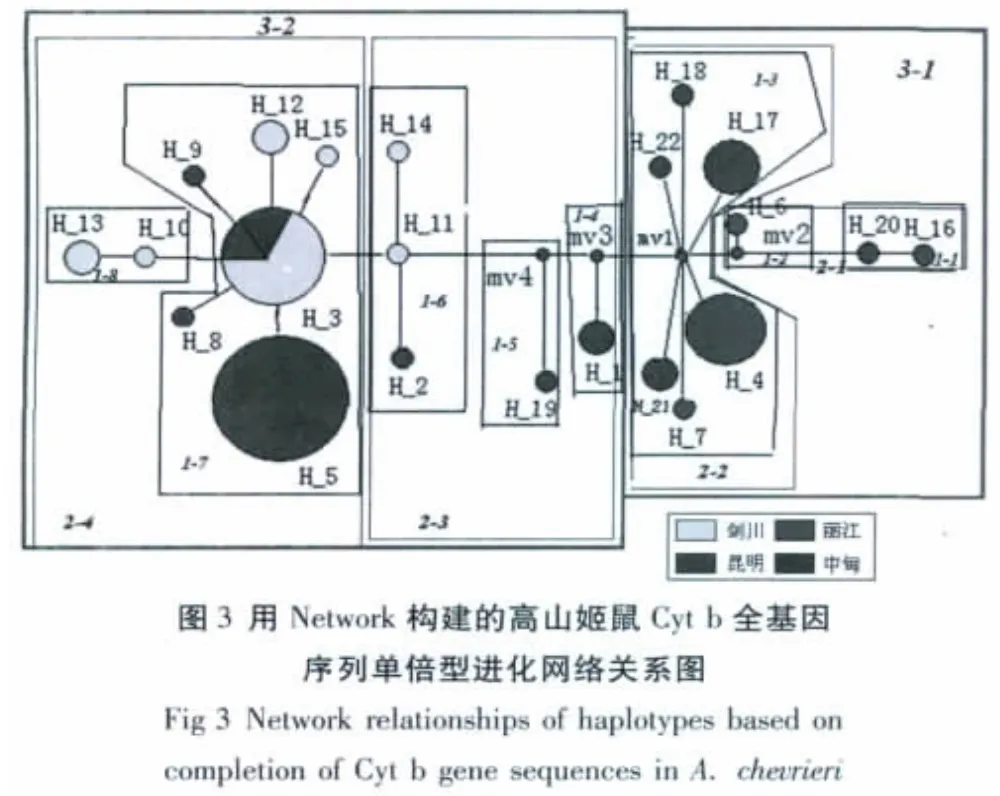
以黑线姬鼠为外群,22个单倍型在UP树中明显聚为两支(A、B),见图2。其中来自横断山区(中甸、丽江、剑川)构成一支(A),其余个体聚为另外一支(B),这一支除中甸、丽江、剑川种群外,还有来自昆明的种群。以Network构建的单倍型进化网络关系图显示,除少部分单倍型是横断山和昆明混杂外,进化网络总体分为两大支,一支是横断山种群,一支是昆明种群(图3)。
3 讨论
遗传多样性的大小最直接的表达形式是遗传变异的高低[14],而遗传多样性的高低与物种的适应能力等有关[15]。单倍型间的平均遗传距离和核苷酸多态性是遗传变异重要的衡量指标[16],国内外已有很多研究[17-19]。本研究结果表明在4个种群中,昆明种群单倍型多样性和核苷酸多样性最高。此外,60.8%的变异在种群内部,39.2%的变异存在于种群之间,遗传变异在种群间和种群内部均有发生,种群内的遗传变异较种群间的大,并且发生在种群内的遗传变异较发生在种群间的遗传大。由此可以推断出,云南4个地区的高山姬鼠种群的演变和发展过程已经出现了一定程度的遗传变异。

地理环境和生活习性的不同会引起种群的分化,进而导致种群间出现遗传分化,衡量遗传分化的指标有基因分化系数(Fst)和基因流(Nm)等。Fst表示不同种群之间遗传差异的程度[20],Nm则表示不同种群之间由个体的迁移所产生的基因流动[21,22]。距离隔离理论[23]认为遗传分化的程度会随着地理距离的增加而增加[24]。本研究Fst结果表明高山姬鼠的4个地理种群之间的遗传差异除中甸和丽江种群之间不显著外,其它地理种群之间的差异都极显著,表明种群之间有较高的遗传分化。昆明种群与其他3个种群间的基因流值相对较小,表明昆明与其他种群间的基因交流没有其他种群之间基因交流频繁,说明昆明与其它种群之间可能存在地理阻隔。从这4个种群的地理位置来看,昆明种群与其余3个种群之间的地理距离最远,中甸、剑川、丽江之间地理距离较近。因此可以初步推测,地理距离可能是影响高山姬鼠各种群间遗传分化的一个重要因素。以上所有结果表明高山姬鼠在滇中昆明地区的遗传多样性显著高于横断山区,并在横断山种群和滇中种群间已出现基因流明显减小的现象,推测横断山种群和昆明种群已有了较明显的种群遗传分化,尽管分化尚未完全形成。
大约200万年前,高山姬鼠的近缘种黑线姬鼠由欧洲扩散到亚洲,进而扩张至整个古北界,并分化出高山姬鼠[25];横断山区核心地带形成时间距今约2300万年至6500万年前的古近纪,云贵高原形成时间是距今约3600万年至5300万年前的第三纪始新世时期[26],这表明在横断山形成之后,高山姬鼠的祖先种才从欧洲或者是亚洲北部扩张进入横断山,推测种群扩散方向可能从横断山地区到昆明地区。22个单倍型在系统发生树中明显聚为两支(横断山种群和滇中昆明种群),进化网络关系表明昆明种群处于进化分支最末端,推测种群进化方向可能从横断山地区到昆明地区,支持了姬鼠属从北向南扩散的理论。此外,经单倍型系统树和Network网络关系分析表明横断山种群和滇中昆明种群间已经出现了遗传分化,尽管还有部分单倍型混杂、分化不完全。
综上所述,本研究显示4个地区的高山姬鼠已经出现一定的遗传变异,推测高山姬鼠种群进化方向可能从横断山地区到昆明地区,支持了姬鼠属从北向南扩散的理论。
[1]Culling M A,Janko K,Boron A,et al.European colonization by the spined loach(Cobitis taenia)from Ponto-Caspian refuge based on mitochondrial DNA variation[J].Molecular Ecology,2006,15:173-190.
[2]Nichols R A,Hewitt G M.The genetic consequences of long dispersal during colonization[J].Heredity,1994,72:312-317.
[3]Birky C W,Fuerst P,Maruyama T.Organelle gene diversity under migration,mutation,and drift:equilibrium expectations,approach to equilibrium,effects of heteroplasmic cells,and comparison to nuclear genes[J].Genetics,1989,121(3):613-627.
[4]Hochachka M.Biochemistry and molecular biology of fishes[M].London:Elsevier Science Publishers,1993,1-38.
[5]郑少华.川黔地区第四纪啮齿动物化石[M].北京:科学出版社,1993.
[6]王 文,施立明.一种改进的线粒体DNA提取方法[J].动物学研究,1993,14(2):197-198.
[7]Pääbo S,Higuchi R G,Wilson A C.Ancient DNA and the polymerase chain reaction[J].Journal of Biological Chemistry,1989,264:9706-9712.
[8]Irwin D M,Kocher T D,Wilson A C.Evolution of cytochrome b gene of mammals[J].J Mol Evol,1991,32:128-144.
[9]Thompson J D,Gibson T J,Plewniak F,et al.The clustal x windows interface:exible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Res,1997,24:4876-4882.
[10]Tamura K,Dudley J,Nei M,et al.MEGA4:molecular evolutionary genetics analysis(MEGA)software version 4.0[J].Molecular Biology and Evolution,2007,24:1596-1599.
[11]Rozas J,Rozas R.DnaSP version3:an integrated program for molecular population genetics and molecular evolution analysis[J].Bioinformatics,1999,15:174-175.
[12]Hudson R R,Slatkin M,Maddison W P.Estimation of levels of gene flow from DNA sequence data[J].Genetics,1992,132:583-589.
[13]Excoffier L,Smouse P E,Quattro J M.Analysis of molecular variance inferred from metric distances among DNA haplotypes:application to human mitochondrial DNA restriction data[J].Genetics,1992,131:476-494.
[14]Bullerwell C E,Gray M W.Evolution of the mitochondrial genome:protest connections to animals,fungi and plants[J].Current Opinion in Microbiology,2004,7(5):528-534.
[15]Kimura M.A simple method of estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences[J].Molecular Evolution,1980,16:111-120.
[16]牛华锋,陈玉林,任战军,等.中国绵羊品种mtDNA遗传多态性与系统进化研究[J].中国农学通报,2011,27(17):21-25.
[17]姜艳艳,孔晓瑜,喻子牛,等.黄海蓝点马鲛mtDNA D-loop序列变异分析[J].中国水产科学,2003,10(3):177-183.
[18]杨玉慧,张德兴,李义明,等.中国黑斑蛙种群的线粒体DNA多样性和生物地理演化过程的初探[J].动物学报,2004,50(2):193-201.
[19]宁恕龙,周立志,张保卫,等.基于线粒体细胞色素b基因的中国大沙鼠系统地理格局[J].动物学报,2007,53(4):630-640.
[20]Nei M.Molecular evolutionary genetics[M].New York:Columbia University Press,1987.
[21]Slatkin M.Gene flow and the geographic structure of nature populations[J].Science,1987,236:787-792.
[22]Millar C L,Libby W J.Strategies for conserving clinal,Ccotypic,an adisjunct population diversity in widespread species[M].New York:Oxford University Press,1991,149-170.
[23]Wright S.Isolation by distance [J].Genetics,1943,28:114-138.
[24]Floyd C H,Van Vuren D H,May B.Marmots on great basin mountain tops:using genetics to test a biogeographic paradigm[J].E-cology,2005,86(8):2145-2153.
[25]Suzuki H,Filippucci M,Chelomina G N,et al.A biogeographic view of Apodemus in Asia and Europe inferred from nuclear and mitochondrial gene sequences[J].Biochemical Genetics,2008,46:329-346.
[26]李炳元,王富葆.滇西北、川西南地区地貌的基本特征.青藏高原研究——横断山考察专集(二)[C].北京:科学技术出版社,1986,174-183.
猜你喜欢
湖北农业科学(2021年2期)2021-02-07
海南医学(2020年1期)2020-01-18
湖北农业科学(2019年20期)2019-12-11
经济研究导刊(2019年22期)2019-09-17
贵州农业科学(2019年7期)2019-08-03
电子制作(2019年24期)2019-02-23
湖北农业科学(2019年23期)2019-01-11
——唐天马长篇小说《横断山》简析
边疆文学(文艺评论)(2018年3期)2018-03-21
广西林业科学(2016年3期)2016-03-16
中国学术期刊文摘(2016年1期)2016-02-13
