
北京西山地区大山雀与其它鸟类种群种间联结分析
2013-12-21 05:00董大颖范宗骥李扎西姐陈卓琳王敏增王奇峰关文彬
生态学报 2013年20期
董大颖,范宗骥,李扎西姐,陈卓琳,王敏增,王奇峰,李 凯,关文彬,*
(1.北京林业大学,北京 100083;2.北京市西山试验林场,北京 100093)
种间联结是指不同物种在空间分布上的相互关联性,通常是由于群落生境的差异影响了物种的分布为引起的[1],是用来解释群落结构的重要数量特征之一[2]。引起种间联结有4种原因:(1)相似与不相似的环境需求;(2)一个种为另一个种创造了定居条件或者前者对后者施加了压力;(3)两者在竞争资源中相互排斥造成负联结;(4)两者在根系中通过物理的或化学的因素相互影响[3]。研究群落的种间联结性,能够有效地反映各物种在群落中的分布情况,各物种对环境因子的适应程度以及物种在特定环境因子作用下的种间相互关系,有助于进一步认清群落的结构、类型、及群落的演替趋势[4]。
在植物生态研究上已经在森林[5-6]、草原[7]、沼泽[8]、灌丛[9]、荒漠[10]等多种类型的植物群落中广泛的应用;微生物群落方面,对鸟类巢穴内的菌群进行了种间联结分析,同时,通过菌落的实例细致评价了不同种间联结测度的应用[11];在昆虫方面,种间联结运用于西双版纳聚果榕榕果小蜂的种间关系研究,发现其在昆虫生态学中同样可以清楚的反映群落内物种间的相互关系[12]。可见,种间联结应用范围很广,同时,在种群的动态、群落结构、演替方面具有重要意义,但种间联结在鸟类群落的研究却没有。
鸟类群落的物种组成并不是随机的,有许多因素影响着群落的结构组成[13]。但是,鸟类群落组成机制的研究一直受到鸟类流动、迁徙等鸟类空间不固定特征的困扰。如,鸟类群落物种是否存在种间联结?种间联结的尺度如何界定?存在种间联结的话,如何检验与测度?导致的原因是什么?因此,本研究以北京西山侧柏古树林及其周边针阔混交林、落叶阔叶林为对象,试图通过改进鸟类调查方法,引入表联结的检验与联结度测度的公式,以森林鸟类广布种大山雀为目标种,探讨鸟类群落种间联结的研究方法,结合鸟类的营巢、取食行为集团[14-16]、生态位[17]等研究,分析鸟类群落的种间联结的关系。以丰富鸟类群落学研究的科学方法,进而为鸟类及其栖息地的保护提供科学依据。
1 研究方法
1.1 研究地概况
北京静福寺位于海淀区四季青乡许木兰陀山麓,西山试验林场的中部,北纬40°00',东经116°11',海拔232 m,年平均温度为11.8℃,年平均降水量为680 mm,土壤以砂质土为主,多石砾,山体坡度在15°—35°之间。该地区植被类型属于暖温带落叶阔叶林,植被主要包括人工针叶林和次生落叶阔叶林,乔木以侧柏(Platycladus orientalis)、刺槐(Robinia pseudoacacia)、构树(Broussonetia papyrifera)等为主,灌木则以荆条(Vitex negundo)、扁担杆(Grewia biloba)、酸枣(Ziziphus jujuba)等为主。
1.2 鸟类调查方法
调查方法采用固定样带法月连续全天定位,植物群落样带(面积300 m×10 m)始于北纬40°00'01.4″东经116°11'10.3″终到北纬 40°00'13″东经 116°10'48.9″。鸟类观测范围是以 25 m 为半径的圆。调查时间从 2012年3月到2013年1月,每月进行2次调查,每日调查时间冬季从7:00—16:00,其余季节从6:00—18:00。记录用8×42倍双筒望远镜或裸眼见到和耳听到的鸟类实体种类、数量、栖息环境等,主要依据《中国鸟类野外手册》辨别种类。将遇见率≥1只/h作为优势种,0.1—1只/h作为常见种,小于0.1只/h作为稀有种。
1.3 群落种间联结的总体检验
总体相关性检验采用方差比率法[18]:按照方差比率法来检验多物种间的关联可说明在某地出现的多物种间是否存在显著的关联性,先假设群落物种间无显著关联,按下列公式计算检验统计量:
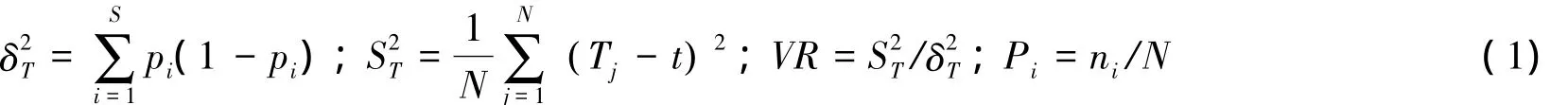
式中,S为总的物种数,N为总样方数,Ti为样方j内出现的研究物种总数,ni为物种i出现的样方数,t为样方中的平均数。在独立性假设条件下VR期望值为1。VR=1表示复合种间关联性是独立的;VR>1表示物种间表现出正的关联;VR<1表示物种间存在负关联。采用统计量W=N×VR来检验VR值偏离1的显著程度,若种间无关联,则W落入卡平方分布给出的界限概率为
种间联结度多采用2×2列联表给出的定性数据来测度[1]:

表1 2×2列联表Table 1 2×2 contingency table
1.4 种间联结的检验
χ2检验 用连续性校正[19]公式计算,这个校正公式考虑了边缘频率的变化[20-21]:
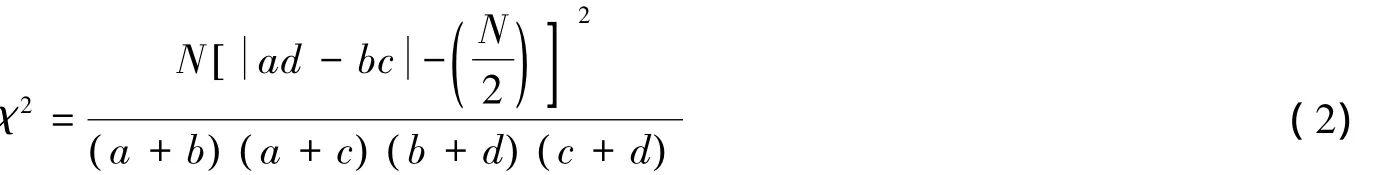
式中,n为取样总数,当χ2<3.841时,种间关联性不强;当χ2≥6.635时,种间有显著的生态联结性。同时,用ad-bc的正负作为种间联结性的判断标准,值大于零为正联结,反之则为负联结。
1.5 种间联结度的计算
(1)点相关系数PCC指数[22]
采用变化幅度由+1通过0而达到-1的有中心指数,表示种间的联结程度,PCC为负值时表示为负联结,其绝对值越大负联结度越高;PCC为正值时表现正联结,其值越大正联结度越高。

(2)成对物种间匹配系数
选用以下2个无负值变化的于0与1幅度变化的无中心指数(noncent redindices)来表示种间关联程度和相伴出现的机率,在“无关联”时等于0,在“最大关联”时为1。
共同出现百分率[23],也称作物种的群落系数[24]:


2 结果与分析
北京西山地区鸟类群落调查共65个鸟类物种,总体相关性检验,鸟类总体种间联结8月份为显著负联结,其余各月为显著正联结(表2)。大山雀的生态位宽度值较大,属广性分布[26]森林鸟类,而且是调查中出现的频率最高的,因而,本文以大山雀作为“目标种”,分析其与留鸟、夏候鸟、冬候鸟、旅鸟等不同居留型鸟类的联结关系。
2.1 种间联结检验与联结度测度指数的选择
联结度的测定及其统计意义的检验常常是混乱的[1],每个指数都有其自己的性质,同时它们能得出很不同的结果[27],因而,选取物种数量较多、大山雀联结种对较多的2012年4月26日全天调查数据作为日尺度的研究对象,通过对χ2统计量、点相关系数PCC、共同出现百分率PC、OI系数4个指数进行对比分析,筛选适合用于鸟类种间联结分析的联结度检验与测度指数。
由χ2检验可知,在18个物种的153个种对中,正负联结联结比为9∶8,其中6个种对呈现极显著正联结(P<0.01),大山雀与大嘴乌鸦(Corvus macrorhynchus)、黄腰柳莺(Phylloscopus proregulus)、喜鹊(Pica pica);大嘴乌鸦与黄腰柳莺、喜鹊;黄腰柳莺与喜鹊。显著正联结(0.01<p<0.05)有8个种对:大山雀与红嘴蓝鹊(Urocissa erythrorhyncha)、沼泽山雀(Parus palustris);大嘴乌鸦与红嘴蓝鹊、沼泽山雀;黄腰柳莺与红嘴蓝鹊、沼泽山雀;喜鹊与红嘴蓝鹊、沼泽山雀。大山雀、大嘴乌鸦、黄腰柳莺、喜鹊4个物种分别与北红尾鸲(Phoenicurus auroreus)、黄眉柳莺(Phylloscopus inornatus)、白眉鹀(Emberiza tristrami)、大斑啄木鸟(Dendrocopos major)、环颈雉(Phasianus colchicus)、黄喉鹀(Emberiza elegans)、灰头绿啄木鸟(Picus canus)、山斑鸠(Streptopelia orientalis)、山噪鹛(Garrulax davidi)、星头啄木鸟(Dendrocopos canicapillus)、银喉长尾山雀(Aegithalos caudatus)、棕头鸦雀(Paradoxornis webbianus)12个物种,组成的48个种对呈显著负联结(0.01<P<0.05)(图 1)。
点相关系数 PCC 指数值可划分 0≤PCC<0.3,0.3≤PCC<0.5,0.5≤PCC<0.7,PCC≥0.7,-0.3≤PCC<0,

图1 全天数据的χ2指数半矩阵Fig.1 Semi-matrix graph ofχ2 value for all-day data
Ochiai系数[25]:-0.5≤PCC<-0.3,-0.7≤PCC<-0.5,-1≤PCC<-0.7 等 8 个区间来表现种间联结程度[28]。PCC 为正值时表示正联结,数值越大联结越强,PCC≥0.7有42个种对,包含χ2检验中呈极显著正联结的6个物种对;0.5≤PCC<0.7 有 11 个物种对,0.3≤PCC<0.5 有 10 个物种对,均为不显著正联结;0≤PCC<0.3 有 18 个种对,其中有8个种对在χ2检验中表现出显著正联结。PCC为负值时表示负联结,指标绝对值越大负联结性越强,-0.3≤PCC<0有58个物种对,其中48个物种对在χ2检验中呈显著负联结关系;-0.5≤PCC<-0.3有11个种对;-0.7≤PCC<-0.5有3个物种对;这两个区间均为不显著负联结。可见,PCC指标在描述显著性种对时绝对值偏小,χ2检验的显著联结种对多数在PCC指标绝对值0.3以下,但PCC能准确的反映物种间的正负联结性,与χ2检验的结论基本一致(图2)。
共同出现百分率 PC 指数值可划分为 PC<0.3,0.3≤PC<0.5,0.5≤PC<0.7,PC≥0.7,4 个区间来表现种间联结程度[5]。PC指数越大正联结越强,PC≥0.7有48个种对,均为正联结关系,其中有6对χ2检验为极显著正联结,4对为显著正联结;0.5≤PC<0.7有15个种对,均为正联结种对,其中4个种对χ2检验为显著正联结;χ2检验为显著负联结的种对PC指标均在0.4以下(图3)。
OI指数值可划分为 OI<0.3,0.3≤OI<0.5,0.5≤OI<0.7,OI≥0.7,4 个区间来表现种间联结程度[29]。OI≥0.7有63个物种对,均为正联结,比同区间下的PC指标多了15个种对,其中6对为极显著正联结,有8个物种对呈显著正联结,0.5≤OI<0.7 有 27 个种对,其中 8 个物种对呈显著负联结,0.3≤OI<0.5 有 42 个种对,多数为负联结。本研究中OI指标表现种间联结时较PC指标偏大,不能很好的揭示联结关系(图4)。

图2 全天数据的PCC指数半矩阵Fig.2 Semi-matrix graph of the point correlation coefficients(PCC)for all-day data

图3 全天数据的PC指数半矩阵Fig.3 Semi-matrix graph of percentage co-occurrence(PC)for all-day data
综上所述,PCC、OI、PC三种关联度在反映种间联结情况上,OI、PC实质上是等效的,其值越高物种对同时出现的机率越大,但OI指标在本研究中数值过大,不能准确的反映联结关系,同时,PCC与PC的结果与整体常是无偏的[30-31],即使是在样方数较少的情况下,也与总体一致。因此,选择χ2检验、PCC、PC指数相结合能充分反映鸟类群落的种间联结。这一结论与菌物群落的种间联结研究是一致的[11]。

图4 全天数据的OI指数半矩阵Fig.4 Semi-matrix graph of Ochiai index(OI)for all-day data
2.2 测度时间尺度确定
植物的种间联结性跟样方大小有较大关系,种间相互作用有一定的空间范围,超过一定的界限就不会有相互作用[6]。由于鸟类是活动的,本研究调查采用林中视能及的以25 m为半径的圆形样方组成的样带进行全天候定位观测[32],这里暂且不考虑空间问题,而是针对鸟类的活动性,探讨在时间尺度上的种间联结的差异。
2.2.1 日尺度数据分析
将4月份观测的数据划分为全天、上午、下午3个尺度,分别进行χ2检验、PCC、PC指数进行联结分析,绘出星座图(图5),探讨在时间维度上鸟类联结性的变化情况。
χ2检验,全天、上、下午共出现18、17、12个种,全天数据有62个显著联结种对,上、下午的显著联结种对别为33、30个,与大山雀有显著联结的种对依次是17、16、11个,分别占各时间尺度总体数量的27%、48%、37%,可见与大山雀有关的种对在总体数量中占有很大比例(图1;图5a,b)。
PCC指数,在-0.3≤PCC<0区间内全天、上、下午分别有58、42、24个种对,全天种对数将近下午的2倍,与大山雀组成的种对依次分别是12、11、7对,占各时间尺度的21%、26%、29%。当PCC≥0.7时,在全天、上、下午的种对数分别为42、31、13个,全天种对数将近下午的3倍,与大山雀组成的种对依次分别是3、2、2对,占各时间尺度的7%、6%、15%(图2;图5c,d)。
PC指数,在区间PC≥0.7上全天、上、下午的种对数分别是47、36、13个,全天种对数将近下午的3倍,与大山雀有关的种对数依次分别是4、3、2对,占各时间尺度的9%、8%、11%。0≤PC<0.3区间内全天、上、下午的种对数分别是73、59、34个,全天种对数将近下午的2倍,与大山雀有关的种对依次是11、10、6个,占各时间尺度的 15%、17%、18%(图 3;图 5e,f)。

图5 不同时间尺度的种间联结的星座图Fig.5 The constellation diagrams of temporal scales
2.2.2月尺度的数据分析
由于鸟类的居留、繁殖、迁移等季节动态性特征,仅以日尺度上分析种间联结存有不确定性。因而,用月连续全天调查数据,计算大山雀与4种居留型的PCC月联结频率。
PCC月联结频率由正、负联结月份数分别除以11(调查取样为11个月,2月份数据由于春节原因没有调查)来计算,这里假设留鸟全年12个月均栖息于研究地区。然而,对于夏候鸟、冬候鸟、旅鸟而言,其在研究地区的全年活动时间并不是12月,北京地区夏候鸟、冬候鸟驻留的时间一般分别在3—10月份和10—3月份,旅鸟春、秋季迁徙时停留在北京的时间分别在3—5月份和8—10月份。因此,为了保证PCC月联结频率可比性,对于夏候鸟,采用驻留期PCC月联结频率,由正、负联结月份数分别除以除以8(北京地区夏候鸟最长驻留月份数)来计算;对于冬候鸟,采用驻留期PCC月联结频率,由正、负联结月份数分别除以除以6(北京地区冬候鸟最长驻留月份数)来计算;对于旅鸟,采用停留期PCC月联结频率,由正、负联结月份数分别除以除以6(北京地区旅鸟最长停留月份数)来计算。
2.3 大山雀与其他物种联结性分析
χ2检验显著性分析可知,大山雀与留鸟的显著正关联种对共有61个,占所有显著种对的31%,联结关系非常密切。进一步结合鸟类的营巢和取食习性集团[15-16]的划分(附录2)、生态位等来分析大山雀与四种居留型鸟类的联结关系:
2.3.1 大山雀与留鸟物种联结性分析
根据χ2检验、PCC、PC与PCC的月联结频率(表2),将大山雀与其他留鸟的联结可划分成3个联结组5类,正联结组:①大嘴乌鸦、喜鹊、灰喜鹊、红嘴蓝鹊;正负均等组:②沼泽山雀(Parus palustris);负联结较强的组:③银喉长尾山雀(Aegithalos caudatus)、褐头山雀,④环颈雉、山噪鹛(Garrulax davidi)、大斑啄木鸟(Dendrocopos majo r),⑤灰头绿啄木鸟(Picus canus)、小嘴乌鸦(Corvus corone)。
(1)大山雀与大嘴乌鸦、喜鹊、灰喜鹊、红嘴蓝鹊的PCC月联结频率正联结较高,达到73%以上(表2),同时,极显著以及显著正联结月份的共同出现比率PC、PCC多大于0.5,说明大山雀与这四种的正联结强度很大。大嘴乌鸦、喜鹊、灰喜鹊、红嘴蓝鹊均属于鸦科鸟类,表明大山雀与鸦科鸟类的联结性较高,这四种鸟类属于林冠树枝营巢集团,大山雀是林冠取食集团[16],大山雀与以上物种在栖息空间上有重叠,同时大山雀、大嘴乌鸦、喜鹊、灰喜鹊、红嘴蓝鹊均为西山林场的优势鸟类物种[32],共同遇见率高(附录1)。
(2)大山雀与沼泽山雀的PCC正负联结频率比是1∶1,联结频率相同,χ2检验显著正联结为4、5、6月份(繁殖期),PC指标均大于0.5;显著负联结为3、7、10、11月份。两物种的栖息生境与取食习性相近,生态位重叠度很大[33],虽然均为洞巢与林冠食虫集团[34],但两物种在取食高度,树冠层取食位置(水平和垂直),取食树种、取食食物大小和数量等方面存在差异,使竞争减少到最低程度[35],说明是繁殖行为导致了正联结。
(3)大山雀与褐头山雀、银喉长尾山雀同科的3个种均为常见种,χ2检验显示种对间整体负联结强于正联结。三者均为林冠食虫集团,食性和取食行为相似(附录1,附录2),但其PCC月联结频率负联结30%—50%之间,其正联结都是9%,种间负联结强于正联结(表2),这缘于有相似习性的山雀类与大山雀在共同分布地区内少见[36]。
(4)大山雀与环颈雉、山噪鹛、大斑啄木鸟的PCC月联结频率负联结均大于50%,正联结小于30%。仅环颈雉在3月份与大山雀表现出显著正联结,而其负联结月份均具有显著性,同时,联结指标显示负联结强度较大。环颈雉属于灌草丛营巢、地面取食集团,与大山雀生态位重叠度较低[37];山噪鹛属于灌草丛营巢、灌丛取食集团,大斑啄木鸟洞巢营巢、树干取食集团[16],两物种不仅在营巢和取食行为上与大山雀具有差异性,而且仅为林区落叶阔叶林的常见种[32],因而,导致环颈雉,大斑啄木鸟,山噪鹛与大山雀的遇见率低,从而导致它们负联结。
(5)大山雀与小嘴乌鸦、灰头绿啄木鸟的PCC月联结频率正负联结频率都低于30%,但负联结强于正联结,同时两物种遇见率较低(附录1)。这是由于大山雀常利用啄木鸟废巢来筑巢[38-39];灰头绿啄木鸟属于洞巢营巢、树干取食集团[40],小嘴乌鸦属于林冠枝干营巢、地面杂食取食集团[16]。
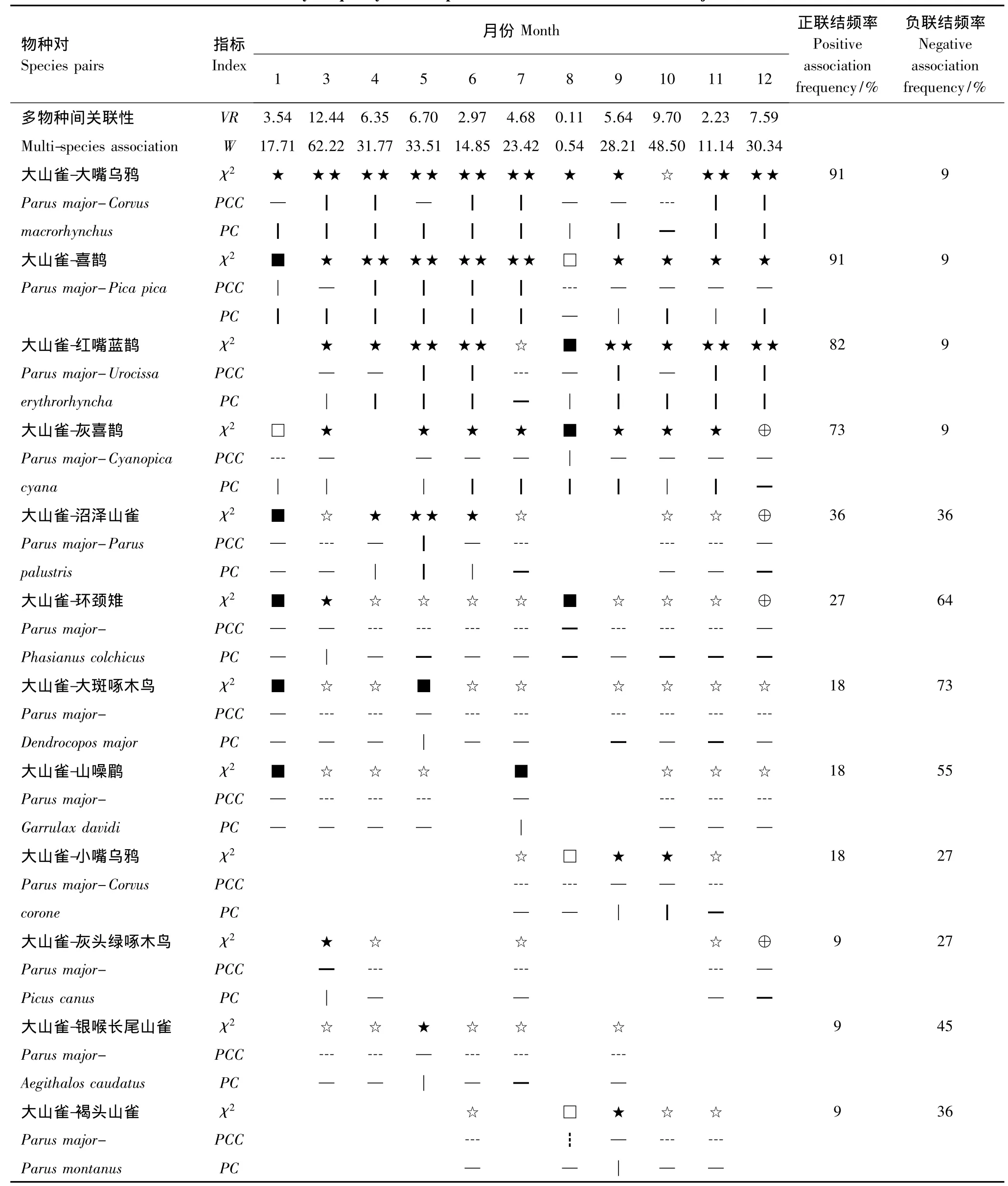
表2 大山雀与其他留鸟种间月联结频率Table 2 The monthly frequency of interspecific associations between Parus major and other residents
2.3.2 大山雀与夏侯鸟物种联结性分析
根据χ2检验、PCC、PC与PCC的月联结频率(表3),将大山雀与夏候鸟的联结划分成3个联结组,正联结组:①四声杜鹃(Cuculus micropterus);②正负均较弱的组为白眉姬鹟(Ficedula zanthopygia)、黄腹山雀(Parus venustulus);③负联结较强的组为黄腰柳莺、北红尾鸲(Phoenicurus auroreus)。

表3 大山雀与夏侯鸟种间月联结频率Table 3 The monthly frequency of interspecific associations between Parus major and summer migrants
(1)大山雀与四声杜鹃在5—7月份均呈显著正联结,共同出现百分率PC较高在0.5以上,驻留期PCC月联结频率正联结38%,由于四声杜鹃营巢属于巢寄生性,灰喜鹊的巢穴是四声杜鹃的常见寄居的巢穴[41],两物种生态位重叠值较高[26],从而导致两物种的正联结。
(2)白眉姬鹟、黄腹山雀调查中仅5—7月份才有发现,导致PCC月联结频率正负联结均小于30%,驻留期PCC月联结频率负联结25%,但整体负联结较强,这是由于白眉姬鹟属于灌丛取食集团(附录2),其在取食高度上白眉姬鹟在0—5 m的范围,大山雀大于15 m的范围[16];大山雀与黄腹山雀是同科物种,在筑巢和取食行为上均有相似性,但两物种存在垂直代替现象[36],呈现负联结。
(3)大山雀与黄腰柳莺、北红尾鸲PCC月联结频率负联结在30%—50%之间,正联结小于30%,负联结强于正联结,黄腰柳莺1、4、5、7—11月份均有出现,1月份为负联结,虽不具显著性,但其PCC绝对值较高,说明大山雀与黄腰柳莺的负联结较强,其在1、11月份出现可能是由于气候变暖,城市热岛效应使得一部分黄腰柳莺居留型发生变化,对其的探讨还有待研究,黄腰柳莺也为林冠层取食集团(附录2),驻留期PCC月联结频率负联结达到63%,与大山雀存在垂直代替现象[36],表明两物种竞争关系明显。大山雀与北红尾鸲多数月份呈负联结,其驻留期PCC月联结频率负联结较高,大山雀与北红尾鸲食性有部分重叠(附录1),虽然为林冠取食、洞巢集团(附录2),但驻留期PCC月联结频率负联结大于正联结,是由于北红尾鸲营巢喜于在人类的活动场所[42],西山地区靠近北京市区,为北红尾鸲选择理想巢址提供便利。
2.3.3 大山雀与冬侯鸟物种联结性分析
根据χ2检验、PCC、PC与PCC的月联结频率(表4),大山雀与冬侯鸟划分成2个联结组:①正负联结均较弱组为大山雀与棕眉山岩鹨(Prunella montanella);②负联结组为斑鸫(Turdus naumanni)、田鹀(Emberiza rustica)、小鹀(Emberiza pusilla)、黄眉鹀(Emberiza chrysophrys)。
(1)大山雀与岩鹨科棕眉山岩鹨的正负联结均较弱,驻留期PCC月联结频率正负联结分别为17%、33%,PC为0.24—0.25;其在1月份与大山雀表现为不显著正联结,且PC指标较小;3、12月份为显著负联结,棕眉山岩鹨为地面属于取食集团[43],与大山雀在习性上有较大差异。
(2)大山雀与鸫科的斑鸫、鹀科多种鸟类呈明显的负联结,驻留期PCC月联结频率负联结为17%—50%,正联结为0,PC为0.19—0.24;斑鸫属地面取食集团(附录2),鹀类属于地面取食集团[43],且与大山雀在习性上有较大差异。
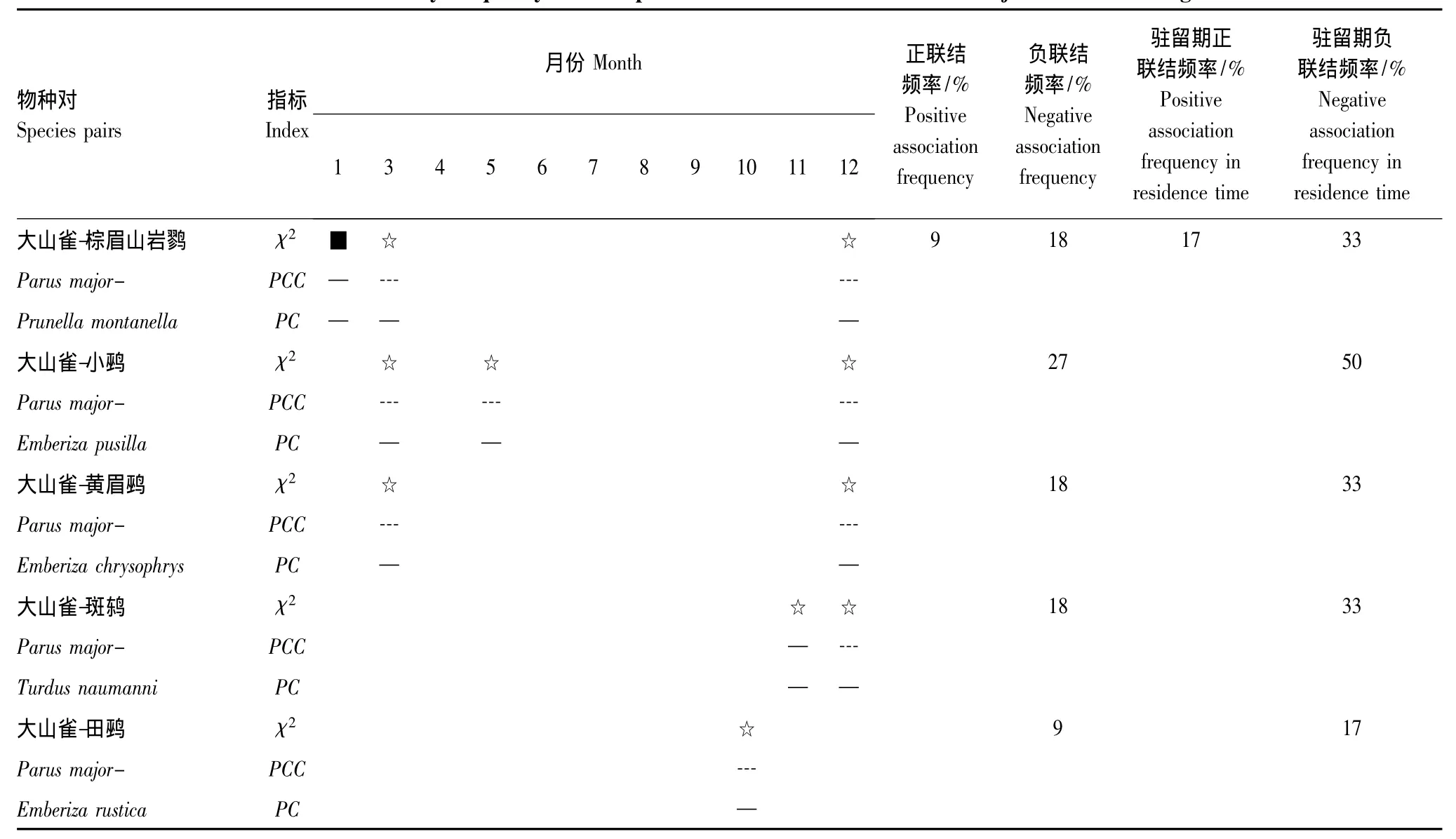
表4 大山雀与冬侯鸟种间月联结频率Table 4 The monthly frequency of interspecific associations between Parus major and winter migrants
2.3.4 大山雀与旅鸟物种联结性分析
与大山雀有显著正联结关系的旅鸟2种,根据χ2检验、PCC、PC指数、停留期PCC月联结频率(表5),可划分成2个联结组,①正负联结均较弱组为大山雀与褐柳莺;②负联结较强组为大山雀与黄眉柳莺。
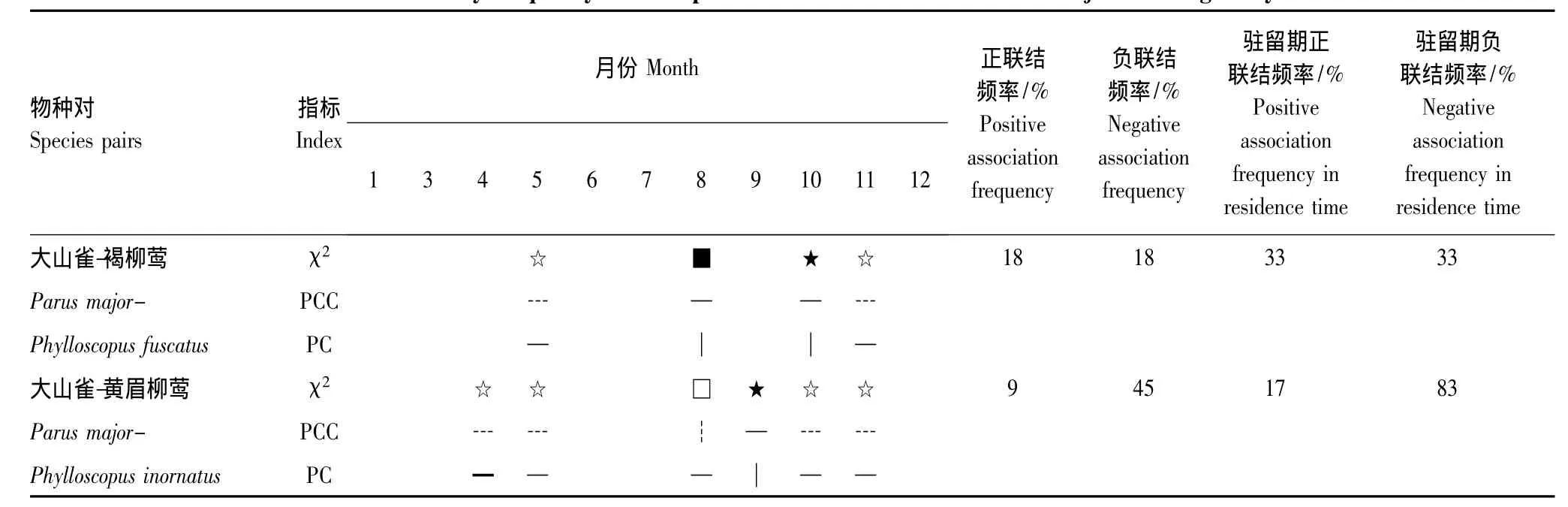
表5 大山雀与旅鸟种间月联结频率Table 5 The monthly frequency of interspecific associations between Parus major and migratory birds
(1)大山雀与褐柳莺正负联结均较弱,停留期PCC月联结频率正负联结相同,5月、11月为显著负联结,8月为不显著正联结、10月为显著正联结。虽然同为林冠食虫集团,由于其体重、嘴峰、跗跖都有明显差异,使得在植物群落停息的水平位置和取食数量、大小上也有差别,说明对资源的再次分割导致了这种联结关系。
(2)大山雀与黄眉柳莺负联结较强,4—5、8、10—11月PC指标均小于0.5,点相关系数PCC在8月份中为-0.61,停留期PCC月联结频率负联结83.3%,虽然2种同属林冠取食集团(附录2),但二者存在垂直代替现象[36]。
3 结论
3.1 鸟类群落种间联结度的测度
用固定样带法月连续全天定位调查,引入2×2列联表以日尺度的定性数据检验与测度联结度,与月尺度的联结频率结合起来,分析鸟类群落的种间联结是可行的,因而,鸟类群落的种间联结具有时间与空间尺度的双重属性。
3.2 以大山雀为“目标种”的月尺度种间联结
大山雀与留鸟、夏候鸟、冬候鸟、旅鸟等不同居留型的联结关系,随着种数的减少,显著联结种对、联结度、联结复杂程度呈递减趋势;留鸟与夏候鸟之间通过PCC月联结频率正负均较弱的山雀科联系着,夏候鸟与冬候鸟之间通过PCC月联结频率呈负联结的鸫科而联系着,夏候鸟与旅鸟之间通过PCC月联结频率呈负联结的莺科而联系着。
3.2.1 大山雀与留鸟的种间联结
大山雀与留鸟的联结关系可划分为正联结组、正负联结均等且较弱组、负联结组。大山雀与生态位有部分重叠的鸦科鸟类呈明显的正联结,如大嘴乌鸦、灰喜鹊、喜鹊、红嘴蓝鹊等,PCC月联结频率正联结为70%—90%,PC为0.25—0.98;大山雀与同科鸟类的由正负均等趋向于负联结,如沼泽山雀、银喉长尾山雀、褐头山雀等,PCC月联结频率正联结为9%—36%、负联结为36%—45%,PC为0.2—0.98;大山雀与体型较大环颈雉、啄木鸟、山噪鹛、小嘴乌鸦等鸟类呈负联结,PCC月联结频率正联结为9%—27%、负联结为27%—73%,PC 为 0.2—0.78。
3.2.2 大山雀与夏候鸟的种间联结
大山雀与夏候鸟的联结关系可划分为正联结组、正负联结均较弱组、负联结组。大山雀与杜鹃科四声杜鹃呈正联结,驻留期PCC月联结频率正联结为38%,PC为 0.59—0.78;大山雀与同科的黄腹山雀、鹟科的白眉姬鹟等的正负联结均较弱,驻留期PCC月联结频率正联结均为13%,负联结均为25%,PC为0.2—0.78;大山雀与莺科的黄腰柳莺、鸫科的北红尾鸲等呈负联结,驻留期PCC月联结频率正联结为9—27%,负联结频率较高均为 63%,PC 为 0.2—0.98。
3.2.3 大山雀与冬候鸟的种间联结
大山雀与岩鹨科棕眉山岩鹨的正负联结均较弱,驻留期PCC月联结频率正负联结分别为17%、33%,PC为0.24—0.25;大山雀与鸫科的斑鸫、鹀科多种鸟类呈明显的负联结,驻留期PCC月联结频率负联结为17—50%,正联结为 0,PC 为 0.19—0.24。
3.2.4 大山雀与旅鸟的种间联结
大山雀与柳莺科鸟类的月尺度联结由正负联结均较弱到明显的负联结,停留期PCC月联结频率,褐柳莺正负联结均为33%,黄眉柳莺负联结为83%;PC为0.2—0.59。
3.3 鸟类群落种间联结度的生态机制
植物群落的种间联结度的测度是空间结构与格局的表征,而鸟类群落的种间联结度不仅是鸟类群落时空结构与格局的表征,而且反映了鸟类群落的动态性。鸟类群落的种间联结对栖息的植物群落具有依赖性与关联性,只有将植物、鸟类、昆虫等组成的生物群落统一起来,才能更好地揭示其种间联结的生态机制,这有待深入研究。
[1]Whittaker R H.Ordination of Plant Communities.The Hague:W.Junk,1978.
[2]Zhang J T.Quantitative Ecology.Beijing:Science Press,2004:107-111.
[3]Kershaw K A,Looney JH H.Quantitative and Dynamic Plant Ecology.London:Edward Arnold Limited.1985:78-94.
[4]Shen Y Y,Li Y,Lu N.Competitive ability and interspecific relationship among four forage species.Acta Pratacultural Science,2002,11(3):8-11.
[5]Wang B S,Peng S L.Studies on the measuring techniques of interspecific association of lower-subtropical evergreen-broadleaved forests-I.the exploration and the revision on the measuring formulas of interspecific association.Acta Phytoecologica et Geobotanica Sinica,1985,9(4):274-285.
[6]Guo Z H,Zhuo Z D,Chen J,Wu M F.Interspecific association of trees in mixed evergreen and deciduous broadleaved forest in Lushan mountain.Acta Phytoecologica Sinica,1997,21(5):424-432.
[7]Fang F,Hu Y K,Zhang W,Gong Y M,Liu Y Y,Yang X J.Numerical analysis of inter-specific relationships in Alpine steppe community in Bayanbulak.Acta Ecologica Sinica,2012,32(6):1898-1907.
[8]Lou Y J,Zhao K Y.Analysis of interspecific associations of Carex lasiocarpa community in recent 30-year succession in Sanjiang Plain.Chinese Journal of Ecology,2008,27(4):509-513.
[9]Kang B,Liu SR,Cai D X,Wen Y G,Shi Z M,Guo W F,Zhu H G,Zhang G J,Liu L.Species composition and correlation of understorey woody plants in chinese fir plantation in the lower subtropical area.Acta Ecologica Sinica,2005,25(9):2173-2179.
[10]Chu G M,Pan C D,Wan M,Jin Y,Meng M,He JC.An analysis on interspecific association of dominant desert species in outer space of Karamay agricultural comprehensive developmental region.Journal of Xinjiang Agricultural University,2004,27(4):1-6.
[11]Hubalek Z.Coefficients of association and similarity,based on binary(presence-absence)data:an evaluation.Biological Reviews,1982,57(4):669-689.
[12]Zhang GM,Yang D R,Xu L,Peng Y Q,Lu Y.Species associations of fig wasps associated with Ficus racemosa in Xishuangbanna.Chinese Journal of Ecology,2003,22(4):20-26.
[13]Cui P,Deng W H.Review on the status and development of bird community research.Chinese Journal of Zoology,2007,42(4):149-158.
[14]De Greef R M,Tilghman N G,Anderson SH.Foraging guild of North American birds.Environmental Management,1985,9(6):493-536.
[15]Bi Z L,Zhang Z W,Song J.Preliminary study on the relationship between the bird community and vegetation structure at Dongling mountain of Beijing.Journal of Beijing Nornal University,2003,39(5):663-668.
[16]Fan X S,Hu D F,Chen H Z,Wang F,Su X.Study on blocstructure and econiche characteristics of bird communities in the farming areas in North China plain.Arid Zone Research,2008,25(4):544-549.
[17]Gao Y,Qian G Z.The niche analysis of bird community in evergreen broad-leaf forest in Tiantong Mountain,Zhejiang Province.Acta Ecologica Sinica,1987,7(1):73-82.
[18]Schluter D.A variance test for detecting species associations,with some example applications.Ecology,1984,65(3):998-1005.
[19]Yates F.Contingency tables involving small numbers and the χ2test.Supplement to the Journal of the Royal Statistical Society,1934,1(2):217-235.
[20]Pielou E C.Association tests versus homogeneity tests:their use in subdividing quadrats into groups.Vegetatio,1969,18(1/6):4-18.
[21]Pielou E C.An Introduction to Mathematical Ecology.New York:Wiley Interscience,1969:286-286.
[22]Yule G U.On the methods of measuring association between two attributes.Journal of the Royal Statistical Society,1912,75(6):579-642.
[23]Whittaker R H,Fairbanks C W.A study of plankton copepod communities in the Columbia Basin,southeastern Washington..Ecology,1958,39(1):46-65.
[24]Walter H,Ellenberg H.Einführung in die Phytologie.Eugen Ulmer,1956,4:136-136.
[25]Driver H E,Kroeber A L.Quantitative Expression of Cultural Relationships.California:The University of California Publications in American Archaeology and Ethnology,1932,31:211-256.
[26]Chen Y.Ecological Distribution,the Relationship of Plants and the Feeding Habits of Birds in An Evergreen Broadleaved Forest in Tiantong's 20hm2plot,Zhejiang[D].Shanghai:East China Normal University,2012.
[27]Morisita M.Measuring of interspecific association and similarity between communities.Memoirs of the Faculty of Science,Kyushu University,Series E(Biology),1959,3:65-80.
[28]Shangguan T L,Zhang F.Research on the pattern and associations between dominants of the vegetation in mian mountain,Shanxi province.Journal of Wuhan Botanical Research,1988,6(4):354-364.
[29]Guo X Y,Li Y H,Lin H P.Studies on the Inter-specific Association among main species of cyclocarya paliurus community in Niumulin natural reserve.Journal of Fujian College of Forestry,2011,21(2):181-185.
[30]Goodall D W.Sample similarity and species correlation//Wittaker P H,ed.Ordination and Classification of Communities.The Hagues:W.Junk,1972:105-154.
[31]Liu P P,Cheng J M.Study on plant interspecific association.Research of Soil and Water Conservation,2000,7(2):179-184.
[32]Fan Z J,Dong D Y,Zheng R,Wang M Z,Wang Q F,Guan W B.Avian community diversity in Platycladus orientalis ancient trees at the Jingfu Temple in Beiing.Journal of Beijing Forestry University,2013,35(5):46-55.
[33]Chen H P,Gao Z X,Li X M.Structure of forest bird communities of Zhanhe forest region in summer.Jourual of Northeast Forestry University,1992,20(6):49-55.
[34]Hou JH,Wu M L,Hu Y F,Zhang X Z,Hu Z T.A school study on the vertical space patterns of bird community in Mulan wei chang forest park.Journal of Agricultural University of Hebei,2000,23(1):78-81.
[35]Gao W,Xiang G Q,Feng H L,Deng Q X,Zhao H.On changes of foraging behavior of the tits of two species in secondary deciduous wood.Chinese Journal of Ecology,1996,15(5):1-5.
[36]Zheng G M.Ecological distribution of birds in Beijing and its vicinity during summer.Zoological Research,1984,5(1):29-40.
[37]Hou JH,Guo SB,Li J L.Structure of bird communities of Xitai,Xiaowutai mountains in summer.Journal of Hebei Forestry College,1994,9(4):332-338.
[38]Cai Q K.The Avifauna of Beijing.Beijing:Beijing Press,1988.
[39]Zhou T.Structure Processes and Determinants of Bird Community in the Secondary Broadleaf Forest[D].Changchun:Northeast Normal University,2011.
[40]Gao W,Xiang G Q,Shang JW,Feng H L.On the guild structure and its relationship of secondary forest bird community.Journal of Northeast Forestry University,1990,18(S3):114-125.
[41]Zheng Z X,Xian Y H,Guan G X.Fauna Sinica Aves,Vol.6.Beijing:Science Press,1991.
[42]Zhao Z J.Ecology of Daurian redstart in Changbai area and the importance of plant protection.Acta Zoologica Sinica,1975,21(3):272-287.
[43]Zheng G M.Ecological distribution of birds during the winter in peking & vicinity.Acta Zoologica Sinica,1962,14(3):321-336.
参考文献:
[2]张金屯.数量生态学.北京:科学出版社,2004:107-111.
[4]沈禹颖,李昀,陆妮.4种牧草种间竞争力和种间关系的研究.草业学报,2002,11(3):8-13.
[5]王伯荪,彭少麟.南亚热带常绿阔叶林种间联结测定技术研究——Ⅰ.种间联结测式的探讨与修正.植物生态学与地植物学丛刊,1985,9(4):274-285.
[6]郭志华,卓正大,陈洁,吴梅凤.庐山常绿阔叶,落叶阔叶、混交林乔木种群种间联结性研究.植物生态学报,1997,21(5):424-432.
[7]房飞,胡玉昆,张伟,公延明,柳妍妍,杨秀娟.高寒草原植物群落种间关系的数量分析.生态学报,2012,32(6):1898-1907.
[8]娄彦景,赵魁义.三江平原毛苔草群落近30年演替过程中的种间联结性分析.生态学杂志,2008,27(4):509-513.
[9]康冰,刘世荣,蔡道雄,温远光,史作民,郭文福,朱宏光,张广军,刘磊.南亚热带人工杉木林灌木层物种组成及主要木本种间联结性.生态学报,2005,25(9):2173-2179.
[10]楚光明,潘存德,万猛,晋瑜,蒙敏,何江成.克拉玛依农业综合开发区外围主要荒漠植物种间联结测定分析.新疆农业大学学报,2004,27(4):1-6.
[12]张光明,杨大荣,徐磊,彭艳琼,卢耀.西双版纳聚果榕榕果小蜂种间联结性研究.生态学杂志,2003,22(4):20-26.
[13] 崔鹏,邓文洪.鸟类群落研究进展.动物学杂志,2007,42(4):149-158.
[15]毕中霖,张正旺,宋杰.北京东灵山地区鸟类群落与栖息环境的关系研究.北京师范大学学报(自然科学版),2003,39(5):663-668.
[16]范喜顺,胡德夫,陈合志,王峰,苏鑫.华北平原耕作区鸟类群落的集团结构及生态位.干旱区研究,2008,25(4):544-549.
[17]高颖,钱国桢.天童常绿阔叶林中鸟类群落结构的空间生态位分析.生态学报,1987,7(1):73-82.
[26]陈宇.天童常绿阔叶林样地鸟类生态分布及鸟类与植物关系[D].上海:华东师范大学,2012.
[28]上官铁梁,张峰.山西绵山植被优势种群的分布格局与种间联结的研究.武汉植物学研究,1988,6(4):357-364.
[29]郭相亿,李裕红,林慧萍.牛姆林区青钱柳群落的主要种群种间关联特征.福建林学院学报,2001,21(2):181-185.
[31]刘萍萍,程积民.植物种间联结关系的研究.水土保持研究,2000,7(2):179-184.
[32]范宗骥,董大颖,郑然,王敏增,王奇峰,关文彬.北京静福寺侧柏古树林鸟类群落多样性研究.北京林业大学学报,2013,35(5):46-55.
[33]陈化鹏,高中信,李先敏.沾河林区夏季森林鸟类群落结构研究.东北林业大学学报,1992,20(6):49-55.
[34]侯建华,武明录,胡永富,张向忠,胡振天.木兰围场国家森林公园鸟类群落垂直空间格局研究.河北农业大学学报,2000,23(1):78-81.
[35]高玮,相桂权,冯贺林,邓秋香,赵虹.次生阔叶林中大山雀和沼泽山雀取食行为的研究.生态学杂志,1996,15(5):1-5.
[35]郑光美.北京及其附近地区夏季鸟类的生态分布.动物学研究,1984,5(1):29-40.
[36]郑光美.北京及其附近地区夏季鸟类的生态分布.动物学研究,1984,5(1):29-40.
[37]侯建华,郭书彬.小五台山西台夏季鸟类群落结构研究.河北林学院学报,1994,9(4):332-338.
[38]蔡其侃.北京鸟类志.北京:北京出版社,1988
[39]周彤.山地次生阔叶林鸟类群落的确定和结构过程[D].长春:东北师范大学,2011.
[40]高玮,相桂权,尚金城,冯贺林.山地次生林鸟类集团结构及关系的研究.东北林业大学学报,1990,18(S3):114-125.
[41]郑作新,冼耀华,关贯勋.中国动物志·鸟纲:第六卷,鸽形目·鹦形目·鹃形目·鸮形目.北京:科学出版社,1991.
[42]赵正阶.北红尾鴝在长白山地区的生态学及其在植物保护中的意义之研究.动物学报,1975,21(3):272-287.
[43]郑光美.北京及其附近地区冬季鸟类的生态分布.动物学报,1962,14(3):321-336.

附录1 北京静福寺鸟类调查名录Appendix1 Bird survey list at Jingfu Temple,Beijing

续表
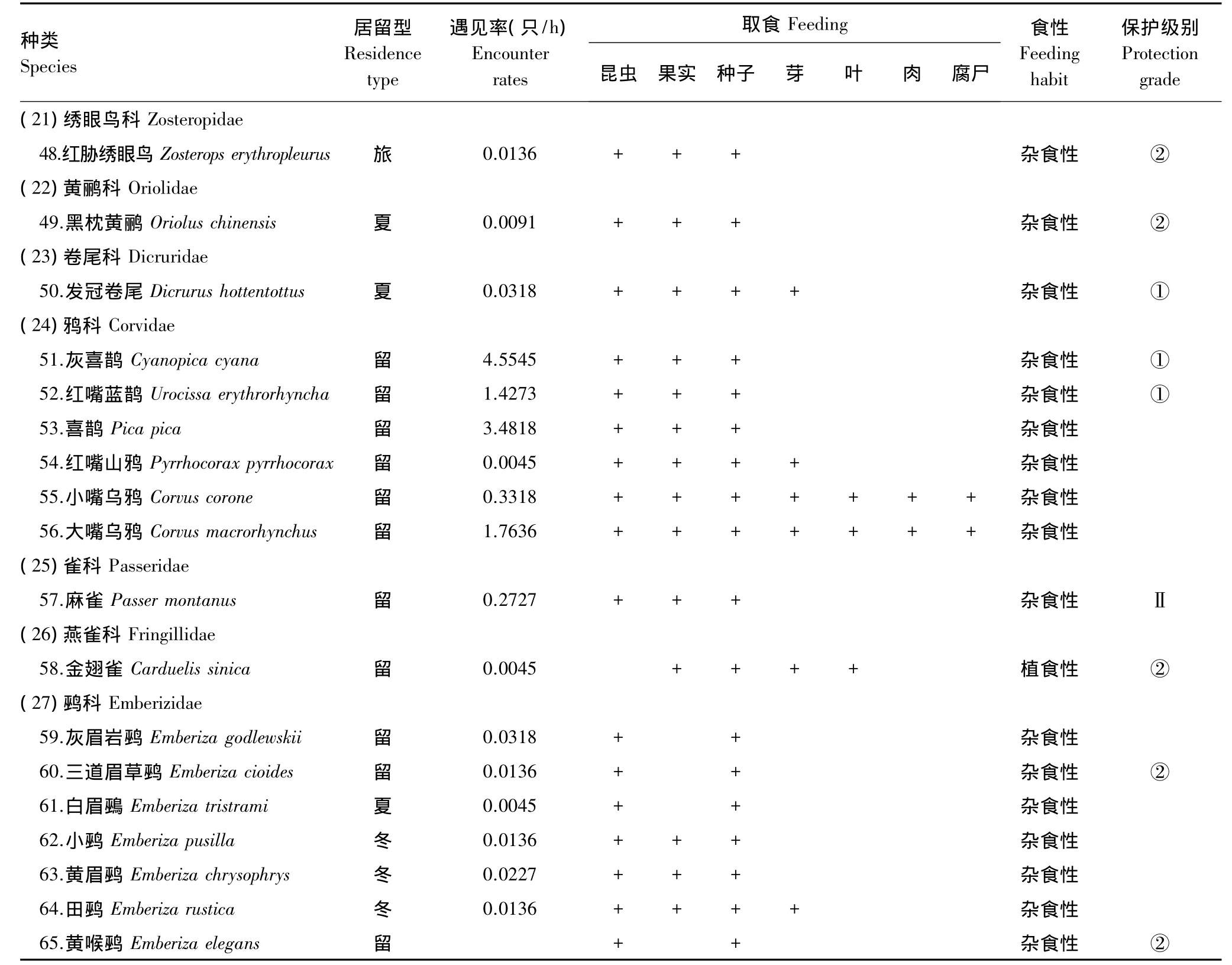
续表

附录2 鸟类营巢及取食集团划分Appendix 2 Divisions of the avian nesting and foraging guilds

续表

续表
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
小学阅读指南·高年级版(2021年9期)2021-10-09
孩子·小学版(2020年2期)2020-01-25
阅读(科学探秘)(2019年4期)2019-06-19
蔬菜(2018年5期)2018-05-17
现代营销·经营版(2017年5期)2017-06-21
杂草学报(2015年2期)2016-01-04
作文与考试·小学低年级版(2015年19期)2015-11-16
小主人报(2015年18期)2015-03-02
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
