
滇西北高原碧塔湖滨沼泽植物群落分布与演替
2013-12-21 04:59韩大勇杨永兴
生态学报 2013年7期
韩大勇,杨永兴,*,杨 杨
(1.同济大学环境科学与工程学院污染控制与资源化国家重点实验室,长江水环境教育部重点实验室,上海 200092;2.同济大学生命科学与技术学院,上海 200092)
植物群落总是沿环境梯度呈间断或连续分布,形成生态序列[1]。湿地植物群落沿水分梯度表现为典型的生态序列分布,并受局部地形等环境因素制约[2-3]。探讨生态序列的分布规律、形成机制和影响因素,对于理解湿地植物种类构成机制、群落演替规律、明确控制群落的关键生态因子乃至生态系统的发生、发展过程均具有重要意义[4-7]。迄今有关湿地群落生态序列已有大量报道,大多集中于滨海湿地和平原湿地,有关高原沼泽的报道相对较少[8-10],尤其有关喀斯特高原沼泽群落生态序列的研究报道更少。高原沼泽是一类具有特殊生态环境功能和保护价值的湿地生态系统,承接和调节着高原的冰雪融水、地表径流和河流水量,具有独特的水文和生态功能,对维持高海拔地区特殊生境和生物多样性具有重要意义[11-13]。但是,长期以来,高原沼泽特别是喀斯特高原沼泽一直是我国湿地基础研究的薄弱领域[14]。
碧塔海湖滨沼泽是我国高原沼泽的重要组成部分,处于横断山系腹地,自然地理上属青藏高原东南缘,为典型的喀斯特型高原沼泽,具有沼泽水文条件不稳定、沼泽发育受岩溶作用影响明显等独特的水文和地貌特征[15]。以往本区沼泽土壤学方面开展了较多研究[16-18],其他少量涉及生态健康评价[19]、植物多样性[20]、景观生态[21]等方面研究,但均未涉及湖滨带沼泽植物群落组成、分布与演替问题。本研究通过建立狭长型沟谷和宽广型沟谷两种类型湖滨沼泽植被样带,开展小尺度上的物种组成和群落演替序列及其驱动因子研究,并在此基础上初步分析了本区沼泽发育特征,这有助于丰富与完善中国沼泽学植物多样性与沼泽演替理论,为高原沼泽保护与退化沼泽生态恢复提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于云南省迪庆藏族自治州香格里拉县碧塔海自然保护区(99°39'—100°08'E,27°46'—27°57'N)的碧塔海片区(另一个为纳帕海片区),面积26869 hm2,平均海拔3200 m。碧塔海湖区呈东西走向,东西长约3000 m,南北宽300—1500 m,平均宽700 m,水域面积159 hm2,属于贫营养性湖泊。本区自然地理属青藏高原东南缘横断山脉三江纵向岭谷区东部。地质构造属滇西地槽褶皱系,古生界印支槽褶皱带,中甸剑川岩相带[16];地貌为横断山系古夷平面的断陷盆地,发育典型高原喀斯特岩溶地貌,主要地层为二迭统安山岩和石炭统灰岩[22];气候属亚热带季风气候区,但叠加了海拔高度与高原地貌作用,形成冷凉湿润的高原气候,主要特点为长冬无夏,春秋短促;气温年较差小,日较差大。年平均气温5.4℃,最热月(7月)均温13.2℃,最冷月(1月)均温-3.8 ℃。年平均降水量 617.6 mm,蒸发量 1013.9 mm,日照时数 2203 h[23],≥10 ℃ 积温1507.3℃[20];碧塔海属金沙江水系,湖区主要接受大气降水、湖周山地地表径流以及季节性河流补给,湖水由东部流出,经石灰岩溶洞泄入地下河,潜流后入洛吉河、尼汝河,最终汇入金沙江;主要湿地植被类型有草本沼泽、湖泊水生植被和湿生灌丛,植物区系以温带成分为主,兼有热带成分[20];主要土壤类型为泥炭土、沼泽土和草甸土[16]。
上述自然地理条件对沼泽发育极为有利,湖泊的南、西、北部湖滨沼泽发育比较广泛,在滇西北高原喀斯特地区具有很好的代表性与典型性。目前碧塔海为香格里拉县普达措国家公园的主要景区之一,木质栈道沿湖区北侧滨岸带穿过,湖滨沼泽存在轻微旅游干扰和极少量放牧行为,但是基本保持原生状态。
1.2 研究样带设置与植被调查
2011年7月植物生长旺季开展野外调查。此前曾于2010年7月对碧塔海湖滨沼泽进行过全面考察。本次调查在前次基础上再次对湖滨沼泽及其毗邻山地进行了详细、系统的考察,按两种沟谷类型建立湖滨沼泽研究样带,一类为狭长沟谷发育型,另一类为宽广沟谷发育型。其中狭长沟谷处于碧塔海西部湖滨带,谷口宽度约360 m,平均宽度约221 m,谷长(沿中轴方向)约860 m,长宽比约3.89∶1,总面积约15.48 hm2,为狭长沟谷型沼泽发育区;宽广沟谷处于北部湖滨带,谷口宽度约340 m,平均宽度约237 m,谷长约460 m,长宽比约1.43∶1,总面积约7.82 hm2,为宽广沟谷型沼泽发育区,前者坡降略小,后者坡降略大。两个沟谷地势均自顶端向沟谷出口湖滨倾斜,沟谷中部均发育入湖的沼泽性河流,与湖水一起补给沼泽。
根据沼泽发育典型性、代表性与完整性原则,分别在两类沟谷各设置一条研究样带,样带Ⅰ是位于西部的狭长型沟谷,长度约380 m;样带Ⅱ是位于北部的宽广型沟谷,长度约400 m。根据补给水源、地形和植物种类构成特点,沿沟谷最长中轴方向,每个样带按湖滨至沟谷顶部方向进一步依次划分样地:样带Ⅰ划分了7个样地,从样地1至样地7地形波动起伏。其中,样地1为湖滨浅水生境;样地2、4、7为沟谷洼地,均发育木里苔草(Carex muliensis)群落,样地2为湖滨沼泽,受稳定的湖水补给,样地4和7为沟谷陡坡下洼地发育的沟谷沼泽,主要接受地表径流与地下水补给;样地3、5、6为沟谷平亢地,相对毗邻区域地势较高,均发育木里苔草-杂类草群落,主要接受地表径流补给。样带Ⅱ划分了9个样地,从样地1至样地9地形略有起伏。其中,样地1—3为湖滨浅水生境;样地4为湖滨沼泽,接受径流和湖水补给;样地5、6、8和9均为沟谷平亢地,均发育了刘氏荸荠-杂类草(Eleocharis liouana-herb)群落,主要接受径流补给;样地7为沟谷洼地,发育大量草丘,以刘氏荸荠和木里苔草为主,主要接受地表径流和地下径流补给。调查时,依次记录每个样地地形、地貌、水文(积水状况、水位、补给水源)、沉积物等环境特征(表1)。每个样地设置3个面积1 m2植物群落样方,调查植物盖度、自然高度和密度。盖度采用目测法,自然高度为地面至植株顶端垂直高度,密度为单位样方内植株个体数。样带Ⅰ合计样方21个,样带Ⅱ合计样方27个。
1.3 数据处理
在样带尺度上分析了物种多度和频度分布,以反映两种类型沟谷沼泽群落的物种分布空间特征及总体环境条件差异。多度基于物种重要值,重要值=100×(相对盖度+相对高度+相对密度)/3,样带Ⅰ和样带Ⅱ的种重要值分别为21个和27个样方的均值,以平均值±标准差(mean±SD)形式示之。频度界定为样带内植物种出现的样方数,仅出现于1个或2个样方的种界定为稀有种。
为体现群落物种组成结构及环境特征的变化,物种多样性计算了α和β多样性。α多样性分为两个尺度:单位样方内的物种数,称物种丰富度SP,样地内的物种数,称总种数ST。其中,物种丰富度为3个样方内出现的物种数的平均值,以平均值±标准差的形式加以表现,在检验方差齐性前提下,用Duncan多重比较分析不同样地间物种丰富度的差异显著性;总物种数由样地内3个样方合并后获得。β多样性采用Whittaker指数βw[24],βw=ST/SP-1。采用Pearson相关[25]分析各多样性指数之间及其与水位之间的相关性。
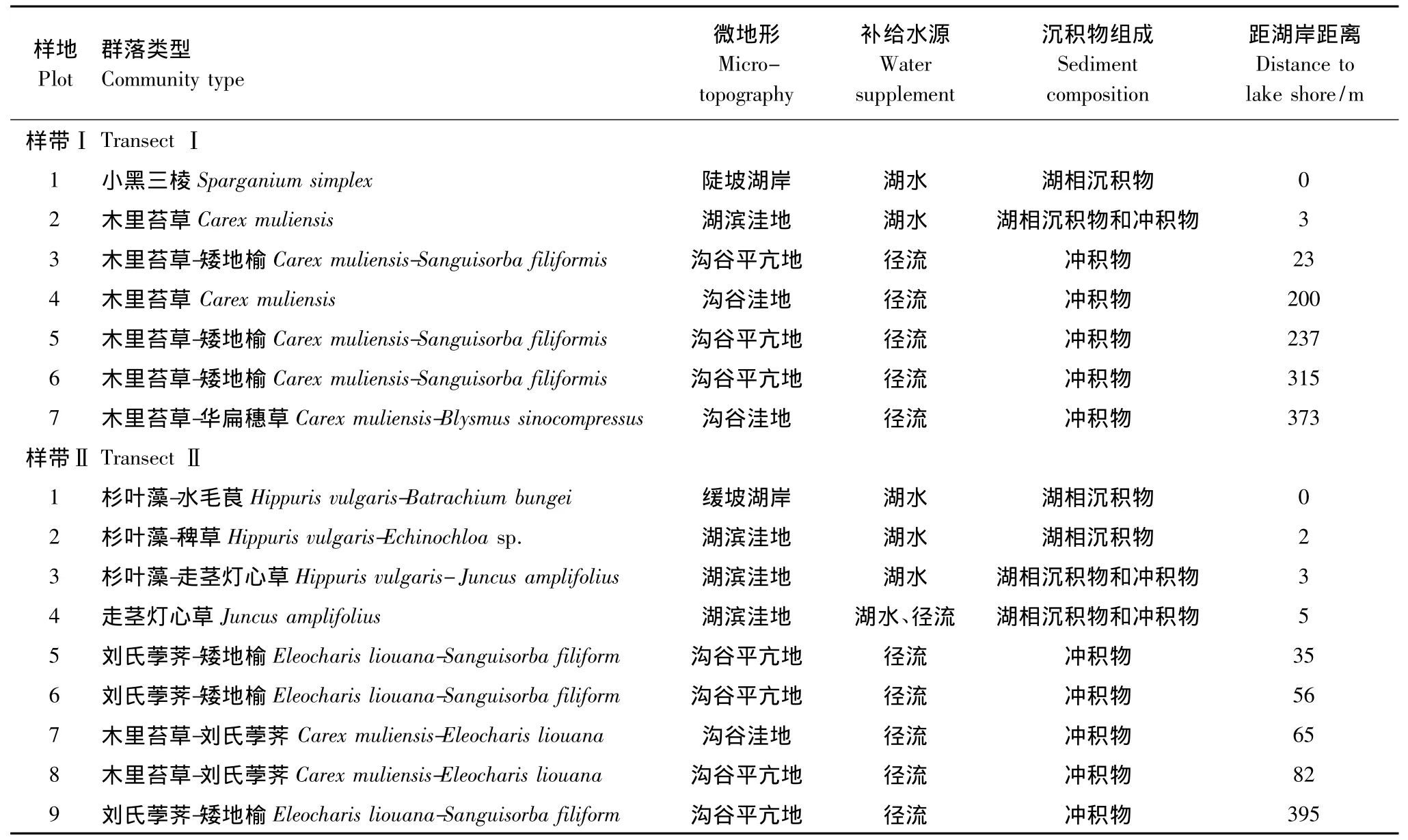
表1 滇西北高原碧塔海湖滨沼泽样地环境特征Table 1 Environment characteristics in different plot of the coastal swamp in the Bita lake wetland of the plateau region,Northwest Yunnan
应用DCA(Detrended Correspondence Analysis)排序分析群落种类组成和生态序列分布特征,排序基于样地的种重要值数据(3个样方均值),数据未进行转换。排序类型为间接梯度排序,参与排序的种重要值 >0.1,存在度 ≥33%,总计30种。
基本数据统计和物种多样性使用Microsoft Excel 2007软件完成;DCA排序使用Canoco 4.5 for Windows软件完成;Duncan多重比较和Pearson相关分析使用SPSS 16.0软件完成。所有统计分析显著水平均为P=0.05 或 0.01。
2 结果与分析
2.1 植物区系组成
两个样带总计出现43种植物(包括42种维管植物和1种苔藓植物),隶属于21科35属。科的物种丰富度以莎草科(Cyperaceae)和毛茛科(Ranunculaceae)最高,均包含6种(各占13.95%),其次是龙胆科(Gentianaceae),包含5种(占11.63%),禾本科(Poaceae)和眼子菜科(Potamogetonaceae)均包含3种(各占6.98%),灯心草科(Juncaceae)、蓼科(Polygonaceae)、伞形科(Umbelliferae)和玄参科(Scrophulariaceae)均包含2种(各占4.65%),其余11科均包含1种(各占2.33%)。属的物种丰富度以苔草属(Carex)、龙胆属(Gentiana)和眼子菜属(Potamogeton)最高,均包含3种,蓼属(Polygonum)包含2种,其余31属均为单种属(表2)。
样带Ⅰ出现28种,隶属于16科25属,重要值>8的有2种,为木里苔草和矮地榆(Sanguisorba filiformis),重要值分别为33.8和11.75;样带Ⅱ出现37种,隶属于20科31属,重要值>8的有4种,从大至小依次为刘氏荸荠>杉叶藻(Hippuris vulgaris)>走茎灯心草(Juncus amplifolius)>矮地榆(表2)。

表2 滇西北高原碧塔海湖滨沼泽植物群落区系组成及种重要值Table 2 Taxonomic composition and importance values(mean±SD)of plant species of the coastal swamp community in the Bita lake wetland of the plateau region,Northwest Yunnan
2.2 植物频度分布
样带Ⅰ的频度分布格局大致表现为L形,样带Ⅱ为U形。与样带Ⅰ相比,样带Ⅱ中出现于1个样方的种数明显减少,而出现于2个、3个和5个样方的种数明显增加,并且出现于14个样方的种数也明显高于样带Ⅰ的,意味着群落的总体均匀程度增加(图1)。但是,两个样带均以稀有种占大多数,分别为11种和10种,占各自总种数的39.29%和27.07%。
2.3 植物多样性
沿湖滨向沟谷顶部方向,按样地设置顺序,两个样带的物种多样性呈不规则变化。其中样带Ⅰ的单位样方物种丰富度、样地总种数、Whittaker指数均为双峰变化格局,最高值均出现在样地5,次高值出现在样地3,最低值出现在样地2;样地Ⅱ的各多样性指数变化格局不尽一致,单位样方物种丰富度和样地总种数的最低值出现在样地1,最高值出现在样地6,Whittaker指数最高值出现在样地7,最低值出现在样地6。样带Ⅰ各样地的水位排序为1>2≈4>6≈7≈5>3,样带Ⅱ各样地的水位排序为 1>2>3≈4>7>8≈6≈5>9(图2)。
进一步的相关分析表明,样带Ⅰ中的水位与物种丰富度具有显著正相关关系(P<0.05),但是与总种数和Whittaker指数无相关关系(P>0.05)。在各指数间,物种丰富度与总种数和Whittaker指数,以及总种数与Whittaker指数之间均存在显著正相关关系(P<0.05);样带Ⅱ中,水位与物种丰富度和总种数均呈显著正相关关系(P<0.05),但与 Whittaker指数无相关性(P>0.05)。在各指数间,仅物种丰富度与总种数存在显著正相关关系(P<0.05)。各多样性指数之间及其与水位之间相关性的差异体现出水分因子对植物群落物种组成的控制作用,以及局地条件的修正作用(表3)。

图1 滇西北高原碧塔海湖滨沼泽植物群落物种频度分布Fig.1 Frequency distribution of plant species of the coastal swamp community in the Bita lake wetland of the plateau region,Northwest Yunnan

图2 滇西北高原碧塔海湖滨沼泽植物群落物种多样性的变化Fig.2 Changes of plant diversity of the coastal swamp community in the Bita lake wetland of the plateau region,Northwest Yunnan
2.4 植物群落分布与演替
为了深入揭示碧塔海湖滨沼泽群落植物种类组成与分布规律,进行了间接DCA排序分析。结果表明:DCA第一排序轴较好地体现了沼泽地表水位变化对两个样带植物群落种类组成的影响和各样地的生态演替关系。在样带Ⅰ中,7个样地可以划分为3个类型:样地1属于挺水植物群落,主要种类有小黑三棱(Sparganium simplex)、竹叶眼子菜(Potamogeton malaianus)、蓖齿眼子菜(P.pectinatus)和轮叶狐尾藻(Myriophyllum verticillatum);样地2、4、7均为沼泽植物群落,均为木里苔草单优群落。其中样地2为湖滨沼泽,接受湖水补给,伴生杉叶藻、稗草(Echinochloa sp.),样地4、7为沟谷沼泽,接受沟谷地表径流与山麓地下水补给,伴生华扁穗草(Blysmus sinocompressus)、走茎灯心草和矮地榆;样地3、5、6地势较高,为轻度退化沼泽植物群落,虽然以木里苔草占优势,但是伴生大量的湿生植物,如矮地榆、走茎灯心草、刘氏荸荠、长茎毛茛(Ranunculus longicaulis)、珠芽蓼(Polygonum viviparum)、花葶驴蹄草(Caltha scaposa)、马先蒿(Pedicularis spp.)等,已经显现出沼泽退化过程,呈现向沼泽化草甸演替的趋势(图3)。

图3 滇西北高原碧塔海湖滨沼泽植物群落和种类组成DCA排序Fig.3 DCA ordination of the coastal swamp community in the Bita lake wetland of the plateau region,Northwest Yunnan
在样带Ⅱ中,9个样地可以划分为4个类型。样地1、2属于挺水植物群落,均为杉叶藻单优群落,伴生针蔺(Eleocharis valleculosa)、竹叶眼子菜、稗草;样地3、4属于挺水-沼泽植物群落过渡类型,以杉叶藻和走茎灯心草为主,伴生针蔺、稗草和乌拉苔草(Carex meyeriana);样地7属于沼泽植物群落,为木里苔草-杂类草群落,伴生华扁穗、长茎毛茛、矮地榆,接受地表径流补给;余下4个样地均为退化沼泽群落,其中,样地5、6、8属于轻度退化沼泽群落,为刘氏荸荠-木里苔草群落,伴生长茎毛茛、矮地榆、矮泽芹(Chamaesium paradoxum)等湿生植物,以及少量珠芽蓼、马先蒿;样地9属于中度退化沼泽,植物群落已经退化为沼泽化草甸植物群落,为刘氏荸荠-杂类草群落,地势相对较高,伴生较多的沼生和湿生种,诸如木里苔草、长茎毛茛、矮地榆、华丽龙胆(Gentiana sino-ornata)、珠芽蓼、马先蒿等(图3)。可见,沿水位变化梯度,两个样带植物群落均呈现出挺水植物群落→沼泽植物群落→沼泽化草甸植物群落的生态序列分布,但是各种群落类型在空间上随地形起伏与水分条件变化交错分布(图4)。
根据沿湖滨带沼泽野外详细、全面的考察、植物群落生态序列分布特征,结合地表水文条件及环境条件情况,得到相应的滇西北高原碧塔海湖滨沼泽植物群落的演替模式(图4)。样带Ⅰ位于狭长型沟谷,湖岸为陡坡湖岸,具有突变生境特征,植物群落也相应的由小黑三棱群落直接演替为木里苔草群落;样带Ⅱ位于宽广型沟谷,湖岸为缓坡湖岸,水深较样带Ⅰ浅,具有渐变生境特征,群落为连续分布,群落间边界不明显,存在过渡型群落。首先由杉叶藻群落演替为挺水的走茎灯心草群落,然后再演替为木里苔草-刘氏荸荠群落和刘氏荸荠-杂类草群落;但在沼泽群落综合演替模式上,沿水位梯度,木里苔草群落应首先演替为木里苔草-刘氏荸荠群落,然后为木里苔草-杂类草群落,最后为刘氏荸荠-杂类草群落。因此群落演替模式同生态序列分布规律一致:既存在水分主导下的渐进式演替,也存在微地形作用下的跳跃式演替,但演替方向都是一致的(图4)。

图4 滇西北高原碧塔海湖滨沼泽植物群落演替模式Fig.4 Succession mode of the coastal swamp community in the Bita lake wetland of the plateau region,Northwest Yunnan
3 讨论
碧塔海湖滨沼泽是喀斯特地貌上发育的高原沼泽,具有独特的水文和地貌特征。本研究中两个样带的植物群落均呈现出较完整的生态演替序列,但也在不同类型沟谷地理环境下演替序列存在差异,并在微地形作用下演替序列有所变异,呈现一定的群落复合体存在,这既反映了碧塔海湖滨沼泽植物群落的结构特征和演替过程的变化,也反映了水文、地貌在沼泽形成与发育过程中的控制作用。
3.1 物种频度与多度
频度和多度是两个重要的群落指标,综合反映出群落分布特征。Raunkiaer频度定律给出了植物群落频度分布的一般模式:A>B>C>=<D<E,即群落频度分布模式为双峰型,并以此来表征群落的同质性(或称均匀度)。但是该分布模式会受到样方大小和数量、物种数和个体数等多种因素的影响,因此Raunkiaer频度定律更适合于均质、稳定生境的群落[26-27],并且人为划分频度等级容易过高估计E级所包含的种数,因为E级的物种往往具有更多的个体,而A级的个体很少,具有不对称性[27-28]。对于本研究区域而言,两个研究样带均从湖滨挺水植物带至沟谷顶部,穿越了不同的生境类型,如水生生境、沟谷深凹洼地、沟谷缓坡平亢地、积水洼地生境等等。狭长型沟谷的高频度种(对应Raunkiaer的D级)包括矮地榆、矮泽芹和木里苔草,宽广型沟谷的高频度种(对应Raunkiaer的D级)仅矮泽芹一种,且两个样带均不存在E级种。因此,简单地将本研究结果与Raunkiaer频度定律进行类比并不具有可比性,而使用物种出现的样方数,能够较好的体现物种的分布范围及其生态幅和生态位的差异,并可在一定程度上克服由于人为分级而过高估计E级种数的不足[29]。参照物种多度分布结果进一步分析,在狭长型沟谷的样带Ⅰ中,重要值最高的木里苔草高达33.8,其他种重要值均较低,更倾向于单优群落特征:均匀度较低,聚集程度较高,而宽广型沟谷的样带Ⅱ中,各个种的重要值相差不大,更倾向于共优群落特征:均匀度高,聚集程度较低。实际上,与狭长型沟谷相比,宽广型沟谷面积小,集水面积也小,而且谷地坡降相对较大(无论横向或纵向),使沼泽补给水量与保存的水量均相对较少,木里苔草在群落中优势地位降低,而其他伴生的沼生或湿生植物多度增加,这是导致两种群落种类分布特征差异的外部环境原因。
3.2 植物多样性格局
物种多样性是植物群落对外界条件综合作用的反映,单位样方物种丰富度、总种数和Whittaker指数则共同反映出物种多样性的内涵,总种数的大小同时受到物种丰富度(样方内物种数)和Whittaker指数(样方间物种周转速率)的影响[30]。在样带Ⅰ中,总种数、物种丰富度和Whittaker指数表现出较一致的规律性变化,且各指数间均具有显著的相关性,表明样带Ⅰ的各样地内植物群落组成较一致,样方的物种丰富度对样地总种数的影响较大。样带Ⅱ的总种数与物种丰富度变化规律较一致,但Whittaker指数的变异性较大,物种丰富度较高的样地5和样地6的Whittaker指数较低,而物种丰富度较低的样地7的Whittaker指数反而较高,相关分析也表明Whittaker指数与物种丰富度和总种数之间均无显著相关性,甚至与物种丰富度表现出弱负相关。出现这种变异的原因可能与这几个样地的环境有关。样地5和6靠近沟谷边缘,地表坡度很小,水位低于地表20 cm以下,地表相对干燥,主要由刘氏荸荠和木里苔草构成,伴生大量双子叶杂类草,如矮地榆、车前(Plantago asiatica)、鳞叶龙胆(Gentiana squarrosa)、花葶驴蹄草、马先蒿、珠芽蓼等,种类分布较均匀;而样地7更靠近沟谷中部,受沟谷内小河河水水位季节变化的影响,沼泽水位波动较大,地表草丘微地貌发育,形成局部的起伏地形,丘上由刘氏荸荠组成,丘间有积水,由木里苔草构成,各群落片段呈复合分布,种类较少且异质性较高,由此导致β多样性出现较大的变异。因此,物种多样性的变化体现了两种类型沟谷中沼泽植物群落种类构成和水文地貌条件的差异。
3.3 植物群落分布、演替及其驱动因素
植物群落分布与演替几乎总是密不可分的,刘慎谔先生即将生态序列等同于空间演替[1],认识生态序列的分布规律是研究植物群落演替的前提之一。沿水分梯度,植物群落的生态序列分布是湖滨、河漫滩、盐沼等湿地植物群落分布的普遍格局。如三江平原的小兴凯湖、若尔盖高原的江错湖等[31]以及与碧塔海处于同一分布区的纳帕海湿地[32],均发育了典型的水生演替序列。本研究中,DCA排序结果提供了如下信息:尽管两个样带的群落种类组成和分布,均沿着水位变化梯度呈现出明显的生态序列,但在微地形的作用下,各种类型在空间上呈复合分布,如样带Ⅰ中的样地2、4、7为木里苔草群落,而样地3、5、6为木里苔草-杂类草群落,两种类型交错分布。其原因在于,样地2、4、7均为沟谷间洼地,均为汇水区,3个样地均发育了较好的木里苔草沼泽;样地3、5、6位于湖岸线以上,相对毗邻区域地势较高,尤其样地3虽然位于湖滨带下部,但是地势高出湖水面约20 cm,除洪水期外不能得到湖水补给,因此,除木里苔草外还生长大量杂类草种类。
对淡水沼泽植物群落分布研究发现,尽管水因子主导群落分布,但微地形的变化对群落分布也有重要影响[3]。对滨海盐沼植物群落分布研究也发现,除水位外,局部的微地形变化、碱斑发育以及由此导致的土壤水分条件的变化也会影响植物丰富度和分布[33-34]。本文的研究结果与上述研究基本一致。当然,植物群落的生态序列分布还受种间竞争、捕食作用等生物关系的修正,如互花米草入侵已显著地改变我国滨海盐沼原生植物群落分布格局[35],但在本研究中,不存在以上生物作用。
综合上述分析,沼泽水分补给类型、沼泽水位、微地貌等环境因子综合作用是影响碧塔海湖滨带植物群落分布和演替的主导因素。其一般分布规律为:挺水水生植物群落→沼泽植物群落→沼泽化草甸植物群落,但也存在由微地形引起的水分条件差异,生态序列分布受到干扰,形成复合群落。相应地群落演替模式为:水分主导下的渐进式演替和微地形作用下的跳跃式演替两种演替模式并存。这从植物群落物种频度分布和植物多样性格局也可以得到佐证。
3.4 植物群落演替与沼泽发育和退化
沼泽发育具有多途径的特点[36-37]。碧塔海湖滨沼泽发育主要由水体沼泽化与陆地沼泽化过程形成,水体沼泽化包括湖泊沼泽化、河流沼泽化、沟谷洼地小泡沼沼泽化;陆地沼泽化为草甸沼泽化。碧塔海湖滨带及其毗邻沟谷沼泽是经过上述诸沼泽化过程形成的复合沼泽体。本研究中两个样带群落的生态序列和复合分布格局较好的体现了这种沼泽发育特点。湖滨小黑三棱沼泽、杉叶藻沼泽为典型的湖泊沼泽化过程;分布于沟谷中部的木里苔草沼泽(样带Ⅰ中的样地4和7,样带Ⅱ中的样地7、8)是经由河流沼泽化过程与谷地洼地小泡沼沼泽化过程发育形成的,均发育泥炭层,甚至厚层泥炭层,泥炭质量很好。但部分样地泥炭层中存在坡麓冲积物,并且受冲积物影响,分布埋藏古泥炭层,存在着泥炭堆积过程中断,之后再次堆积发育的过程。沟谷边缘刘氏荸荠沼泽等(样带Ⅰ中的样地4、5、6以及样带Ⅱ中的样地5、6、9)土壤泥炭层发育不好,下覆典型的腐殖质层,为经过草甸化过程发育的沼泽。综上所述,碧塔海湖滨带沼泽水文补给条件相对稳定,泥炭堆积过程普遍,发育泥炭沼泽湿地。湖滨带沼泽是经过复合沼泽化过程发育的沼泽湿地,体现了高原断陷盆地湖滨沼泽形成与发育的典型模式。
沼泽的发育和退化分别是沼泽顺向演化与逆向演化的两种不同过程,沼泽发育过程也存在退化过程[38]。在两个样带中,均发育了刘氏荸荠-杂类草沼泽,这在样带Ⅱ宽广型沟谷的中、上部几个样地尤为普遍,表现出一定程度的退化。其中的样地5、6属于正在发育的沼泽,但已呈现退化的趋势。样地9则已退化为沼泽化草甸。目前碧塔海湖滨沼泽尽管存在轻微的旅游干扰,但尚未影响沼泽的形成与发育过程,而且有数据表明,近30年来,碧塔海湖区水位降低了约3 m[39]。推断水文条件的变化可能是沼泽退化的主要原因,碧塔海湖滨沼泽呈现退化的趋势是自然因素作用的结果,与狭长型沟谷相比,宽广型沟谷的局部区域沼泽退化趋势更为明显。因此,碧塔海湖滨沼泽的发育过程具有其脆弱性,严重受制于水文条件和局部地形,尤其对于只接受地表径流补给的沟谷沼泽,很容易因为湖泊水位下降,侵蚀基准面降低,沼泽地表发生排水过程,导致沼泽发育中断,出现沼泽退化。因此,不同水文条件下的沼泽对外界干扰具有不同的抵抗力,保持水文地貌条件的稳定,是人为调控沼泽发育和应对沼泽退化的对策之一。在喀斯特地区高原沼泽管理和利用中,应注意保持水文地貌条件的稳定,防止人为因素导致沼泽退化。
4 结论
(1)两种沟谷类型的湖滨沼泽群落植物多样性均与水位呈显著正相关关系,体现出水分因子对群落种类组成的控制作用,受沟谷地形影响,狭长型沟谷的沼泽群落总体均匀程度高于宽广型沟谷。
(2)碧塔海湖滨沼泽群落生态序列分布与湖岸特点和沟谷类型有关:狭长型沟谷具有陡坡湖岸,沼泽群落生态序列倾向于间断分布,宽广型沟谷具有缓坡湖岸,沼泽群落生态序列倾向于连续分布。
(3)碧塔海湖滨沼泽植物群落分布既存在水分主导的生态序列:挺水水生植物群落→沼泽植物群落→沼泽化草甸植物群落,也存在由微地形引起的水分条件再分配导致的群落复合分布。相应的植物群落演替特点为渐变式演替和跳跃式演替两种模式并存,但演替方向完全一致。
(4)碧塔海湖滨沼泽发育主要由湖泊沼泽化、河流沼泽化、沟谷洼地小泡沼沼泽化和草甸沼泽化等沼泽化过程形成的复合沼泽体。碧塔海湖滨沼泽发育过程具有退化倾向,与狭长型沟谷相比,宽广型沟谷的局部区域退化趋势更为明显。
[1]Zhu T C,Zhong Z C,Li JD.Plant Ecology.Beijing:Higher Education Press,1988:203-216.
[2]Crain CM,Silliman B R,Bertness SL,Bertness M D.Physical and biotic drivers of plant distribution acrossestuarinesalinity gradients.Ecology,2004,85(9):2539-2549.
[3]Raulings E J,Morris K,Roache M C,Boon P I.The importance of water regimes operating at small spatial scales for the diversity and structure of wetland vegetation.Freshwater Biology,2010,55(3):701-715.
[4]Casanova M T,Brock M A.How do depth,duration and frequency of flooding influence the establishment of wetland plant communities?Plant Ecology,2000,147(2):237-250.
[5]Xiao D R,Yuan H,Tian K,Yang Y.Distribution patterns and changes of aquatic communities in Lashihai Plateau Wetland after impoundment by damming.Acta Ecologica Sinica,2012,32(3):815-822.
[6]Pennings SC,Grant M B,Bertness M D.Plant zonation in low-latitude salt marshes:disentangling the roles of flooding,salinity and competition.Journal of Ecology,2005,93(1):159-167.
[7]Yao C,Wan SW,Sun DL,Qin P.Ecological mechanisms of vegetation succession of coastal wetland in Yancheng Nature Reserve.Acta Ecologica Sinica,2009,29(5):2203-2210.
[8]Li K,Yang Y X,Yang Y,Han D Y,Yang Y M,Tian K,Zhang CB.Research progress of the degradation and restoration of the plateau wetlands in China.Journal of Anhui Agricultural Science,2011,39(11):6714-6716,6719-6719.
[9]Yang Y X.Ecological environment deterioration,mire degeneration and their formation mechanism in the Zoige Plateau.Journal of Mountain Science,1999,17(4):318-323.
[10]Han D Y,Yang Y X,Yang Y,Li K.Species composition and succession of swamp vegetation along grazing gradients in the Zoige plateau,China.Acta Ecologica Sinica,2011,31(20):5946-5955.
[11]Yang Y X.Main characteristics,progress and prospect of international wetland science research.Progress in Geography,2002,21(2):111-120.
[12]Yang Y X.The 21st century hot point and forward position field of international wetland research from Quebec 2000-Millennium Wetland Event.Scientia Geographica Sinica,2002,22(2):150-155.
[13]Yang Y X.New knowledge on the progress of international wetland science research and priority field and prospect of Chinese wetland science research.Advance in Earth Sciences,2002,17(4):508-514.
[14]Yang Y X,Yang Y,Liu C E.The basic academic issue of wetland and wetland science and ecological and environmental function of wetland ecosystem.Landscape Architecture China,2009,3(1):14-17.
[15]Tian K,Mo JF,Lu M,Chang F L,Yang Y X.Human disturbances on the ecological environment degradation of Napahai wetland in the upstream of Yangtze river.Resources and Environment in the Yangtze Basin,2004,13(3):292-295.
[16]Tian K.Mechanism and Process of Soil Degradation in Napahai Wetland on Yunnan Plateau[D].Changchun:Northeast Institute of Geography and Agroecology,Chinese Academy of Sciences,2004.
[17]Tian K,Chang F L,Lu M,Mo JF,Yang Y X.Impacts of human disturbances on organic carbon and nitrogen in Napahai wetlands,Northwest Yunnan.Acta Pedologica Sinica,2004,41(5):681-686.
[18]Bei R T,Chen H Q,Tian K.Comparable research of soil nutrient in the Bitahai Nature Reserve.Journal of Southwest Forestry College,2003,23(2):61-64.
[19]Wang J L,Wang P,Lu F,Yang G H.Impact of tourist activities on the environment in Bitahai Lake wetland ecotourism scenic spot.Progress in Geography,2004,23(5):101-108.
[20]Yin WY,Du F,Zhang D C.A study on the rare and precious plant species in the Bitahai Nature Reserve.Journal of Southwest Forestry College,2004,24(1):20-22,30-30.
[21]Li SH,Wang JL,Chen H F,Chen Y.Research on spatial structure of Bitahai Lake Nature Reserve.Yunnan Environmental Science,2005,24(4):10-13.
[22]Wang Y H,Zhong L S,Yang GH.Discussion on exploitation of the scientific exploration tour in the Bita Lake Natural Reserve.Chinese Journal of Ecology,2000,19(2):64-67.
[23]Li Q,Wu Z L,Liu L L,Xu N,Yang X D.Grassland insect community diversity in Tibetan region of northwestern Yunnan under different management patterns.Chinese Journal of Ecology,2006,25(11):1375-1379.
[24]Whittaker R H.Evolution and measurement of species diversity.Taxon,1972,21(2/3):213-251.
[25]Lu W D.Statistical Analysis Using SPSS(4th ed).Beijing:Publishing House of Electronics Industry,2010:292-317.
[26]Williams C B.The application of the logarithmic series to the frequency of occurrence of plant species in quadrats.The Journal of Ecology,1950,38(1):107-138.
[27]McIntosh R P.Raunkiaer's“law of frequency”.Ecology,1962,43(3):533-535.
[28]Gleason H A.The significance of Raunkiaer's law of frequency.Ecology,1929,10(4):406-408.
[29]Hanski I.Dynamics of regional distribution:the core and satellite species hypothesis.Oikos,1982,38(2):210-221.
[30]Han D Y,Li H Y,Yang Y F.β-diversity patterns of plant in fragmented habitat in a degenerated meadow in Songnen Plain,China.Chinese Geographical Science,2009,19(4):375-381.
[31]Lang H Q,Zu W C,Jin SR.Mire in China.Ji'nan:Shandong Science and Technology Press,1983:122-128,168-174.
[32]Xiao D R,Tian K,Yuan H,Yang Y M,Li N Y,Xu SG.The distribution patterns and changes of aquatic plant communities in Napahai Wetland in northwestern Yunnan Plateau,China.Acta Ecologica Sinica,2006,26(11):3264-3630.
[33]Ewanchuk P J,Bertness M D.Structure and organization of a northern New England salt marsh plant community.Journal of Ecology,2004,92(1):72-85.
[34]Wang C H,Lu M,Yang B,Yang Q,Zhang X D,Hara T,Li B.Effects of environmental gradients on the performances of four dominant plants in a Chinese saltmarsh:implications for plant zonation.Ecological Research,2010,25(2):347-358.
[35]Chen Z Y,Li B,Chen J K.Ecological consequences and management of Spartina spp.invasions in coastal ecosystems.Chinese Biodiversity,2004,2(2):280-289.
[36]Huang X C.An approach to distribution and development law of mire.Scientia Geographica Sinica,1982,2(3):193-201.
[37]Yang Y X,Wang SY.Study on mire development and paleoenvironment change since 8.0 ka B.P.in the Northern part of the Sangjiang plain.Scientia Geographica Sinica,2003,23(1):32-38.
[38]Yang Y X.Study on the relationship between mire development and palaeogeographical environment changes since the late period of the late pleistocene in the Sanjiang plain.Oceanologia et Limnologia Sinica,1990,21(1):27-38.
[39]Guo J X,Wang Z Y,Bai B,Lu Y B.Climatic changes of the three-river-area and its effect on ecological environment in Yunnan.Yunnan Geographic Environment Research,2006,18(2):48-52.
参考文献:
[1]祝廷成,钟章成,李建东.植物生态学.北京:高等教育出版社,1988:203-216.
[5]肖德荣,袁华,田昆,杨杨.筑坝扩容下高原湿地拉市海植物群落分布格局及其变化.生态学报,2012,32(3):815-822.
[7]姚成,万树文,孙东林,钦佩.盐城自然保护区海滨湿地植被演替的生态机制.生态学报,2009,29(5):2203-2210.
[8]李珂,杨永兴,杨杨,韩大勇,杨宇明,田昆,张昌兵.中国高原湿地退化与恢复研究进展.安徽农业科学,2011,39(11):6714-6716,6719-6719.
[9]杨永兴.若尔盖高原生态环境恶化与沼泽退化及其形成机制.山地学报,1999,17(4):318-323.
[10]韩大勇,杨永兴,杨杨,李珂.放牧干扰下若尔盖高原沼泽湿地植被种类组成及演替模式.生态学报,2011,31(20):5946-5955.
[11]杨永兴.国际湿地科学研究的主要特点、进展与展望.地理科学进展,2002,21(2):111-120.
[12]杨永兴.从魁北克2000-世纪湿地大事件活动看21世纪国际湿地科学研究的热点与前沿.地理科学,2002,22(2):150-155.
[13]杨永兴.国际湿地科学研究进展和中国湿地科学研究优先领域与展望.地球科学进展,2002,17(4):508-514.
[14]杨永兴,杨杨,刘长娥.湿地与湿地科学基本理论问题与湿地生态系统的生态、环境功能.景观设计学,2009,3(1):14-17.
[15]田昆,莫剑锋,陆梅,常凤来,杨永兴.人为活动干扰对纳帕海湿地环境影响的研究.长江流域资源与环境,2004,13(3):292-295.
[16]田昆.云南纳帕海高原湿地土壤退化过程及驱动机制[D].长春:中国科学院东北地理与农业生态研究所,2004.
[17]田昆,常凤来,陆梅,莫剑锋,杨永兴.人为活动对云南纳帕海湿地土壤碳氮变化的影响.土壤学报,2004,41(5):681-686.
[18]贝荣塔,陈慧泉,田昆.碧塔海自然保护区土壤养分研究.西南林学院学报,2003,23(2):61-64.
[19]王金亮,王平,鲁芬,杨桂华.碧塔海景区旅游活动对湿地生态环境影响研究.地理科学进展,2004,23(5):101-108.
[20]尹五元,杜凡,张大才.碧塔海自然保护区珍稀保护植物研究.西南林学院学报,2004,24(1):20-22,30-30.
[21]李石华,王金亮,陈华芳,陈姚.碧塔海自然保护区景观空间格局研究.云南环境科学,2005,24(4):10-13.
[22]王跃华,钟林生,杨桂华.碧塔海自然保护区科考旅游开发探讨.生态学杂志,2000,19(2):64-67.
[23]李青,吴兆录,刘玲玲,徐宁,杨效东.滇西北藏区草地管理方式对草地昆虫群落多样性的影响.生态学杂志,2006,25(11):1375-1379.
[25]卢纹岱.SPSS统计分析 (第四版).北京:电子工业出版社,2010:292-317.
[31]郎慧卿,祖文辰,金树仁.中国沼泽.济南:山东科学技术出版社,1983:122-128,168-174.
[32]肖德荣,田昆,袁华,杨宇明,李宁云,徐守国.高原湿地纳帕海水生植物群落分布格局及变化.生态学报,2006,26(11):3264-3630.
[35]陈中义,李博,陈家宽.米草属植物入侵的生态后果及管理对策.生物多样性,2004,2(2):280-289.
[36]黄锡畴.试论沼泽的分布和发育规律.地理科学,1982,2(3):193-201.
[37]杨永兴,王世岩.8.0ka B.P.以来三江平原北部沼泽发育和古环境演变研究.地理科学,2003,23(1):32-38.
[38]杨永兴.三江平原沼泽发育与晚更新世末期以来古地理环境演变的研究.海洋与湖沼,1990,21(1):27-38.
[39]郭菊馨,王自英,白波,鲁亚斌.云南三江并流地区气候变化及其对生态环境的影响.云南地理环境研究,2006,18(2):48-52.
猜你喜欢
中国野生植物资源(2022年12期)2023-01-09
生态学报(2022年19期)2022-10-27
江苏安全生产(2022年5期)2022-06-16
作文成功之路·小学版(2020年5期)2020-06-11
山东工业技术(2019年16期)2019-07-19
西南农业学报(2018年6期)2018-08-04
贵州科学(2016年5期)2016-11-29
中国水土保持科学(2016年3期)2016-07-19
中国地质灾害与防治学报(2015年2期)2015-03-08
中国水利(2015年1期)2015-02-28
