
不同潮带红海榄群落胎生繁殖体空间分布格局分析
2013-12-27 05:32黄剑坚李际平刘素青韩维栋
中南林业科技大学学报 2013年3期
黄剑坚,李际平,刘素青,韩维栋
(1. 广东海洋大学 a.寸金学院,b.农学院,广东 湛江524094;2.中南林业科技大学,湖南 长沙 410004)
不同潮带红海榄群落胎生繁殖体空间分布格局分析
黄剑坚1a,李际平2,刘素青1b,韩维栋1b
(1. 广东海洋大学 a.寸金学院,b.农学院,广东 湛江524094;2.中南林业科技大学,湖南 长沙 410004)
在广东省湛江市高桥不同潮带不同红海榄群落展开胎生繁殖体空间分布格局调查。分析比较胎生繁殖体在的上、中、下等不同冠层,东、南、西、北等不同方位,以及面潮、背潮两部分等的8种聚集度指标(I、C、M*、CA…).结果表明,高桥内带红海榄胎生繁殖体主要分布在上层冠层、东方位和背潮部分;中带红海榄胎生繁殖体主要分布在中层冠层,在各个方位以及面潮和背潮两部分的分布无显著性区别;红海榄胎生繁殖体的空间分布格局皆为聚集分布。结合研究结果,提出在造林困难的潮带可进行红海榄胎生繁殖体的合理密集种植,提高胎生繁殖体的环境抵御能力,从而提高存活率。
红树林;红海榄;胎生繁殖体;分布格局;广东省湛江市
红 海 榄Rhizophora stylosa属 红 树 科Rhizophoraceae的红树植物,生长于热带、亚热带海岸潮间带中,为常绿灌木或小乔木,支柱根极为发达,果实倒圆锥状卵形,下垂,种子在果实离开母树前发芽,具有特殊的“胎生繁殖”方式,主要分布于广西、香港、广东和海南等地,是沿海红树林生态恢复的优选树种[1-2]。空间分布格局是指生物个体在与环境相互作用中所形成的,种群所在生境内个体的空间配置,是种群自身生物
学特性与外界因素综合作用的产物[3-4]。红树林种群以自身特有的生态学和形态学属性,在与各环境因子的相互作用下,形成一个相对稳定的水平空间分布格局。有关红树林分布格局亦有专文报道,主要从群落角度得出每个种群的分布格局[5-8]。并且亦有研究指出红树林种群的分布格局主要受自身的生物学特征和微地形影响,呈随机或集群分布[8]。目前,有关红海榄胎生繁殖体空间分布格局至今未见报道。分析红海榄胎生繁殖体的空间格局不仅可以了解胎生繁殖体的分布情况与环境之间的相互关系,并且有助于揭示林下及林窗红海榄更新幼苗空间分布格局形成的主要原因,对阐明红海榄群落的稳定、演替及恢复机理有重要的意义。本研究以廉江高桥红树林保护区的红海榄天然林的成熟胎生繁殖体为研究对象,描述在不同潮带、群落、冠层、方位等分布格局状况。
1 研究区概况
广东省湛江市廉江高桥红树林保护小区属于全日潮区,地处广东、广西交界的北部湾英罗港,东侧卖皂海,西侧官寨海,为湛江红树林国家级自然保护区的核心区之一。高桥红树林保护小区保护总面积2 249.4 hm2,有红树林1 361.6 hm2,可划分为44个小班(小班数为当地林业部门的林地区划),是全国面积最大红树林保护区[3]。该小区内主要有白骨壤Avicennia marina、桐花树Aegiceras corniculatum、红海榄Rhizophora stylosa、秋茄Kandelia obovata、木榄Bruguiera gymnorrhiza等主要红树植物,有海檬果Cerbera manghas、苦郎树Clerodendurm inerme、黄槿Hibiscus tiliaceus、杨叶肖槿Thespesia populnea等主要半红树植物。
2 研究方法
2.1 调查方法
2011年7月17号至23号于广东省湛江市廉江高桥红树林保护小区的内带和中带,共调查4个红海榄优势种样地,内带纯林和内带混交林(红海榄+木榄混交)立地条件相同,中带纯林与混交林(红海榄+木榄+桐花树+秋茄混交)林立地条件相同。每个样地随机抽取30株红海榄调查木,把每株调查木的冠层平均分为上、中、下三层,记录每层胎生繁殖体的数量和整株红海榄的胎生繁殖体数量;再对每株样木以主干为中心,以东、南、西、北四个方向平均分为四层,然后按面潮和背潮将树冠分成两部分。内带纯林树高(5.2±0.5) m,混交林树高(5.4±0.4) m;中带纯林树高(2.7±0.3) m,混交林树高(2.7±0.2) m。
2.2 分析方法
2.2.1 聚集度指标分析法
聚集度指标主要采用包括扩散系数C、丛生指数I、Cassie.R.M.指标CA、负二项参数K、平均拥挤度m*、聚块性指数指标m*/,详细计算方法见文献[4,9-11]。
2.2.2 回归模型分析法
2.3 数据处理分析
采用Spss16.0进行数据分析
3 结果与分析
3.1 聚集度指标计算结果
3.1.1 内带和中带红海榄群落胎生繁殖体聚集度比较分析
从表1可得,扩散系数C(12.24~31.72)>1;丛生指数I(11.24~30.72)>1;负二项参数 K(4.29~ 6.32)> 1; Cassie.R.M.指标CA(0.16~0.23)>0;每项平均拥挤度m*>;m*/(1.10~1.23)>1。因此,内带纯林、内带混交林(红海榄+木榄混交)、中带纯林、中带混交林(红海榄+木榄+桐花树+秋茄混交)的胎生繁殖体分布格局呈聚集分布。4个样地胎生繁殖体聚集强度分析可得:中带红海榄+木榄+桐花树+秋茄混交林>内带红海榄+木榄混交林>中带纯林>外带纯林。

表1 内带和中带红海榄群落聚集度比较分析Table 1 Comparative and analysis of R. stylosa community viviparous propagules aggregation degree in inner tide and middle tide
3.1.2 内带和中带红海榄群落不同冠层胎生繁殖体分布格局比较分析
内带和中带红海榄群落不同冠层胎生繁殖体的(见表2)扩散系数皆C>1;丛生指数I除了内带纯林下层和内带木榄+红海榄混交林下层外,其它皆>1;负二项参数K除了内带纯林下层外,其它皆>1; Cassie.R.M.指标CA>0;每项平均拥挤度m*>;m*/指标皆>1。因此,内带和中带红海榄群落不同冠层胎生繁殖体呈聚集分布。

表2 内带和中带红海榄群落不同冠层胎生繁殖体聚集度分析Table 2 Aggregation degree analysis of R. stylosa community viviparous propagules of different canopys in inner tide and middle tide
从图1可见,红海榄胎生繁殖体主要集中在中上层,占86%~91%以上,其中内带纯林和内带木榄+红海榄混交林的分布顺序是上>中>下,中带纯林和中带红海榄+木榄+桐花树+秋茄混交林的分布顺序是中> 上>下。通过多重比较分析(LSD)分析可得:内带纯林和内带木榄+红海榄混交林的上、中、下三层胎生繁殖体分布呈显著性区别;中带纯林和中带红海榄+木榄+桐花树+秋茄混交林的上中层无显著性区别,中下层和上下层呈显著性区别。
3.1.3 内带和中带红海榄群落不同方位胎生繁殖体分布格局比较分析
根据各参数判定标准可知(见表3),胎生繁殖体的扩散系数皆C>1;丛生指数I皆>1;负二项参数K除了内带纯林北方位,其它皆>1;Cassie.R.M.指标CA>0;每项平均拥挤度m*>;m*/指标皆>1。因此,内带和中带不同红海榄群落不同方位胎生繁殖体呈聚集分布。内带红海榄胎生繁殖体(见图2)主要分布在东边,中带主要分布在南边。通过多重比较分析
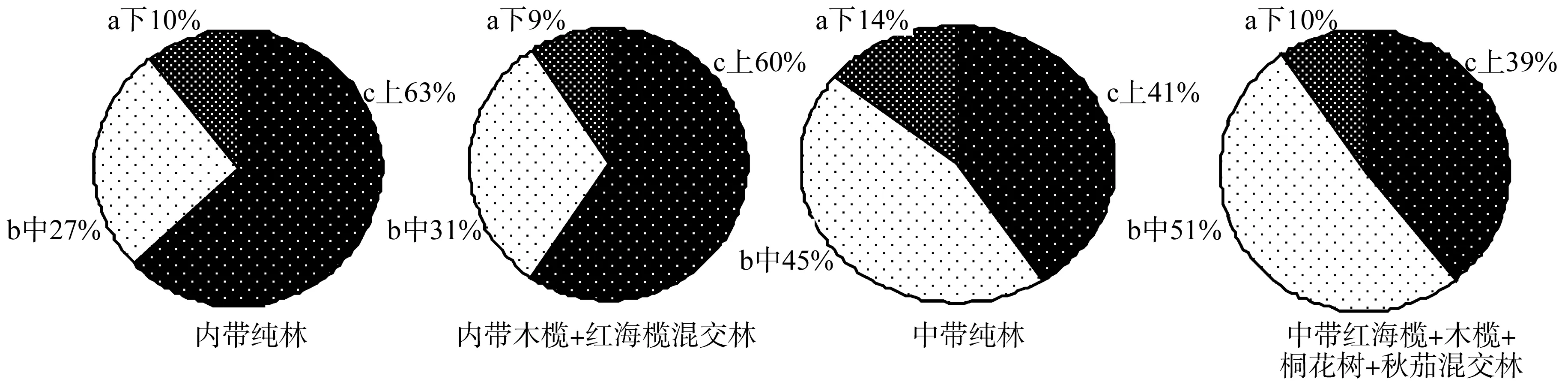
图1 不同潮带红海榄不同冠层胎生繁殖体分布Fig.1 Distribution analysis of R. stylosa community viviparous propagules of different canopys
(LSD)分析可得:内带纯林和内带混交林的东方位胎生繁殖体分布对其他方位皆呈显著性区别,内带纯林东和南方位的胎生繁殖体总和占总体的69%,西北占31%;内带混交林东和南方位的胎生繁殖体总和占总体的60%,西北占40%;中带纯林和中带混交林各个方位的胎生繁殖体分布无显著性区别,呈均匀分布。中带纯林东和南方位的胎生繁殖体总和占总体的52%,西北占48%;中带混交林东和南方位的胎生繁殖体总和占总体的55%,西北占45%。
3.1.4 背潮和面潮的胎生繁殖体空间分布比较分析
根据各参数判定标准可知(见表4),胎生繁殖体的扩散系数皆C>1;丛生指数I皆>1;负二项参数K除了内带纯林,其它皆>1; Cassie.R.M.指标CA>0;每项平均拥挤度m*>;m*/指标皆>1。胎生繁殖体在背潮和面潮呈聚集分布。从饼图比例中(图3)可知,内带纯林面潮和背潮的胎生繁殖体量差别最大,通过配对样本T检验可得:T=4.51,相伴概率为0.006,比显著性水平0.05和极显著水平0.01要小,拒绝T检验的零假设,因此,内带纯林面潮和背潮胎生繁殖体存在极显著区别;同理,内带木榄+红海榄混交林的T=2.420,相伴概率为0.046,因此存在显著区别;中带纯林的 T=1.821,相伴概率为0.111,无显著性区别;中带红海榄+木榄+桐花树+秋茄混交林的 T=0.389,相伴概率为0.707,无显著性区别。

表3 内带和中带红海榄群落不同方位胎生繁殖体聚集度分析Table 3 Aggregation degree analysis of R. stylosa community viviparous propagules in different directions in inner tide and middle tide
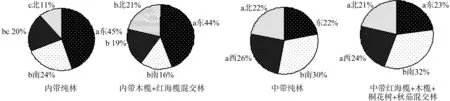
图2 内带和中带红海榄群落不同方位胎生繁殖体分布Fig.2 Distribution analysis of R. stylosa community viviparous propagules in different directions in inner tide and middle tide
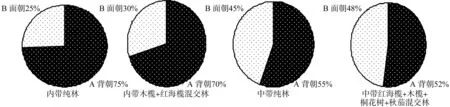
图3 背潮和面潮的胎生繁殖体分布Fig.3 Viviparous propagules distribution of back tide and face tide
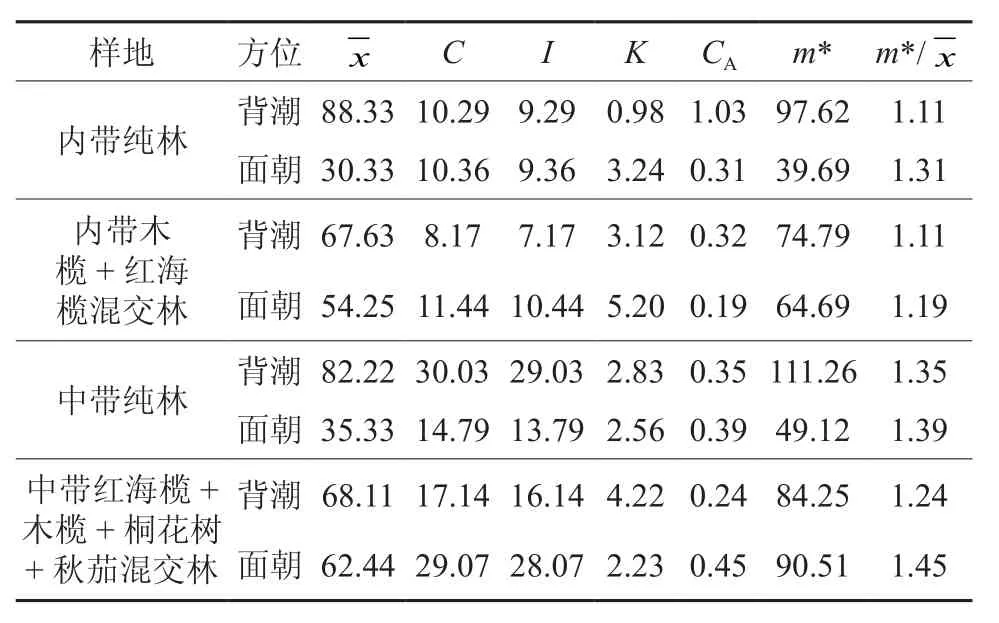
表4 背潮和面潮聚集度分析Table 4 Aggregation degree analysis of back tide and face tide
3.2.1 Lwaode m*-x回归分析
以内带和中带不同红海榄群落的胎生繁殖体拥挤度和均值进行回归分析,可得m*与的直线相关式:m*=8.9257+1.859(相关系数r=0.754)。Iwao m*-回归分析法:m*=α+β,中α=8.925 7>0,说明4个样地的红海榄胎生繁殖体在树上的分布是个体群;β=1.859>1,红海榄
3.2 回归模型分析
胎生繁殖体是呈聚集分布的。r=1或r=-1,这是相关紧密度的极端情况。一般r的绝对值愈接近于1,说明m*与x的相关愈紧密。计算的结果r值为0.754, m*和x的关系密切相关,即红海榄胎生繁殖体呈现聚集分布与密度相关。
3.2.2 Toylar冥法则分析
以内带和中带不同红海榄群落胎生繁殖体的方差s2和均值x进行对数回归,可得:lgs2=1.243+1.571lg x(相 关 系 数r=0.637),lgs2=lga+blg x式中,b=1.571>1,证明在4个样地中的红海榄胎生繁殖体呈聚集分布,而a>1,说明胎生繁殖体分布具有密度依赖性,聚集度随密度的上升而增高。
4 讨 论
不同潮带不同群落红海榄胎生繁殖体聚集强度的结果表明,一般情况下的混交林胎生繁殖体聚集度比纯林高,多种树种的混交林比两种树种的混交林聚集度高;中带纯林比外带纯林的聚集度高。由于混交林发挥了种间互补作用,合理占据了生活空间,充分利用了温度、光照、水分、土壤中的养分等生态因子,从而显著提高了林分的生产力,减轻病虫危害[13]。纯林种内间的竞争,产生种群内的自疏现象,每株红海榄之间的空间范围广,枝条生长较长,胎生繁殖体产量不多,生长较好,可能采取了K-策略,从而降低了胎生繁殖体的聚集度。
红海榄属于嗜热广布性生态类群[14-15]。在内带红海榄群落不同冠层胎生繁殖体分布格局中,内带红海榄纯林和混交林的树冠外部接受阳光总量最多,接收热量较大;同时,加上内带红海榄郁闭度高,群落平均高度有5 m以上,具有较强烈的顶端优势作用,使得中下层接受阳光的照射少,上层的枝叶比中下层更加茂盛。因此,内带红海榄胎生繁殖体主要分布在上层冠层。中带红海榄树高均低于3 m,加上该群落的其它树种普遍比红海榄低,林分空间较宽,中层的胎生繁殖体也可以得到较为充分的光照和温度,中层的枝叶多,呈圆锥状;再加上风浪对冠层上层的强烈干扰,使得胎生繁殖体主要分布在冠层中层。故中带红海榄中层的胎生繁殖体比上层和下层多。
高桥内带红海榄群落不同方位胎生繁殖体主要集中分布在东方位。从样地特征可知,内带树冠内部得到光照较少,东方位的枝叶相对能得到较多的光照。因此,胎生繁殖体平均拥挤度东侧最高。中带红海榄群落的其它树种比红海榄低,生长空间较大,红海榄可得到较多独立的生长空间,因此各个方位的胎生繁殖体数量较为平均。
高桥内带红海榄群落胎生繁殖体的背潮方位是向阳处,加上内带的面潮方向受到海风的影响较大,因此背潮面的胎生繁殖体分布较多;相对而言,中带的红海榄树高较低,接受光热比较均匀,受空间约束少,因此中带背潮和面潮面分布较为均匀。
I wao m*-x回归分析法和Toylar(1961)冥法则得出的结果表明红海榄胎生繁殖体呈现聚集分布,验证了上述胎生繁殖体不管是上中下冠层还是东南西北方位或者是背潮、面潮皆是呈聚集分布的结果。有研究表明,在一般情况下(环境均一,生存条件良好),以母株为中心的繁殖方式使低龄树呈现聚集分布;随着年龄的增加,树木对光因子和营养条件等的要求加大及自疏、它疏等作用,种群会趋于均匀分布;随着年龄继续增加,种群将趋于随机分布的格局变化规律[16]。但物种和生境的差异,格局趋势会发生相应的变化,无完全统一的格局变化规律[17-19]。胎生繁殖体的聚集分布为了适应和抵御严酷的海陆潮间带的环境,有利于种群的生长和存活,是必要的。大部分的文献并没有解释为什么红海榄散布的胎生繁殖体落在附近长成植株而形成群状呈聚集的问题。从胎生繁殖体的聚集分布,可解释红海榄的自然更新时林下幼苗和林窗幼苗为什么呈现聚集分布的其中原因之一。胎生繁殖体聚集分布的研究结果,也证明了绝大多数自然种群都服从聚集分布而不服从随机分布的规律[20]。因此,在近自然森林经营理论和森林生态系统经营理论的前提下,结合本文的研究结果,笔者提出:在严酷的环境海陆潮间带进行造林,特别是造林困难的潮带,适宜进行红海榄的合理密集种植,提高红海榄胎生繁殖体或者其他树种胎生繁殖体的环境抵御能力,从而提高存活率。同时,参照了不同年龄阶段的锐齿栎种群[10]、胡杨林[21]、不同林区栓皮栎种群[22]、红松林主要树种[23-25]等空间格局,笔者提出红海榄从胎生繁殖体至衰退老林的分布格局演替过程(见图4)。
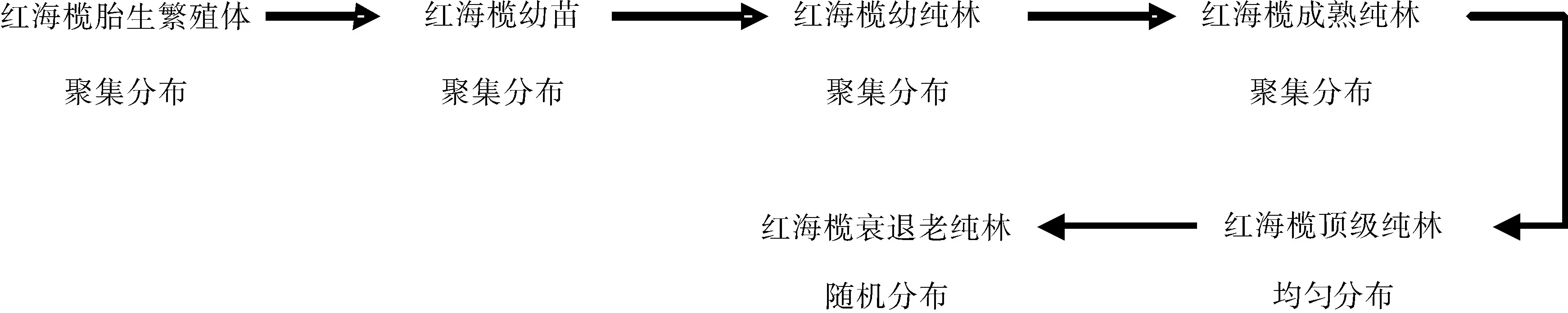
图4 红海榄分布格局演替Fig.4 Succession of distribution pattern of R. stylosa
5 结 论
红海榄生长在临海地带,受海风、海浪等外界因素的剧烈影响。红海榄除了具有胎生现象这一特征外,聚集分布对胎生繁殖体数量增长具有较大的帮助。在不同潮带不同群落中的上、中、下等不同冠层,东、南、西、北等不同方位,以及面潮和背潮两部分,通过胎生繁殖体在各个部分的分布比较和在各个部分的6种聚集度指标分析,表明高桥内带红海榄胎生繁殖体主要分布在上层冠层、东方位和背潮部分;中带红海榄胎生繁殖体主要分布在中层冠层,在各个方位以及面潮和背潮两部分的分布无明显区别;红海榄胎生繁殖体的空间分布格局皆为聚集分布。
[1] 韩维栋,高秀梅.雷州半岛红树林生态系统及其保护策略(“十五”国家重点图书出版规划)[M].广州:华南理工大学出版社,2009.
[2] 陈鹭夏,王文卿,林 鹏.潮汐淹水时间对秋茄幼苗生长的影响[J].海洋科学学报,2005,27(2):35-37.
[3] 邓英英,汤孟平,徐文兵,等.天目山近自然毛竹纯林空间分布格局研究[J].竹子研究汇刊,2011,30(1):11-22.
[4] 黄志伟,彭 敏,陈桂琛,等.青海湖几种主要湿地植物的种群分布格局及动态[J].应用与环境生物学报,2001,7(2):113-116.
[5] 李明顺,蓝崇钰,陈桂珠,等.深圳福田红树林的群落学研究Ⅱ. 多样性与种群格局[J].生态科学,1994,(1):82-86.
[6] 梁 华.澳门红树林植物组成及种群分布格局的研究[J].生态科学 ,1998,17(1):25-31.
[7] 李洪旺,王 旭,余雪标,等.海南东寨港秋茄+桐花天然红树林群落林分结构特征研究[J].热带林业,2008,36(1):30-33.
[8] 梁士楚.广西北海海岸沙生白骨壤种群分布格局研究[J].广西科学 ,2001,8 (1):57-60,69.
[9] 卢 杰,罗 建,潘 刚,等.长鞭红景天种群空间分布格局[J].辽宁林业科技 ,2009,(4):9-12.
[10] 梅世秀,袁志良,叶永忠,等.宝天曼自然保护区锐齿栎种群空间分布格局研究[J].河南农业大学学报,2011,45(2):220-225.
[11] 于传宗,慕宗杰,特日格勒,等.植物种群空间分布格局的研究方法[J].西北林学院学报,2008,(5):40-42.
[12] 张庆国,李桂亭,徐 丽.山楂叶螨种群空间格局及其应用的研究[J].应用生态学报,1993,4(4):399-403.
[13] 张庆国,徐 丽,李桂亭.苹果树冠山楂叶螨若螨空间格局的研究[J].安徽农业大学学报,1993,20(3): 234-239.
[14] 张娆挺,林 鹏. 中国海岸红树植物区系研究[J].厦门大学学报:自然科学版,1984, 23(2): 232-239.
[15] 叶 勇,曹长青.潮间带红树林分带的影响因素[J].生态学杂志 ,2008,27(4):615-618.
[16] 蔡 飞,宋永昌.武夷山木荷种群结构和动态的研究[J].植物生态学报 ,1997,21(2):138-148.
[17] 戚继忠,张 生.榛子种群空间格局及其动态的研究[J].吉林林学院学报,1996,12(2):99-103.
[18] 王得祥,陈海滨.秦岭华山松种群分布格局及动态研究[J].西北植物学报,1998,18(4):622-628.
[19] 刘仁林,朱玲琳.南方红豆杉天然群落种群空间分布规律的研究[J].环境与开发,1997,12(1):3-4.
[20] 郑元润.不同方法在沙地云杉种群分布格局分析中的适用性研究[J].林业科学,2001,37(1):6-10.
[21] 韩 路,王海珍,周正立,等.塔里木胡杨林主要空间分布格局与动态研究[J].西南林学院学报,2007,27(2):1-4.
[22] 张文辉,卢志军,李景侠,等.陕西不同林区栓皮栎种群空间分布格局及动态的比较研究[J].西北植物学报,2002,2(3):476-483.
[23] 侯向阳,韩进轩.长白山红松林主要树种空间格局的模拟分析[J].植物生态学报,1997,21(3):242-249.
[24] 周元满,聂 页,刘美欣,等.天然红海榄树冠结构的分形特征[J].中南林业科技大学学报,2012,32(8):37-41.
[25] 刘 秀,郝海坤,庞世龙,等.两种半红树植物幼苗的耐盐性研究[J].中南林业科技大学学报,2012,32(6):43-47.
Spatial distribution patterns of Rhizophora stylosa community viviparous reproduction in different tide zones
HUANG Jian-jian1a, LI Ji-ping2, LIU Su-qing1b, HAN Wei-dong1b
(1a. Cunjin College ,1b. Agriculture College,Guangdong Ocean University, Zhanjiang 524094, Guangdong, China;2.Central South University of Forestry & Technology, Changsha 410004, Hunan, China)
The investigations of the spatial distribution patterns of Rhizophora stylosa viviparous propagule in Gaoqiao of Zhanjiang,Guangdong province were conducted. The 8 aggregation degree indicators (I、C、M*、CA…) were compared and analyzed of viviparous reproductions located at upper, middle and lower canopy, in east, south, west and north azimuth, face to tide and back tide.The results indicate that R. stylosa viviparous reproduction of inner tide in Gaoqiao mainly distributed in upper crown, east position and back tide; and the viviparous reproduction of middle tide mainly distributed in middle crown, had little signif i cance difference distribution of each position and face-back tide; the distribution belonged generally to the aggregated distribution pattern. Finally, it is proposed that the R. stylosa viviparous reproduction should be planted with reasonable density in the diff i cult tide zones for afforestation,thus increasing resistance ability form environment and enhancing the survival percentage.
mangrove; Rhizophora stylosa; viviparous Propagule; distribution patterns, Zhanjiang, Guangdong province
S796
A
1673-923X(2013)03-0034-06
2012-10-16
广东省海洋渔业科技推广专项基金项目 (C20100813102523)
黄剑坚(1981-),男,广东清远人,讲师,硕士,主要研究方向为红树林生态系统和滨海生态旅游系统
刘素青(1963-),男,江西赣州人,教授,博士,主要研究方向为红树林生态恢复与生态系统经营管理
[本文编校:吴 毅]
猜你喜欢
故事作文·低年级(2020年7期)2020-07-28
儿童时代·幸福宝宝(2020年3期)2020-04-07
青少年科技博览(中学版)(2019年12期)2019-04-10
共产党员·下(2018年6期)2018-12-05
方圆(2018年23期)2018-01-07
小学阅读指南·低年级版(2017年11期)2017-12-06
小学科学(2017年11期)2017-11-23
今古传奇·故事版(2017年17期)2017-10-17
中国市场(2016年45期)2016-05-17
