
不同壳色虾夷扇贝外套膜形态学及显微结构的研究
2014-02-17 08:40赵乐丁君王俊杰郝振林常亚青
大连海洋大学学报 2014年5期
赵乐,丁君,王俊杰,郝振林,常亚青
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023)
虾夷扇贝Patinopecten yessoensis原产于俄罗斯、日本和朝鲜等海域,为冷水性双壳贝类,20世纪80年代初由日本引入中国,经多年规模化养殖,现已成为中国北方沿海地区主要经济养殖贝类。普通虾夷扇贝左壳呈褐色,右壳呈白色,且左壳较右壳稍小。大连海洋大学科研人员经过长期研究,选育出双白壳虾夷扇贝品系[1](俗称“象牙白”品系),该品系虾夷扇贝具有外形美观、出柱率高、抗逆性强、营养价值高等优点[2-3]。
外套膜是双壳贝类的重要组织器官,研究外套膜的形态结构特征,对探讨虾夷扇贝不同壳色的形成机制具有重要意义。目前,关于双壳贝类壳色形成机理的研究报道极少,仅见张安国等[4]推测,文蛤在生长发育过程中,外套膜外表皮细胞中色素细胞分泌的色素颗粒会对贝壳表面花纹的形成产生影响,而对不同壳色虾夷扇贝外套膜的组织学研究至今尚未见报道。本研究中,选取2龄普通右壳褐色虾夷扇贝、“象牙白”品系虾夷扇贝外套膜进行形态学和组织学研究,首次从形态学和细胞学水平描述不同壳色虾夷扇贝外套膜的特性,通过对外套膜上皮细胞中色素颗粒的分布进行分析,以期为研究虾夷扇贝不同壳色的形成机理提供参考依据。
1 材料与方法
1.1 材料
试验用虾夷扇贝取自大连海洋大学农业部北方海水增养殖重点实验室培育的普通褐色虾夷扇贝和“象牙白”品系虾夷扇贝2龄个体,壳高平均为4.82 cm,湿体质量平均为 12.64 g。
1.2 方法
1.2.1 形态学观察 随机选取普通虾夷扇贝和“象牙白”品系虾夷扇贝各30枚,置于盛满海水的透明水族缸中,观察外套膜充分伸展状态下的形态特征,并对其活体进行解剖,进一步观察外套膜的形态特征。
1.2.2 显微结构观察 活体解剖样品,分别取普通褐色虾夷扇贝和“象牙白”品系虾夷扇贝左、右壳外套膜具黑色条斑部位,以及无黑色条斑部位的组织块,置于卡诺氏液中固定,用常规石蜡进行包埋,切片、H.E染色后,对其进行组织学观察。
2 结果与分析
2.1 虾夷扇贝外套膜的形态学特征
虾夷扇贝外套膜是紧贴左右贝壳内面的两片薄膜,除靠近贝壳边缘部位呈游离状外,其余部位与贝壳紧密相连,与文蛤Meretrix meretrix、缢蛏Sinonovacula constricta、菲律宾蛤仔Ruditapes philippinarum等不同,虾夷扇贝无完整的水管。
虾夷扇贝外套膜主要由边缘膜和中央膜组成,中央膜是一层紧贴贝壳内面的透明薄膜,具有伸缩能力。边缘膜为一层由肌肉和感觉器官组成的较厚且呈游离状的膜,其上包含3种突起,分别为生壳突起、感觉突起和缘膜突起。生壳突起为一层白色、短小的触手,位于边缘膜最靠近贝壳一侧;感觉突起具发达的触手,位于边缘膜的中层,触手之间夹有发达的外套眼;缘膜突起 (也称帆状物)靠近内脏团,部分缘膜突起呈黑色。通过比较发现,帆状物呈黑色的部位所对应的贝壳表面颜色较其余部位所对应的贝壳表面颜色深,不同壳色虾夷扇贝左壳帆状物上的黑色条斑普遍多于右壳外套膜,90%以上普通虾夷扇贝外套膜的帆状物上均存在黑色条斑,而“象牙白”品系虾夷扇贝外套膜的帆状物上存在黑色条斑的个体较少 (图1)。
2.2 虾夷扇贝外套膜的显微结构特征
H.E染色结果显示,两种虾夷扇贝外套膜组织学结构相似,主要由内外上皮层、肌纤维、结缔组织和分泌细胞组成。虾夷扇贝外套膜显微结构概览如图2-A、B所示。
2.2.1 边缘膜 虾夷扇贝外套膜的边缘膜 (图2-C)由靠近内脏团一侧的内表皮、靠近贝壳一侧的外表皮、结缔组织和肌纤维组成,在表皮细胞间夹有分泌腺细胞。
边缘膜内表皮由高5~13 μm的单层柱状纤毛细胞构成,细胞紧密排列呈嵴状突起,核呈圆形,位于细胞基部,上皮细胞之间分布有中性空泡状分泌腺细胞。部分内表皮上皮细胞的游离端可观察到一排分泌小泡,脱离表皮细胞形成圆形或块状的分泌物 (图2-D);部分内表皮上皮细胞游离端呈明显破损状态,破损缺口周围分布有少量细胞质和分泌物 (图2-E),表明虾夷扇贝边缘膜内表皮主要以局部分泌和顶浆分泌为主。
边缘膜外表皮由高13~22 μm的单层柱状细胞构成,细胞折叠隆起呈指状突起物排列,核呈圆形,位于细胞基部,上皮细胞之间分布有大量中性空泡状分泌腺细胞。部分外表皮上皮细胞排列整齐为未分泌区域;部分外表皮上皮细胞排列松散且不整齐为分泌区域,可观察到细胞脱离上皮组织后留下的空隙 (图2-F),表明虾夷扇贝边缘膜外表皮主要以全浆分泌为主。
边缘膜内外表皮之间存在着丰富的结缔组织填充于肌纤维之间,内含较多的血腔隙、血细胞、神经细胞和神经节等。神经节呈圆形或椭圆形,位于边缘膜近缘膜突起处 (图2-G),个体较大,周围分布有神经细胞。外套膜边缘膜上的肌纤维十分发达,特别是靠近内表皮的部位含量最多,主要为排列不规则的平滑肌,越靠近中央膜,肌纤维越多,结缔组织越少。
通过比较发现,不同壳色虾夷扇贝外套膜的边缘膜在组织学结构上无明显差别,仅在个别取自不同壳色虾夷扇贝黑色条斑部位的外套膜组织块切片中,观察到内表皮上有少量色素颗粒存在 (图2-H)。
2.2.2 生壳突起 贴近贝壳的第一个呈叶片状或指状的突起为生壳突起,与感觉突起相隔的外沟处具有一团不规则的细胞团 (图2-I)。生壳突起外表皮 (近壳侧)由17~51 μm的单层柱状细胞构成,沿着生壳突起基部向顶端逐渐由矮柱状细胞变为顶端高柱状细胞。内表皮 (近感觉突起侧)由18~23 μm的单层柱状细胞排列成栅栏状。大量中性分泌腺细胞分布在生壳突起表皮细胞中,内外表皮下填充着丰富的疏松结缔组织,肌纤维呈细丝状分散在结缔组织中且数量较少 (图2-J)。
通过比较发现,不同壳色虾夷扇贝的生壳突起在组织学结构上无明显差别,也无色素颗粒存在。
2.2.3 感觉突起 虾夷扇贝的感觉突起十分发达,与缘膜突起之间以内沟相隔。感觉突起通常由1个主突和3~6个次突组成。最靠近生壳突起一侧,感觉突起上皮细胞的游离端具纤毛,为单层柱状纤毛细胞呈栅栏状排列,其余部位的感觉突起的上皮细胞均为单层柱状细胞,紧密排列呈嵴状突起,且上皮细胞间有嗜中性分泌腺细胞分布。感觉突起表皮下为结缔组织,发达的神经节呈长条状贯穿每一个次突分支中,周围分布着神经细胞体,肌纤维主要为纵肌和少量的环肌,掌管着感觉突起的伸缩(图2-K)。外套眼有规律地分布在次突之间,且靠近缘膜突起侧,眼基部以短柄与外套膜连接,主要由外角膜、色素细胞层、水晶体、血腔隙、网膜、色素层等组织构成 (图2-L)。
通过比较发现:不同壳色虾夷扇贝的感觉突起在组织学结构上无明显差别,取自不同壳色虾夷扇贝无黑色条斑部位的外套膜组织切片中,除少数个体的感觉突起表皮细胞内分布有少量色素颗粒外(图2-M),其余个体的感觉突起均无色素颗粒存在 (图2-N);而取自不同壳色虾夷扇贝具黑色条斑部位外套膜组织,感觉突起表皮细胞的顶端胞质中分布有大量棕褐色色素颗粒(图2-O、K)。
2.2.4 缘膜突起 虾夷扇贝的缘膜突起十分发达,向外翻折形成帆状物 (图2-A)。缘膜突起的外表皮由10~18 μm的单层柱状细胞构成,细胞折叠隆起呈嵴状排列,内表皮由5~12 μm的单层柱状细胞排列成栅栏状,未观察到分泌腺细胞。缘膜突起基部的结缔组织较为丰富,肌纤维含量较少,结缔组织含量由缘膜突起的基部到顶端逐渐减少,肌纤维含量逐渐增多且结构致密。
通过比较发现:不同壳色虾夷扇贝的缘膜突起在组织学结构上无明显差别,但取自具黑色条斑部位的外套膜,缘膜突起外表皮细胞的顶端胞质中分布有大量棕褐色色素颗粒 (图2-P),个别内表皮细胞中分布有色素颗粒 (图2-Q);而取自无黑色条斑部位的外套膜,缘膜突起内外表皮细胞均无色素颗粒分布 (图2-R、S)。
2.2.5 中央膜 虾夷扇贝的中央膜为紧贴贝壳的一层薄膜 (图2-T),厚度为48~135 μm,外表皮(靠近壳一侧)由9~15 μm的单层柱状纤毛细胞构成,细胞紧密排列成栅栏状,上皮细胞间分布有大量黏液细胞。内表皮 (近内脏团一侧)由19~36 μm的单层柱状纤毛细胞构成,纤毛较长,上皮细胞间分布有少量黏液细胞。中央膜内外表皮之间含有疏松的结缔组织,在结缔组织中夹杂血细胞、血腔隙、极少的肌纤维和部分成束的神经纤维。
3 讨论
在自然界中,贝壳的颜色十分丰富,比较常见的有白色、黄色、红色、紫色、黑色、橙色和蓝色等。贝壳壳色所呈现出的多态性,一是受到遗传基因的影响,例如贻贝[5]、福寿螺[6]、海湾扇贝[7]等存在壳色多态性的贝类都已证实壳色可以遗传;二是壳色的多态性与环境的影响密切相关,如温度、光照强度、盐度、生存区域 (沙层或水层)、生存深度 (水深深度)、代谢水平、背景值、耗氧率,甚至饵料等都会在一定程度上影响壳色的形成。Sokolova等[8]认为,俄罗斯白海海域盐度的突变导致该地区贝壳壳色发生多态性变化;Gonzales等[9]认为,贝壳壳色的变化受到生存环境背景值的影响;Ekendahl等[10-11]认为,滨螺不同壳色的形成与光照、生存区域和深度相关;Mitton[12]发现,壳色较深的贝类个体多分布于温度较低的水域中,而壳色较浅的个体多分布于光照充足、温度较高的水域中,深色壳色可以更多地吸收光照以用来维持自身温度,并提供运动所需能量,相反,浅色的贝壳可以反射太阳光,避免体温过热导致死亡。据此推测,“象牙白”品系所具有的高抗逆性,特别是抗高温的优点,与其贝壳颜色有着直接的关系。
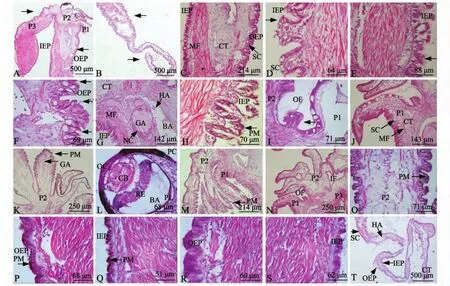
图2 虾夷扇贝外套膜显微结构Fig.2 Microstructure of mantle in yesso scallop Patinopecten yessoensis
双壳类动物的贝壳分为角质层、棱柱层和珍珠层三层。最外侧的角质层由生壳突起分泌的一层富含赖氨酸残基的胶蛋白质构成,这些硬化蛋白从生壳突起与感觉突起之间的不规则细胞团开始,紧贴感觉突起外侧走向该突起的顶部,然后包向生壳突起表面[13],角质层薄且透明,覆盖于贝壳外表面,可防止外界环境对贝壳中碳酸钙的腐蚀。对不同花色文蛤[4]外套膜的研究结果显示,有花纹文蛤个体外套膜的生壳突起外表皮细胞质中含有丰富的色素颗粒,无花纹文蛤个体外套膜的生壳突起外表皮细胞质中色素颗粒含量较少且小。但与对不同花纹文蛤的研究结论不同,本研究中并未在不同壳色虾夷扇贝的生壳突起中观察到色素颗粒,推测虾夷扇贝贝壳壳色的色素富集并未发生在生壳突起内。贝壳最中间的一层棱柱层由边缘膜外表皮分泌形成,珍珠层由中央膜外表皮分泌形成[14],而本研究中不同壳色虾夷扇贝外套膜的边缘膜外表皮和中央膜表皮细胞中均未观察到色素颗粒分布,仅个别具黑色条斑的外套膜内表皮细胞中分布有少量色素颗粒,而内表皮为靠近内脏团侧,故推测虾夷扇贝贝壳壳色的色素沉积与边缘膜和中央膜没有直接关系;不同壳色虾夷扇贝在具黑色条斑外套膜和无黑色条斑外套膜之间存在明显差异,具黑色条斑外套膜的感觉突起表皮细胞、缘膜突起外表皮细胞和个别缘膜突起内表皮细胞中均分布有大量色素颗粒,仅极少数无黑色条斑外套的感觉突起分布有少量色素颗粒,同时通过形态学观察发现,具黑色条斑外套膜所对应的贝壳表面颜色较无黑色条斑对应贝壳表面颜色深,推测虾夷扇贝感觉突起和缘膜突起上色素颗粒的分泌和富集,在一定程度上影响扇贝表面色素沉积,从而影响贝壳表面颜色的形成。
本研究结果为贝壳颜色形成机制提供一定的线索,但仅仅通过组织学对虾夷扇贝不同壳色形成机制进行研究是远远不够的。在以后的研究中,课题组将从转录组水平对虾夷扇贝壳长形成机制进行深入探讨。
[1]丁君.双白壳虾夷扇贝的培养方法[P].中国发明专利:CN101103709.2008-1-16.
[2]常亚青,陈晓霞,丁君,等.虾夷扇贝5个群体的遗传多样性[J].生态学报,2007,27(3):1145-1152.
[3]丁君,常亚青,张婧,等.虾夷扇贝“象牙白”品系与普通品系营养成分分析及评价[J].中国农业科技导报,2011,13(2):121-128.
[4]张安国,李太武,苏秀榕,等.不同花纹文蛤外套膜的显微及亚显微结构的初步研究[J].水产科学,2011,30(3):132-135.
[5]Gary J N.Genetics of shell color in Mytilus edulis L.and the association of growth rate with shell color[J].Journal of Experimental Marine Biology and Ecology,1980,47:89-94.
[6]徐建荣,沈辉,李旭光,等.不同品系福寿螺壳色性状的遗传规律分析[J].江苏农业科学,2008(4):210-212.
[7]Zheng H P,Zhang G F,Liu X.Comparison of growth and survival of larvae among different shell color stocks of bay scallop Argopecten irradians(Lamarck 1819)[J].Chinese Journal of Oceanology and Limnology,2005,23(2):183-188.
[8]Sokolova I M,Berger V J.Physiological variation related to shell color polymorphism in White Sea Littorina saxatilis[J].Journal of Experimental Marine Biology and Ecology,2000,245(1):1-23.
[9]Gonzales R,Leticia R,Silva A.Shell color polymorphism in the chitin Ischnochiton striolatus(Gray,1828)(Mollusca:Ployplacophora)and habita heterogeneity[J].Biological Journal of the Linnean Society,2005,85:543-548.
[10]Ekendahl A,Johannesson K.Shell colour variation in Littorina saxatilis Olivi(Prosobranchia:Littorinidae):a multi-factor approach[J].Biological Journal of the Linnean Society,1997,62:401-419.
[11]Ekendahl A.Colour polymorphic prey(Littorina saxatilis Olivi)and predatory effects of a crab population(Carcinus maenas L.)[J].Journal of Experimental Marine Biology end Ecology,1998,222:239-246.
[12]Mitton J B.Shell color and pattern variation in Mytihus edulis and its adaptive significance[J].Chesapeake Sci,1997,18:387-390.
[13]唐敏,石安静.贝类钙代谢研究概况[J].水产学报,2000,24(1):86-91.
[14]孙虎山,王晓安.紫彩血蛤外套膜的组织学与组织化学研究[J].烟台师范学院学报,1999,15(2):125-129.
猜你喜欢
趣味(语文)(2021年12期)2021-04-19
食品与发酵工业(2020年23期)2020-12-22
蔬菜(2020年7期)2020-12-16
家教世界·创新阅读(2019年11期)2019-12-10
家教世界(2019年31期)2019-12-05
小哥白尼(野生动物)(2019年6期)2019-10-10
农学学报(2018年2期)2018-03-03
人生与伴侣·共同关注(2018年8期)2018-02-16
农学学报(2018年11期)2018-01-16
读友·少年文学(清雅版)(2018年4期)2018-01-14
