
番茄抗TYLCV基因新标记的开发及其在多抗聚合选择中的应用
2014-02-23 10:52李菲菲杨文才
中国蔬菜 2014年10期
胡 霞 王 蓉 李菲菲 周 涛 杨文才
(中国农业大学农学与生物技术学院,北京 100193)
自20世纪90年代我国发现番茄黄化曲叶病毒病(TYLCV)以来,该病害已遍布我国番茄主产区,严重威胁我国的番茄生产(叶青静 等,2009;李小靖和叶志彪,2010;李常保 等,2012;吴篆芳 等,2013)。尽管利用杀虫剂等方法能在一定程度上控制该病毒传播者——烟粉虱的数量,但是经常施用杀虫剂会使烟粉虱产生抗药性(Horowitz et al.,2007),也会造成严重的环境污染(Palumbo et al.,2001)。采用细孔网或者紫外吸收塑料膜遮盖等物理方法(Cohen & Antignus,1994)起到了一定的防治效果,然而这些物理防治方法不能保证正常的光照,还会形成高温高湿的环境,影响番茄植株的正常生长。利用植物自身的抗性基因是防治TYLCV 最有效的方法(Morales,2001;Lapidot &Friedmann,2002)。
番茄在生长发育过程中除受到多种病毒侵染外,还会同时受到诸如斑点病、疮痂病和溃疡病等细菌性病害的危害,培育抗多种病害的品种一直是番茄育种的目标(杜永臣 等,1999)。采用常规育种方法难以在田间进行抗单一病害材料的筛选,分子标记辅助选择为解决这一难题提供了可能。Sun等(2011)根据番茄斑点病抗性基因Pto位点在抗感病材料中的序列差异建立的InDel 标记和Pei 等(2012)利用番茄疮痂病抗性基因Rx4位点在抗感材料中的序列差异建立的InDel 标记都是功能基因分子标记,能够对抗病个体进行准确的选择。而对于尚未克隆的抗性基因,则可选择与基因最紧密连锁的标记,以提高选择的准确性,如Yang 和Francis(2005)利用与番茄疮痂病抗性位点Rx3紧密连锁的标记成功地将Rx3和Pto两个基因聚合到同一材料中,育成抗番茄斑点病和疮痂病的材料。从1994年至今,人们已经开发了与5个抗TYLCV基因Ty-1~Ty-5紧密连锁的可用于抗性基因选择的分子标记(杨晓慧 等,2012),但这些标记在实际应用中都存在一些问题,如检测太复杂、稳定性和可靠性不高等。番茄基因组的出现以及抗TYLCV基因Ty-1/Ty-3的克隆,为开发稳定可靠且检测简单易行的标记提供了可能。因此,本试验的目的是开发可用于准确选择Ty-1/Ty-3和Ty-4的分子标记,并将抗病毒病基因与抗细菌病害基因聚合,培育抗多个病害的育种材料。
1 材料与方法
1.1 试验材料
本试验用于标记辅助选择的番茄(SolanumlycopersicumL.)亲本有10个,OH 9242 和里格尔87-5 分别是美国和我国广泛种植的两个加工番茄品种,但缺少抗TYLCV 的基因;IBL 2361 和ZN 17是两个育种中间材料;另有6 份材料分别作为不同病害抗性基因的供体(表1)。将这些材料配制一些杂交组合,加代得到F2群体,供标记辅助选择用。
2013年6月28日将F2种子播种于288 孔育苗盘中,育苗基质为2V草炭∶1V蛭石。从幼苗上取幼嫩叶片提取DNA 供标记辅助选择使用。2013年8月9日,将选出的与抗病亲本带型相同的F2单株定植到中国农业大学上庄实验站日光温室,收获F2:3种子。将F2:3种子用4%次氯酸钠消毒5 min,并用去离子水漂洗3~4次,干燥后保存备用。2014年1月23日播种F2:3种子。待幼苗长出2~3片真叶时,取幼嫩叶片提取DNA,供标记选择和验证使用。挑选抗性基因位点纯合的单株于2014年3月5日移栽到中国农业大学科学园温室,进行抗性鉴定。
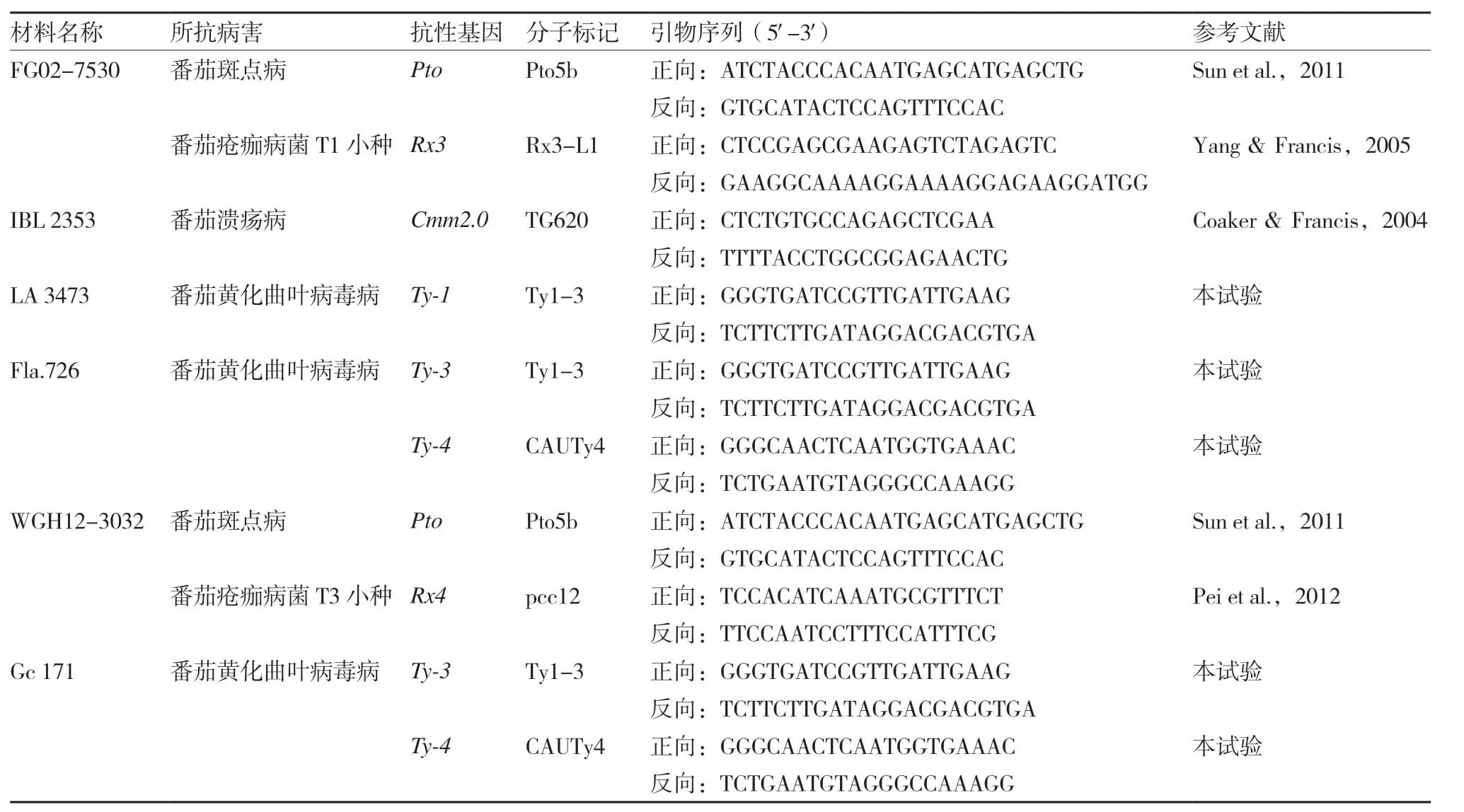
表1 所用抗病亲本及标记信息
1.2 抗TYLCV 新标记设计
为了获得稳定可靠的分子标记,本试验根据已经克隆的Ty-1和Ty-3基因(Verlann et al.,2013)位点在抗感品种中的序列差异设计引物,获得了功能基因分子标记Ty1-3。该标记是一个InDel 标记,从抗病材料中扩增到的片段大小为209 bp,从感病材料中扩增到的片段大小为197 bp。同时根据Ji 等(2009)报道的Ty-4在染色体上的位置,选择与其最紧密连锁的标记C2_At4g173000,从抗感材料中进行PCR 扩增和序列分析,发现在抗感材料中序列长度不同,据此设计了InDel 标记,命名为CAUTy4。该标记从抗病材料中扩增到的片段大小为219 bp,从感病材料中扩增到的片段大小为214 bp。原来的C2_At4g173000 是一个CAPs 标记(Ji et al.,2009),需要经过PCR、酶切、电泳等过程进行基因型检测,新设计的两个InDel 标记都只需要PCR 和凝胶电泳即可完成基因型检测。
1.3 标记辅助选择策略
采用碱裂解法(Yang & Francis,2005)提取番茄植株基因组DNA。鉴于有些杂交组合涉及到选择3个或3个以上抗性基因,为了减少工作量,本试验首先用一个功能基因标记对相应F2群体的所有植株进行筛选,只有携带纯合抗性基因位点的植株才被用于筛选第2个基因,依此类推。由于有些群体单株数不够多,没有能够获得所有基因位点都纯合的个体,因此保留了部分基因位点杂合的单株供F2:3继续选择。
番茄疮痂病T1 小种抗性位点Rx3的标记检测参考Yang 和Francis(2005)的方法,番茄疮痂病T3 小种抗性位点Rx4的功能基因标记检测参考Pei等(2012)的方法,番茄斑点病抗性基因Pto的功能基因标记检测参考Sun 等(2011)的方法,番茄溃疡病抗性位点Cmm2.0的标记检测参考Coaker 和Francis(2004)的方法,抗TYLCV基因Ty-1/Ty-3和Ty-4的检测采用如下方法。
PCR反应体系为10 μL,包括2 μL(约10 ng)DNA模板、3.4 μLTaq10 2×Master Mix(北京奥赛博科技发展有限公司生产)、0.3 μL 10 μmol·L-1正向引物、0.3 μL 10 μmol·L-1反 向引物和4 μL ddH2O。PCR 反应条件为:94℃变性5 min,38个循环的94℃变性45 s、55℃退火45 s和72℃延伸45 s,最后一步反应在72℃下保持5 min。PCR 产物经聚丙烯酰胺凝胶电泳和硝酸银染色显带后,记录每个单株的基因型。
1.4 抗TYLCV 鉴定
从F2中选出的单株定植到中国农业大学上庄实验站日光温室后自然发病,观察并记载发病情况。F2∶3家系采用人工注射接种在日光温室内进行抗性鉴定,病毒所属株系来自上海的TYLCVSH2,接种后28 d 和42 d 各调查一次。病害分级参考Friedmann 等(1998)的方法进行:0级—接种和没接种的植株一样,没有感病症状,能够正常生长;1级—顶端叶片(叶边缘)出现轻微泛黄;2级—叶片末端有些泛黄和轻微卷曲;3级—大面积叶片变黄,影响更为严重,新叶面积减少,但植株可以继续生长;4级—植物生长发育受到严重影响,叶片基本变黄,甚至变紫,杯状卷曲,停止生长。将0~2级划分为抗病,3~4级划分为感病(Ji et al.,2009)。
由于少数入选的F2:3家系群体较小,因此将来自于同一个F2群体的不同F2:3家系算作一个群体,按100×∑(各级病株数×各级代表值)/(调查总株数×最高级代表值)进行病情指数计算。
2 结果与分析
2.1 抗TYLCV基因新标记的可靠性分析
为了检验新设计的用于选择抗TYLCV基因标记的可靠性,本试验用Ty1-3 和CAUTy4 这两个标记分析了10 份携带Ty-1、Ty-3、Ty-4中1个或多个基因的番茄材料和1 份新的抗病材料LA 1589 的基因型,以感病品种OH 88119 作为对照。结果显示,两对引物扩增得到的基因型与材料所携带的已知抗性基因完全一致(表2、图1),表明用这两个标记来检测各自的基因型是准确可靠的。同时也发现,新的抗性材料LA 1589 不含有Ty-1、Ty-3和Ty-4基因中的任何一个。
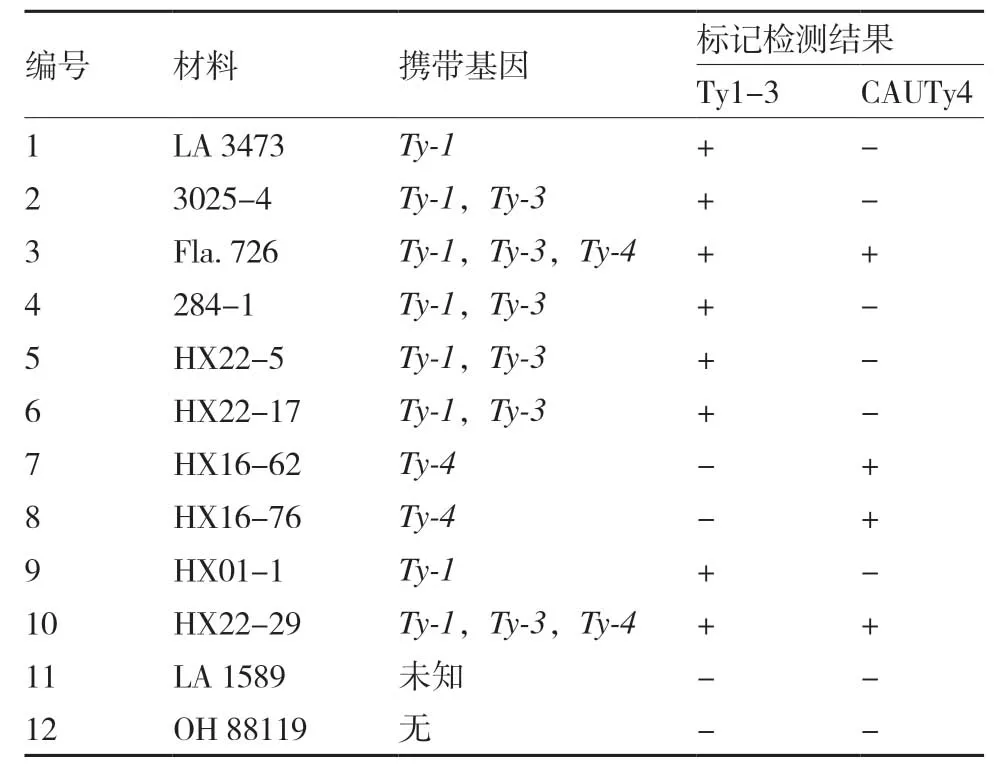
表2 12 份番茄材料经标记Ty1-3 和CAUTy4 检测的基因型及其携带的抗性基因
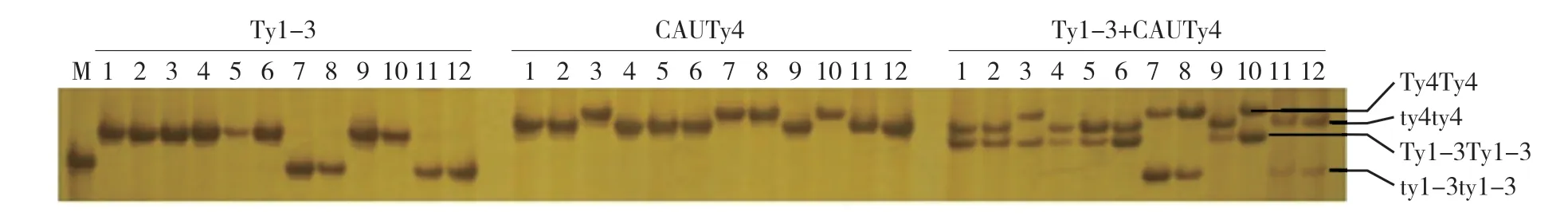
图1 普通PCR和多重PCR的电泳对比
2.2 标记Ty1-3和CAUTy4的多重PCR体系建立
为了提高检测效率,本试验尝试构建Ty1-3 和CAUTy4 这两个标记的多重PCR 体系,在PCR 体系中同时加入两个标记的引物,对上述12 份材料的基因组DNA 进行PCR 扩增。结果显示(图1),两对引物单独使用时都能从12 份材料中扩增出清晰的条带,同时使用两对引物扩增时,两对引物的PCR 产物条带依然清晰,表明这两个标记可以用多重PCR 进行检测。
2.3 不同抗性基因的聚合选择和抗TYLCV 的鉴定结果
采用功能基因标记Ty1-3 对8个杂交组合(表3)的F1进行PCR 扩增,所有F1植株都带有各自亲本的条带,为杂合型,表明所有单株都是真杂种,为获得用于标记辅助选择的F2群体提供了保障。
根据不同杂交组合所选择基因的需要,确定了每个F2群体的播种种子数,但由于出苗等原因,有些群体没有足够的苗供筛选所有抗性基因位点都纯合的个体,因此在实际筛选时保留少量杂合的基因型,8个组合共获得了57个单株(表3)。

表3 各组合标记选择获得植株数和抗性鉴定结果
将这57个单株于2013年8月9日定植到日光温室里,植株在生长过程中受到了烟粉虱为害,植株自然发生TYLCV 病害。调查显示,所有单株的TYLCV 病级都在0~2 之间,表现出了较好的抗性,并能正常开花坐果,特别是从FG 02-7530×Gc 171组合中选出的4个单株,植株生长健壮,没有任何发病症状(图2-A)。而同时种植在该日光温室里的不带抗性基因的材料则表现出TYLCV 的典型症状,叶片向上卷曲,叶片黄化,植株矮小,生长极其缓慢,不能开花坐果(图2-B)。

图2 FG 02-7530×Gc 171 组合入选植株和不带抗TYLCV基因的植株自然发病情况
对从F2群体中选出的部分一个位点纯合而另外位点杂合的单株F2:3家系进行了标记辅助选择,连同从F2中选出的纯合单株,共得到21个F2:3家系582个单株,包括4个携带5个抗性基因的家系、9个携带3个抗性基因的家系、2个携带2个抗性基因的家系和6个携带1个抗性基因的家系(表3)。
由于接种TYLCV后的两次调查结果基本一致,因此这里只用了接种42 d后的病级及病情指数。接种42 d后,所有单株的病级都在0~2 之间,其中0级的占88.0%,1级的占8.2%,2级的占3.8%。表明采用标记选出的所有单株对所用的病毒株系都具有抗性。
从里格尔87-5×Gc 171 组合中选出两种类型的家系,一种只带Ty-1/Ty-3基因,另一种携带Ty-1/Ty-3和Ty-4基因。前者共有60个单株,其中22个为0级,18个为1级,20个为2级,病情指数为24.2。后者共41个单株,包括16个0级,24个1级和1个2级,病情指数为15.9,而且这两个家系的病情指数都明显高于其他家系(表3)。
3 结论与讨论
用于聚合育种的分子标记必须具有检测的可靠性和操作的简单性,这样才能快速准确地选到目标基因。根据已经克隆的基因在功能区域的序列差异设计标记,是获得较为可靠的功能基因分子标记的途径之一。已有的抗TYLCV基因定位研究表明,抗性基因Ty-1和Ty-3都位于6号染色体上(Zamir et al.,1994;Ji et al.,2007),进一步的研究指出,Ty-1和Ty-3为等位基因(Verlaan et al.,2013)。本试验依据已经克隆的TYLCV基因Ty-1和Ty-3与感病材料中对应基因的序列差异设计引物,建立了功能基因分子标记Ty1-3。用该标记对Zamir 等 (1994)报道的携带Ty-1基因的材料LA 3473 进行基因型分析,结果显示其条带与携带Ty-3材料的条带一致,也验证了Ty-1和Ty-3是等位基因。由这个标记选择的单株都具有很好的抗病性,表明该标记的确为功能基因分子标记,可用于对这两个基因的选择。而Ji 等(2009)报道的与Ty-4基因紧密连锁的标记是CAPs 标记,需要经过PCR、酶切等相对复杂的过程,本试验中根据该标记在抗感材料中的序列差异设计成InDel 标记,只需要PCR 即可,简化了检测过程。用两个新设计的标记筛选相应的基因,所得到的植株都表现出很好的抗性,表明这两个新标记是可靠的。
利用不同的材料将不同的抗性基因聚合到同一个材料中,能够增加番茄材料抗病毒生理小种的范围,大大提高植株对病害的抗性,使其抗性更加稳定 持 久(Banerjee & Kallo,1987;Vidavski et al.,2008)。Mejía 等(2005)利用不同的番茄材料在危地马拉进行抗病性评估,含有纯合Ty-2基因型的材料在当地表现为感病,含有Ty-2和Ty-3a杂合基因型的材料表现抗病。另外,含有杂合基因型Ty-2和Ty-3a的材料比只含Ty-3纯合基因型的材料表现出更高的抗性水平。在本试验中,无论是只含有Ty-1,或者Ty-1和Ty-3,还是同时含有Ty-1、Ty-3和Ty-4的材料,对所用的病毒菌株都具有较好的抗性。但在不同的遗传背景下抗性略有差异,当以FG 02-7530、IBL 2353、IBL 2361、OH9242、ZN 17、WGH12-3032 等为受体材料时,所得到的植株都表现出很强的抗性,而以里格尔87-5 为受体时,后代中无论是携带Ty-1/Ty-3基因还是同时携带Ty-1/Ty-3和Ty-4基因的植株,其抗性都比其他材料的略差,不过同时携带Ty-1/Ty-3和Ty-4基因的植株抗性要好于只携带Ty-1/Ty-3基因的植株。这些结果表明,遗传背景对抗性基因的表达可能有一定的影响。
群体大小是影响多基因聚合育种效率的因素之一,聚合效率随着群体中个体数的增加而加大,如果基础群体太小,就有可能得不到所需要的纯合基因型个体。本试验根据每个F2群体所聚合的基因数目确定的播种量播种,但由于种子发芽率偏低(种子收获于2012年12月),因此8个F2群体中有6个没有达到预期的供选植株数,导致从群体中选择所有基因位点都纯合的单株数偏少,在5个聚合2~5个基因的F2群体中只有2.7%的单株在所有基因位点都表现为纯合型。为了获得更多在所有位点都纯合的单株,在实际选择时保留了一些杂合基因型个体供F2:3进一步选择,这样做虽然最终获得了所有位点都纯合的基因型个体,但增加了世代数和选择成本,降低了聚合效率。因此,在进行聚合育种时,可先检测种子的发芽率,然后结合聚合的基因数确定播种量,以便充分发挥分子标记聚合育种的优势。
本试验所用的番茄材料涉及到斑点病、疮痂病、溃疡病和TYLCV,但仅对所选到的植株进行了抗TYLCV 的鉴定。原因在于选择番茄斑点病抗性基因Pto和疮痂病抗性基因Rx4用的是功能基因分子标记,这两个基因的功能分子标记已经被证实可直接用于抗性基因的选择(Yang & Francis,2005;Pei et al.,2012)。疮 痂 病 抗 性位点Rx3和溃疡病抗性位点Cmm2.0是两个尚未被克隆的QTL,但是已有的研究表明,利用与Rx3位点紧密连锁的标记可准确选择到携带该位点的抗性单株(Yang & Francis,2005;张晓敏 等,2009),而利用与Cmm2.0紧密连锁的标记也可准确获得抗溃疡病的单株(Coaker & Francis,2004)。鉴于本试验所用的抗性材料和分子标记与文献中(Coaker & Francis,2004;Yang & Francis,2005;张晓敏 等,2009;Pei et al.,2012)的相同,因此推测本试验所选的携带抗性基因的材料即为抗病材料。
综上所述,本试验开发了抗TYLCV基因Ty-1/Ty-3和Ty-4的新标记,并建立了多重PCR 体系。通过杂交结合标记辅助选择,将抗TYLCV、斑点病、疮痂病、溃疡病等的基因分别聚合,获得了7份兼抗斑点病、TYLCV 和疮痂病的材料,6 份携带多个TYLCV基因(Ty-1/Ty-3和Ty-4)的材料,以及1 份兼抗TYLCV 和溃疡病的材料,为番茄抗多个病害分子标记聚合育种提供了材料。
杜永臣,严准,王孝宣,李树德,朱德蔚.1999.番茄育种研究主要进展—文献综述.园艺学报,26(3):161-169.
李常保,崔彦玲,张丽英,李传友.2012.番茄黄化曲叶病毒的快速分子检测.遗传,34(3):366-370.
李小靖,叶志彪.2010.我国番茄黄化曲叶病发生规律和研究进 展.长江蔬菜,(2):1-5.
叶青静,杨悦俭,王荣青,李志邈,阮美颖,周国治,姚祝平.2009.番茄抗黄化曲叶病育种研究进展.中国农业科学,42 (4):1230-1242.
吴篆芳,曹坳程,郑建秋,郭美霞,王秋霞,李园,颜冬冬,毛连纲,马涛涛.2013.番茄黄化曲叶病毒病研究进展.作物杂志,(1):18-23.
杨晓慧,国艳梅,王孝宣,高建昌,杜永臣.2012.番茄抗黄化曲叶病基因与基因工程研究最新进展.园艺学报,39(11):2283-2290.
张晓敏,Francis D M,杨文才.2009.我国部分番茄主栽品种抗疮痂病评价和标记辅助选择.华北农学报,24(4):183-187.
Banerjee M K,Kallo O.1987.Inheritance oftomato leaf curl virusresistance inLycopersicon hirsutumf.glabratum.Euphytica,36:581-584.
Coaker G L,Francis D M.2004.Mapping,genetic effects,and epistatic interaction of two bacterial canker resistance QTLs fromLycopersicon hirsutum.Theor Appl Genet,108:1047-1055.
Cohen S,Antignus Y.1994.Tomato yellow leaf curl virus(TYLCV),a whitefly-borne geminivirus of tomatoes.Adv Dis Vector Res,10:259-288.
Friedmann M,Lapidot M,Cohen S,Pilowsky M.1998.A novel source of resistance totomato yellow leaf curl virusexhibiting a symptomless reaction to viral infection.J Amer Soc Hort Sci,123 (6):1004-1007.
Horowitz R,Denholm I,Morin S.2007.Resistance to insecticides in the TYLCV vector,Bemisia tabaci// Czosnek H.Tomato yellow leaf curl virusdisease.Netherlands:Springer:305-325.
Ji Y,Schuster D J,Scott J W.2007.Ty-3,a begomovirus resistance locus near thetomato yellow leaf curl virusresistance locusTy-1on chromosome 6 of tomato.Mol Breed,20:271-284.
Ji Y,Scott J W,Schuster D J.2009.Molecular mapping ofTy-4,a newtomato yellow leaf curlvirusresistance locus on chromosome 3 of tomato.J Amer Soc Hort Sci,134:281-288.
Lapidot M,Friedmann M.2002.Breeding for resistance to whitefly- transmitted geminiviruses.Ann Appl Biol,140:109-127.
Mejía L,Teni R E,Vidavski F,Czosnek H,Lapidot M,Nakhla M K,Maxwell D P.2005.Evaluation of tomato germplasm and selection of breeding lines for resistance to begomoviruses in Guatemala.Acta Hortic,695:251-255.
Morales F J.2001.Conventional breeding for resistance toBemisia tabacitransmitted geminiviruses.Crop Protect,20:825-834.
Palumbo J C,Horowitz A R,Prabhaker N.2001.Insecticidal control and resistance management forBemisia tabaci.Crop Protect,20:739-766.
Pei C C,Wang H,Zhang J Y,Wang Y Y,Francis D M,Yang W C.2012.Fine mapping and analysis of a candidate gene in tomato accession PI128216 conferring hypersensitive resistance to bacterial spot race T3.Theor Appl Genet,124:533-542.
Sun W Y,Zhao W Y,Wang Y Y,Pei C C,Yang W C.2011.Natural variation ofPtoandFengenes and marker-assisted selection for resistance to bacterial speck in tomato.Agric Sci China,10 (6):827-837.
Verlaan M G,Hutton S F,Ibrahem R M,Kormelink R,Visser R G F,Scott J W,Edwards J D,Bai Y L.2013.Thetomato yellow leaf curl virusresistance genesTy-1andTy-3are allelic and code for DFDGD class RNA dependent RNA polymerases.PLoS Genet,9(3):e1003399.
Vidavski F,Czosnek H,Gazit S,Levy D,Lapidot M.2008.Pyramiding of genes conferring resistance totomato yellow leaf curl virusfrom different wild tomato species.Plant Breed,127(6):625-631.
Yang W C,Francis D M.2005.Marker-assisted selection for combining resistance to bacterial spot and bacterial speck in tomato.J Amer Soc Hort Sci,130:716-721.
Zamir D,Ekstein M I,Zakay Y,Navot N,Zeidan M,Sarfatti M,Eshed Y,Harel E,Pleban T,van-Oss H,Kedar N,Rabinowitch H D,Czosnek H.1994.Mapping and introgression of atomato yellow leaf curlvirustolerance gene,Ty-1.Theor Appl Genet,88:141-146.
猜你喜欢
广西林业科学(2022年6期)2023-01-16
南方农业(2022年17期)2022-10-21
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
河北科技师范学院学报(2022年2期)2022-08-26
农业科技通讯(2022年7期)2022-07-19
安徽农学通报(2022年6期)2022-04-07
空间科学学报(2021年1期)2021-05-22
广西农学报(2019年1期)2019-07-18
中国果菜(2016年9期)2016-03-01
