
红平菇漆酶基因异源表达及对染料脱色的研究
2014-03-27 02:05郑苗苗邵淑丽张东向张令昂焦战战
纺织学报 2014年12期
郑苗苗, 邵淑丽, 张东向, 张令昂, 焦战战
(齐齐哈尔大学 生命科学与农林学院, 黑龙江 齐齐哈尔 161006)
漆酶(laccase)是一种含铜的多酚氧化酶,属于蓝色多铜氧化酶家族[1]。漆酶通过催化O2将4个电子还原生成水,底物作用范围广泛,在很多方面有较大的应用价值[2-3]。最早由日本吉田在漆树的分泌物中发现漆酶,随后又在一些真菌中发现漆酶。近些年来,漆酶一直是医学、生物学、化学和环境科学等领域中十分活跃的研究热点[3]。迄今为止,漆酶广泛存在于植物、真菌、细菌、昆虫中[2]。其中真菌中白腐菌是最重要的漆酶生产菌,但是该菌漆酶产量低、热稳定性差、生长周期较长,而且漆酶的分泌还需要酚类化合物作为诱导,不利于大规模工厂化生产[4-5]。近年来,漆酶相关研究在国际上越来越受到重视,漆酶不仅具有降解木质素特性,还能降解多种复杂化合物,所以漆酶在印染工业、纺织工业、造纸工业及其他领域有相当大的研究价值及应用潜力,因此,探索廉价、安全的漆酶生产方法策略吸引了国内外学者的兴趣。国内外学者已在对漆酶异源表达的研究方面有了许多报道。多个漆酶基因已实现了异源表达,宿主有酿酒酵母(Saccharomyces cerevisiae)[6]、甲醇毕赤酵母(Pichia methanolica)[7]和构巢曲霉(Aspergillusnidulans)[8]等。
染料的降解已成为污水处理的首要问题[9]。各种染料在化学结构上差异较大,而且污染后的水对生物具有毒性。由于复杂的化学结构,许多染料很难降解。以往化学和物理方法处理废水效果较差,而且成本高。白腐真菌漆酶在处理废水中染料具有相当大的优势。目前为止,白腐真菌降解染料是最有效的途径[10],因此,通过异源表达获得的重组漆酶对染料脱色效率的研究,以及在污水处理中的应用有着非常重要的研究价值。
红平菇是一种味道鲜,具蟹味,既可作美味佳肴,又可作盆景观赏的食用菌,也是一种白腐真菌。到目前为止,已经对红平菇的栽培技术、多糖的结构和纯化、抗肿瘤方面和保健功能以及相关基因等有了较深入的研究[11]。本文中所用的红平菇菌株购自福建三明真菌研究所,菌株生长条件适合于北方地区栽培。由于红平菇既具有食用价值,又具有分解各种合成染料的潜在能力,因此,研究其相关基因和异源表达具有十分重要意义。本文研究了红平菇漆酶Pd-lac1基因在酵母中完成异源表达、酵母重组菌株分泌漆酶变化曲线和对不同复杂合成染料的脱色率,报道了从红平菇中克隆的漆酶基因在毕赤酵母中实现了异源表达和分泌表达的产酶变化曲线,及重组漆酶对染料脱色的研究,为今后漆酶在治理环境方面的应用奠定了一定的基础。
1 实验部分
1.1 材 料
菌株:红平菇菌株Pleurotusdjamor,由齐齐哈尔大学生物技术学科微生物实验室提供。PichiapastorisSMD1168H(菌株号)购自Invitrogen公司;E.coliTop10感受态细胞,E.coliJM109感受态细胞,购自大连宝生物公司。
质粒:表达载体pPICZB购自Invitrogen公司;克隆载体pMD18-T simple veetor购自大连宝生物公司。
YPD/Zeocin平板培养基:15 g/L蛋白胨;15 g/L琼脂粉;15 g/L酵母粉;15 g/L葡萄糖; 100 μg/mL Zeocin。
Low Salt LB液体培养基:10 g/L酵母粉;5 g/L胰蛋白胨;10 g/L NaCl。
Low Salt LB/Zeocin平板培养基:5 g/L酵母粉;15 g/L琼脂粉;10 g/L胰蛋白胨;5 g/L NaCl; 25 μg/mL Zeocin。
BMMY液体培养基:20 g/L蛋白胨;4×10-5%生物素;10 g/L酵母粉;50 mmol/L磷酸钾缓冲液(pH值为6.5); 1.34% YNB;1%甲醇。
BMGY液体培养基:20 g/L蛋白胨;4×10-5%生物素;10 g/L酵母粉;50 mmol/L磷酸钾缓冲液(pH值为6.6); 1.34% YNB; 1%甘油。
试剂:Zeocin、XbaI、BglⅡ、EcoRI限制性内切酶(NEB)、T4 DNA连接酶(NEB)购自Invitrogen公司;DNAquick_真菌基因组DNA提取系统(TIANDZ)、E.Z.N.A真菌RNA提取试剂(TIANGEN)、两步法RT-PCR试剂盒、TaKaRa Genome Walking Kit试剂盒、质粒DNA小提试剂盒(TIANGEN)、高保真Taq酶、DL 15,000 DNA Marker(TaKaRa)、Tryptone、Yeast Extract(Oxoid)、生物素D-biotin(sigma);山梨醇D-sorbitol(Difco);溶壁酶Lyticase(Tiangen);YNB(Difcol)、DMP(2,6-Dimethoxyphenol)、ABTS[2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic)],购自大连宝生物公司。
1.2 红平菇总RNA的提取
采用文献[12]中优化后的漆酶培养基培养红平菇。用OMEGA试剂盒提取红平菇总RNA,产物用核酸检测仪测定纯度,通过琼脂糖凝胶电泳检验完整性。
1.3 红平菇漆酶Pd-lac1完整CDS序列克隆
1.3.1试验酶切引物的设计
扩增漆酶CDS序列引物如表1所示。

表1 扩增漆酶CDS序列引物Tab.1 Primers for amplifying laccase CDS
1.3.2漆酶完整CDS序列的PCR扩增
使用二步法RT-PCR试剂盒进行RT-PCR扩增,以总RNA为模板进行RT-PCR的反转录cDNA第一链扩增,用酶切引物进行PCR扩增,获得漆酶CDS序列。反应体系为50 μL,含5×Prime STARTM Buffer(Mg2+plus)9 μL,dNTP Mixture(10 mM)5 μL,Prime STARTM HS DNA Polymerase(2.5 U/μL)0.5 μL,引物Pdlac1-RT1和Pdlac1-RT2各1 μL,反转录cDNA 5 μL。反应条件: 95 ℃预变性5 min,95 ℃ 40 s,55 ℃ 45 s,72 ℃ 5 min,35个循环,72 ℃ 8 min。PCR产物经凝胶电泳检测,检测结果用TaKaRa Genome Walking Kit试剂盒纯化,与pMD18-T simple veetor连接,转化到E.coliJM109中,在含有IPTG、X-Gal、Amp的LB琼脂培养基上进行培养,形成单菌落。挑取白色单菌落放入含有Amp抗性的液体LB培养基中,25 ℃、180 r/min振荡培养约8 h,提质粒进行PCR鉴定。
1.4 红平菇漆酶Pd-lac1重组质粒的构建
首先用EcoRI、XbaI限制性内切酶将目的基因Pd-lac1与载体pMD18-T Semple Vector进行双酶切,之后用T4 DNA连接酶将载体与Pd-lac1基因进行连接,获得重组质粒pPICZB-Pd-lac1,连接液转化E.coliTop10,涂布在Low Salt LB/Zeocin平板上。随机选择转化子提取质粒,用限制性内切酶对质粒进行酶切分析,转化后送到生物公司进行测序,验证目的片断是否Pd-lac1基因。
1.5 转化毕赤酵母及鉴定转化子
pPICZB-Pd-lac1质粒用BglⅡ进行酶切线性化。产物进行凝胶回收,保存备用。Pichiapastoris在YPD平板上分离单菌落,28 ℃培养3 d,挑取酵母单菌落到5 mL YPD培养基中,取100~500 μL转入100 mL YPD液体培养基中,培养24 h时OD600值为1.1~1.4。获得细胞并重悬于70 mL冰冷无菌水中, 4 ℃、4 000 r/min离心3 min,重复3次。再将菌体重悬于5 mL冰冷的1 mol/L山梨醇中,4 ℃、4 000 r/min离心3 min。最后用200 μL 1 mol/L冰冷山梨醇悬浮细胞使其终体积为400 μL。获得感受态细胞存放在冰块上,当天使用。
将质粒pPICZB-Pd-lac1、0.2 cm电转杯、酵母感受态细胞放在冰块上预冷。设置转化参数:电压为1 600 V,电击时间为6 ms。取5~10 μg线性化的重组质粒与60 μL感受态细胞混合,转至0 ℃预冷的0.2 cm电转杯中。在2 s之内按PULSE键2次,进行电击转化,之后转移到离心管内,28 ℃培养3 h。加入1 mL YPD培养基,28 ℃、180 r/min培养3 h。取100 μL培养物涂布在含Zeocin的YPDS平板上,形成阳性转化子。
以破壁后的酵母裂解液为模板,用特异性酶切引物进行PCR扩增。反应条件同1.3.2。将PCR产物经琼脂糖凝胶电泳检测,验证Pd-lac1基因是否插入酵母染色体中。
1.6 含漆酶酵母重组子的诱导表达
将正确阳性转化子菌落和含有空pPICZB载体的对照菌落分别接种至25 mL BMGY液体培养基。28 ℃振荡培养至吸光值OD600为3~4之间。离心收集菌体,转入BMMY培养基,28 ℃、180 r/min培养96 h,每24 h加入无水甲醇至终用量为0.5%。每12 h取菌液进行漆酶活性测定。
1.7 漆酶活性测定
1个酶活力单位定义为每分钟氧化1 μmol底物所需的酶量。420 nm下检测漆酶与底物在室温反应3 min的吸光度值。测定反应体系为:空白对照管中加1.95 mL柠檬酸-磷酸盐缓冲液(pH值为3.4)和50 μL稀释后的酶液;检测管中加0.95 mL柠檬酸-磷酸盐缓冲液(pH值为3.4),50 μL稀释后的酶液,1 mL浓度为1 mmol/L的ABTS。测定结果重复3次。
1.8 粗漆酶的制备
将重组菌毕赤酵母pPICZB-Pd-lac1接种到BMGY培养基,27 ℃、240 r/min 振荡培养15~25 h至吸光值OD600为2~6之间;然后将粗酶液经4 000 r/min,离心12 min,收集沉淀重新悬浮BMMY诱导培养基中,同时添加0.25 mmol/L的CuSO4溶液,振荡培养,间隔一定时间加入5%甲醇作为诱导物,培养到第3天,离心收集粗酶液,在-80 ℃冰箱中冷冻保存。
1.9 漆酶的纯化
对收集的上清液进行超滤浓缩,通过DEAE-Sepharose CL-6B离子交换柱层析,再用聚乙二醇浓缩。收集漆酶蛋白,上样到凝胶过滤层析柱SephadexG-75上,用pH值为6.5的20 mmol/L醋酸缓冲液进行洗脱,流速设为0.5 mL/min,自动部分收集器收集100管,每管3 mL。对纯化的漆酶蛋白用SDS-PAGE进行电泳。
1.10 粗酶液对合成染料的脱色
染料:SN4R,杭州方顺化工有限公司;甲基橙,沈阳市新光化工厂公司;孔雀绿,上海抚生实业有限公司;中性红,天津市大茂化学试剂厂。以上染料分别置于锥形瓶内,放置在干燥、阴凉处妥善保管。
利用紫外-可见分光光度计在300~800 nm区间扫描,得到在此区间4种染料的最大吸收峰值:孔雀绿为558 nm,吸光度=3.511;中性红为527 nm,吸光度=3.514;甲基橙为486 nm,吸光度=1.876;SN4R为432 nm,吸光度=0.659。脱色体系中加入50 mmol/L(pH值=4.6)柠檬酸-磷酸盐缓冲溶液1 mL,分别从锥形瓶中吸取1 mL(40 mg/L)待降解4种染料,最后加入重组漆酶酶液1 mL 启动反应,在25 ℃下静置反应,每隔一定时间检测4种染料吸光度值为A1;以同样方法,反应中加入等量含pPICZB载体的酵母酶液作对照,测其吸光度值为A0。脱色率计算公式为:

2 结果与讨论
2.1 漆酶完整CDS序列的PCR扩增
用酶切引物进行PCR扩增,得到Pd-lac1 CDS全长序列,条带大小为1.6 kb(见图1)。图中:泳道1—Marker 2000; 泳道2—对照; 泳道3—样本。将PCR产物进行凝胶回收,回收大小与PCR结果一致,约1.6 kb,将回收的CDS与载体连接、进行转化,筛选出阳性转化子,挑取阳性菌落至含抗性液体LB培养基中进行培养,以菌液为模板进行PCR验证,结果为单一目的条带,之后进行质粒提取,同时将菌液送到生物公司测序,进一步验证是否为漆酶CDS基因。将该序列测序结果输入到GenBank中Blastx比对后,与其他真菌漆酶基因具有较高的同源性。其中与偏肿革裥菌Lenzitesgibbosa的基因序列同源性评价最高,相似性达86%,与褶孔栓菌Trametesgibbosa、毡毛栓孔菌Trametesvelutina、云芝Trametesversicolor的漆酶基因序列相似性达到77%~79%。GenBank上的登录号为KF700372。
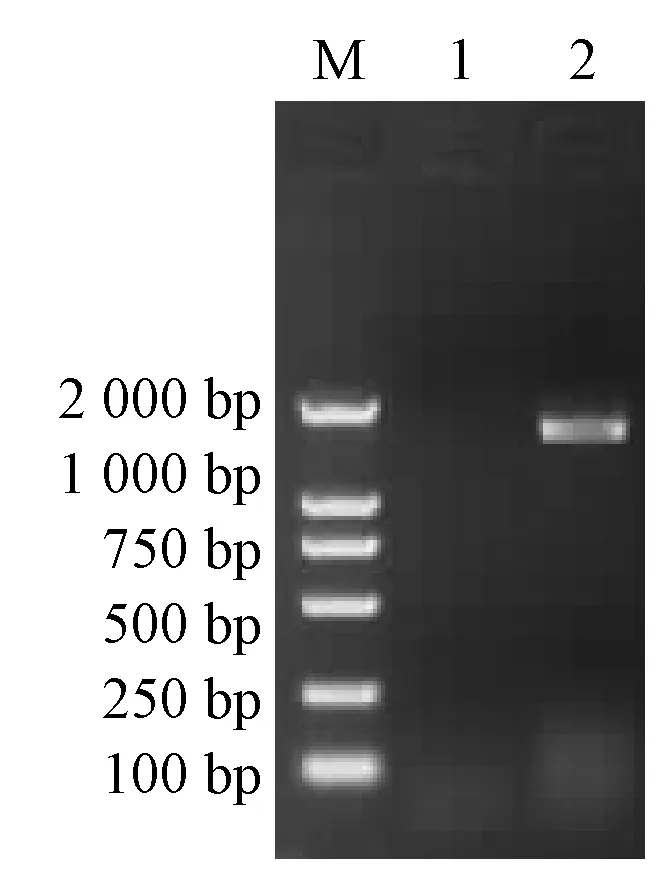
图1 Pd-lac1的RT-PCR扩增完整CDS序列结果Fig.1 Pd-lac1 by RT-PCR amplification of complete CDS sequence results
2.2 pPICZB-Pd-lac1表达载体构建及转化
将带有自身信号肽漆酶CDS克隆到毕赤酵母表达载体pPICZB的AOX1启动子下游(见图2),表达载体pPICZB-Pd-lac1用EcoRI和XbaI进行双酶切,得到与漆酶CDS序列大小一致的片段,表明漆酶CDS序列已经插入到表达载体pPICZB中,同时验证了表达载体pPICZB双酶切产物(见图3)。其中泳道1为Marker 15000,泳道2~4为样本。从Low Salt LB平板上挑取阳性转化子,进行菌液培养后,提取重组质粒pPICZB-Pd-lac1,进行PCR验证。将验证正确的重组酵母菌株接种到含有Zeocin抗性的YPD培养基上,筛选转化子。在YPD/Zeocin培养基上随机挑取3个重组酵母单菌落,破壁后分别用带酶切位点的引物进行PCR验证(见图4)。图中:泳道1为Marker 2000;泳道2为对照;泳道3~5为样本。

图2 重组表达载体pPICZB-Pd-lac1结构示意图Fig.2 Construction of P. djamor expression pPICZB-Pd-lac1

注:1—pPICZB载体质粒双酶切产物;2—重组质粒双 酶切验证结果;3—完整CDS序列扩增结果。图3 酶切验证结果Fig.3 Digested validation results

图4 重组酵母菌落PCR鉴定结果Fig.4 Direct PCR screening of recombinant Pichia colonies
2.3 重组漆酶的诱导表达
分别挑取8株含有重组表达载体pPICZB-Pd-lac1和1株含空载体pPICZB的重组酵母进行培养,从诱导开始,每天检测重组菌漆酶酶活。从前者发酵液中检测到漆酶酶活,空载体对照中没有检测到。酶活性较高的重组菌株发酵液,在72 h时最高漆酶活性达到54 U/L,之后漆酶活性逐渐下降(见图5)。
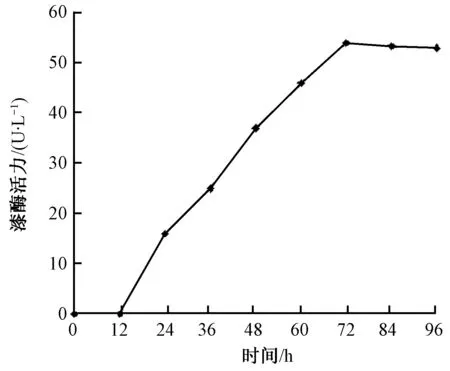
图5 重组毕赤酵母甲醇诱导表达的漆酶活性Fig.5 Enzyme activity of laecase of recombinant Pichia Pastoris under indueing condition
重组菌株在甲醇诱导下,第3天漆酶活性达到最高(54 U/L),是原菌株漆酶活性的1.5倍,并且重组菌漆酶诱导时间比原菌株缩短了6 d[12]。近几年,Flammulinavelutipes[13]、Pycnoporuscinnabarinus[14]、Pleurotussajor-caju[15]、Coriolusversicolor[16]和Trametessp. 420[17]等已经实现异源表达,前3个重组菌株漆酶活性比本研究漆酶活性低;后2个重组菌株漆酶活性比本研究漆酶活性高,可能与表达载体选用自身信号肽有关,以及培养方法和条件因素有关。为提高红平菇漆酶基因在毕赤酵母中表达的活性,在今后工作中可采取液体发酵罐并实施高密度发酵;或者通过培养基的优化提高重组毕赤酵母pPICZB-Pd-lac1的高密度表达;还可对重组菌株进行纯化。通过以上方法,红平菇漆酶基因异源表达活性会得到进一步提高,从而实现重组菌株分泌目的蛋白在工业生产中的应用。
2.4 漆酶的分离纯化
将培养6 d后的重组酵母粗酶液进行超滤,SephadexG-75凝胶层析和DEAE-Sepharose阴离子交换层析纯化后经检测为单一条带,大小约为62.5 kDa,与预测红平菇漆酶分子质量大小相似,表明大部分杂蛋白在纯化过程中得到去除(见图6)。图中:泳道1—对照; 泳道2—样本; 泳道3—Marker。
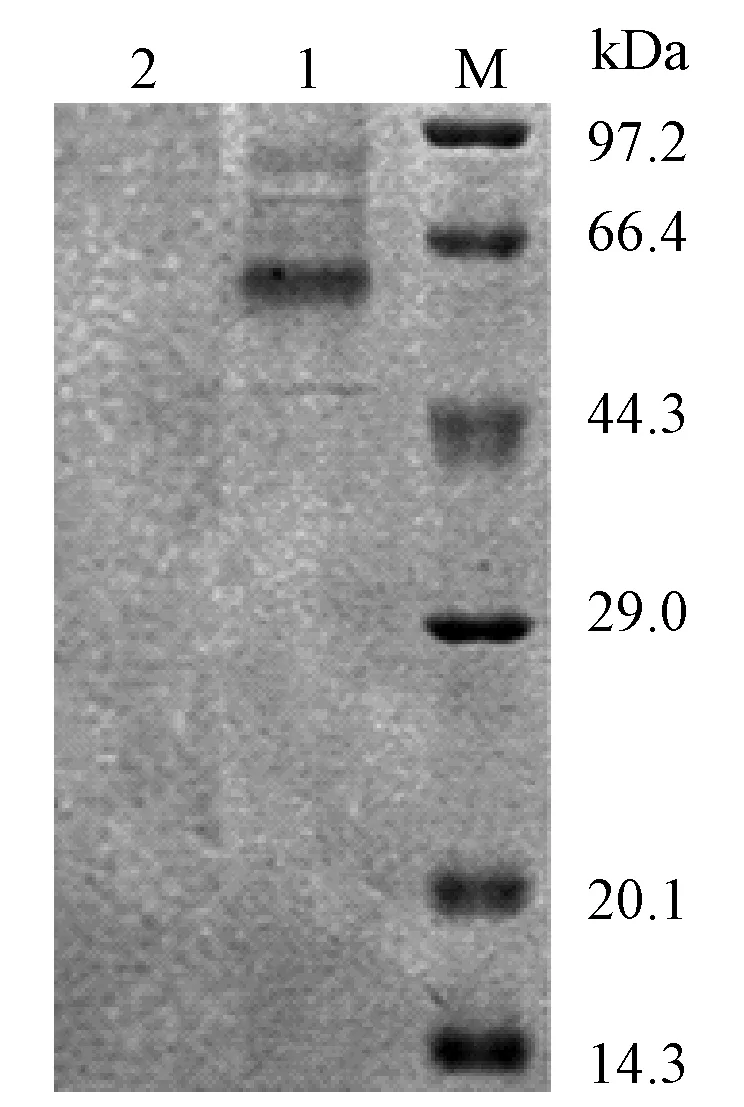
图6 漆酶纯化后的SDS-PAGE电泳图Fig.6 SDS-PAGE of recombinant laccase fron P. pastoris
2.5 重组菌株漆酶Pd-lac1对染料的脱色
分别在4种染料最大吸收峰下检测其吸光度,从而绘制标准曲线。利用BMMY培养基培养Pd-lac1重组毕赤酵母菌株,当漆酶活性到达最大值时,每间隔12 h吸取1 mL粗酶液,6 000 r/min离心4 min,检测重组漆酶对不同染料的脱色效率。反应体系:柠檬酸-磷酸盐缓冲液(pH值3.4)1 mL、粗酶液1.5 mL和不同种染料2.5 mL。反应条件为25 ℃。空白实验不加粗酶液检测对染料脱色效率。所有数据均重复3次取平均值。SMD1168H(pPICZB)作对照。
经过计算,Pd-lac1重组毕赤酵母菌株对SN4R的脱色率在24 h达到最大值,为52.8%,对中性红、甲基橙和孔雀绿在24 h脱色效率达到最大值,分别为14.8%、14.5%和0.24%;而对照组随着时间变化都没有检测到脱色效果,对4种染料脱色效率随时间变化曲线见图7。
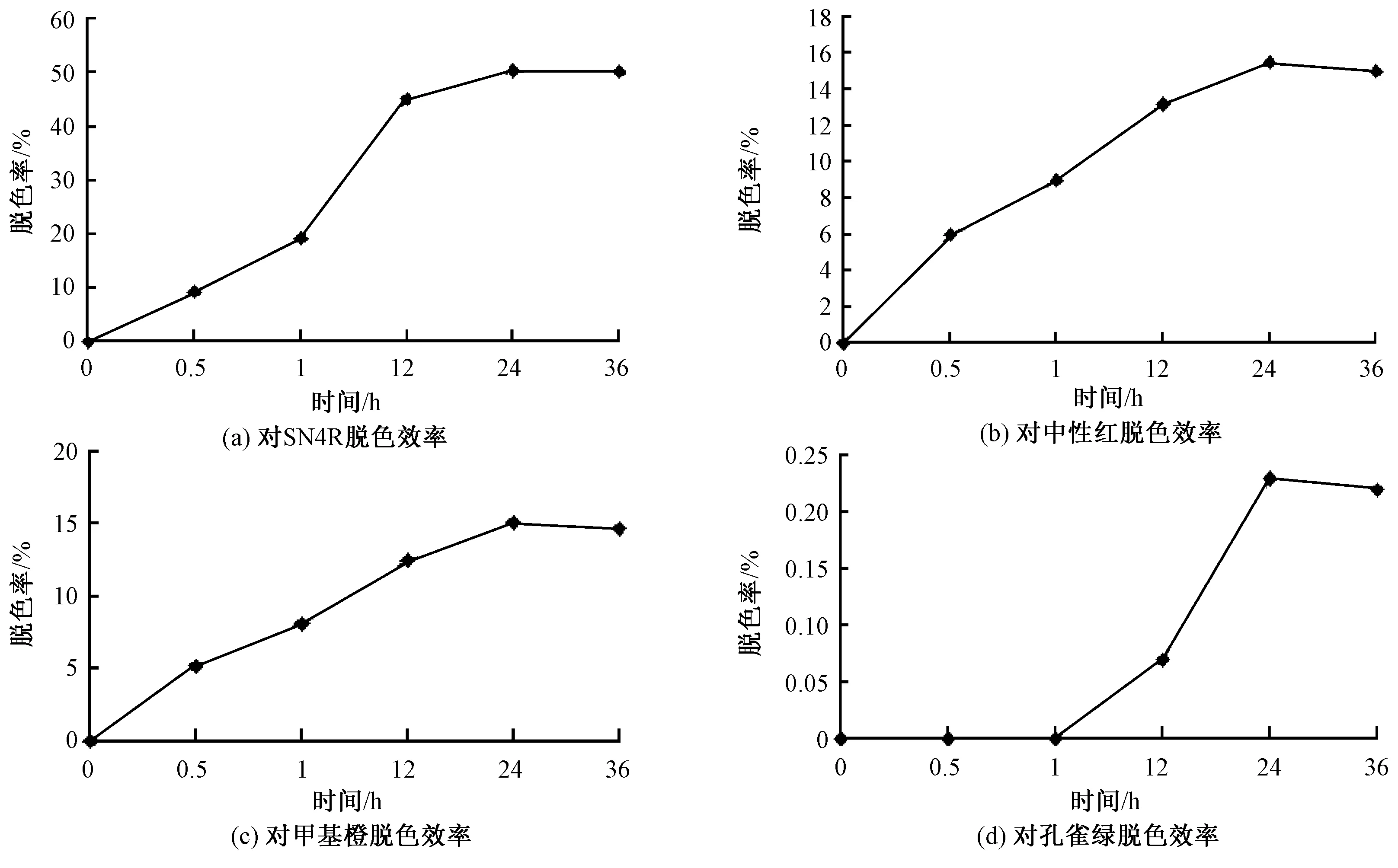
图7 Pd-lac1重组毕赤酵母菌株对染料脱色效率结果Fig.7 Decolorization efficiency of Pd-lac1 recombinant Pichia strains on alizarin red (a), neutral red (b), methyl orange (c), and malachite green (d)
3 结 论
随着人工合成染料的使用量及排放量的逐年上升,环境污染及水体污染问题日趋严重。SN4R、甲基橙、中性红和孔雀绿4种染料在自然水体中很难被生物降解,而且还具有毒性,另外染料还会影响水体的透明度,使水体质量受到严重的破坏。本文研究结果显示,重组菌株漆酶在不需要小分子介体物质的参与情况下,对4种合成染料都具有降解能力。并且对SN4R染料的降解,在24 h脱色率达到53.8%,而原菌株红平菇粗酶液对SN4R染料在24 h时脱色率为49.4%,证明重组菌株漆酶具有较高的脱色效率;而对中性红、甲基橙、孔雀绿3种染料的脱色效率较低,因为SN4R属于蒽醌类染料,化学结构比杂环类中性红、偶氮类甲基橙和三苯基甲烷类孔雀绿相对简单,容易分解,因此重组菌株漆酶对SN4R染料脱色具有较大的应用潜力,进而对含蒽醌类染料废水处理具有更好的应用前景。
总之,本文研究从Pleurotusdjamor克隆了一条漆酶基因,并实现了在毕赤酵母中的异源表达及重组漆酶对不同染料的脱色效率,为进一步研究异源表达优化后染料脱色及重组漆酶纯化奠定了基础。
FZXB
[1] 丛汉卿,徐立,信彩云,等. 植物漆酶的研究进展[J]. 安徽农业科学,2009, 37(18): 8322-8323.
CONG Hanqing, XU Li, XIN Caiyun, et al. Research development of laccase[J]. Journal of Anhui Agri, 2009, 37(18): 8322-8323.
[2] BOURBONNAIS R, PAICE M G, FREIERMUTH B, et al. Reactivities of various mediators and laccases with kraft pulp and lignin model compound[J]. Appl Environ Microbiol , 2000, 66(2): 524-528.
[3] CAI C, CHEN J. Direct electron transfer of glucose oxidase promoted by carbon nanotubes[J]. Biochem Soc Trans , 2004, 332(1): 75-83.
[4] SODEN D M, DOBSON A D W. Differential regulation of laccase gene expressionPleurotussajor-caju[J]. Microbiology, 2001,147: 1755-1763.
[5] XIAO Y Z, TU X M, WANG J, et al. Purification, molecular characterization and reactivity with aromatic compounds of a laccase from basidiomyceteTrametessp. strain AH28-21[J]. Appl Microbiol Biotechnol, 2003,60: 700-707.
[6] KOJIMA Y, TSUKADA Y, KAWAI Y,et al. Cloning, sequence analysis, and expression of ligninolytic phenoloxidase genes of the white-rot basidiomyceteCoriolushirsutus[J]. J Biol Chem, 1990, 265: 15224-15230.
[7] GUO M, LU F P, PU J,et al. Molecular cloning of the cDNA encoding laccase fromTrametesversicolorand heterologous expression inPichiamethanolica[J]. Appl Microbiol Biotechnol, 2005, 69: 178-183.
[8] LARRONDO L F, AVILA M, SALAS L, et al. Heterologous expression of laccase cDNA fromCeriporiopsissubvermisporayields copper-activated apoprotein and complex isoform patterns[J]. Microbiology, 2003,149: 1177-1182.
[9] KILARU S, HOEGGER P J, MAJCHERCZYK A, et al. Expression of laccase genelcc1 inCoprinopsiscinereaunder control of various basidiomycetous promoters[J]. Appl Microbiol Biotechnol, 2006,71: 200-210.
[10] WESENBERG D, KYRIAKIDES I, AGATHOS S N. White-rot fungi and their enzymes for the treatment of industrial dye effluents[J]. Biotechnology Advances, 2003, 22: 161-187.
[11] CUI F J, TAO W Y, XU Z H, et al. Structural analysis of anti-tumor heteropolysaccharide GFPS1b from the cultured mycelia ofGrifolafrondosaGF9801[J]. Bioresource Technology, 2007, 98: 395-401.
[12] 闫洪波,池玉杰. 红平菇木质素降解酶系统漆酶、锰过氧化物酶及木质素过氧化物酶的检测[J]. 林业科学, 2009, 45(12): 154-158.
YAN Hongbo, CHI Yujie. Detection on laccase, manganese peroxidase and lignin peroxidase in ligninolytic enzymes ofPleurotusdjamor[J]. Scientia Silvae Sinicae, 2009, 45(12): 154-158.
[13] 张银波,姜琼,江木兰.金针菇漆酶基因的克隆及其在毕赤酵母中的表达研究[J].微生物学报,2004,44(6): 775-779.
ZHANG Yinbo, JIANG Qiong, JIANG Mulan. Cloning of a laccase gene fromFlammulinavelutipesand study on its expression inPichiapastoris[J]. Acta Microbiologica Sinica, 2004,44(6): 775-779.
[14] LUDOVIC O, ERIC R, SONIA L et al. Molecular cloning of the cDNA encoding laccase fromPycnoporuscinnabarinusI-937 and expression inPichiapastoris[J]. Appl Microbiol Biotechnol, 2000, 76: 1619-1625.
[15] SODEN D M, O′Callaghan J, DOBSON A D. Molecular cloning of a laccase isozyme gene fromPleurotussajor-cajuand expression in the heterologousPichiapastorishost[J]. Biotechnology Advances, 2002,148(12): 4003-4014.
[16] VYAS B R M,MOLITORIS H P.Determination of catecholamines in pharmaceutical formulations using a biosensor modified with a crude extract of fungi laccase (Pleurotusostreatus) [J].Journal of the Brazilian Chemical Society,2003,61(1): 3919-3927.
[17] 周宏敏,洪宇植,肖亚中,等.栓菌漆酶在毕赤酵母中高效表达及重组酶的性质[J]. 生物工程学报,2007, 23(6): 1055-1059.
ZHOU Hongmin, HONG Yuzhi, XIAO Yazhong, et al. High output of aTrametesLaccase inPichiapastorisand characterization of recombinant enzymes[J]. Chinese Journal of Biotechnology, 2007, 23(6): 1055-1059.
猜你喜欢
故事作文·低年级(2023年1期)2023-02-23
小学生(看图说画)(2020年10期)2020-10-29
安徽农学通报(2017年8期)2017-05-12
湖南农业(2016年3期)2016-06-05
农家科技中旬版(2016年12期)2016-04-16
湖北农业科学(2014年10期)2014-09-22
江苏农业科学(2014年4期)2014-07-11
特产研究(2014年4期)2014-04-10
新疆农垦科技(2014年12期)2014-02-28
中国粮油学报(2014年8期)2014-02-06
