
五华三黄鸡线粒体DNA控制区全序列分析
2014-03-30 08:45黄勋和林雅媚李威娜姚琼凤钟福生
嘉应学院学报 2014年2期
黄勋和,林雅媚,李威娜,姚琼凤,钟福生
(嘉应学院生命科学学院,广东梅州514015)
五华三黄鸡是优良的小型肉用地方品种,主要分布于广东省梅州市五华县中部和北部.该品种具有悠久的历史,长期以来在独特的生态环境条件下自然选择形成了独特的生物学特性.在《中国禽类遗传资源》记载的7种广东省地方鸡种中,五华三黄鸡位列其中,也是粤北地区唯一入选的地方鸡[1].1964~1982年五华三黄鸡一直是作为商品出口香港等地.1983年后,五华三黄鸡受到改良商品鸡“石岐杂”大量引入的冲击,饲养数量急剧下降、品种特性受到严重侵蚀.同时,由于该鸡种生长速度慢、繁殖性能低,五华三黄鸡未能得到足够重视,品种保护意识薄弱,饲养规模小而分散[2].目前五华县大部分地区已被外来品种所取代,如胡须鸡、麻鸡、石岐杂鸡等.现在品种纯正的五华三黄鸡数量已经很少,主要分布在五华县的边远山区,作为散养家鸡自给自足.因此,保护濒危的地方特性品种五华三黄鸡刻不容缓.
线粒体DNA控制区(mtDNA Control Region,D-loop)是非编码基因区,在进化过程中受环境的选择压力小,具有进化速率快和母系遗传的特点,常被用于探讨鸟类的起源与分类问题[3~8].国内外对五华三黄鸡线粒体D-loop区的研究还未见报道.本研究通过比较五华三黄鸡两个类群与其他品种的mtDNA控制区D-loop的序列变异,分析它们的亲缘关系,为五华三黄鸡的保种选育提供理论依据.
1 材料与方法
1.1 试验材料
试验用的13只五华三黄鸡取自梅州市丰华有机农业发展公司种禽场.采用拔取背部羽毛的方法,置于95%酒精-70℃保存备用.采用改良的方法提取羽毛基因组,-20℃保存备用[9].
1.2 PCR反应及测序
PCR扩增引物为:PHDL(5'-AGGACTACGGCTTGAAAAGC-3')和PHDH(5'-CATCTTGGCATCTTCAGTGCC-3')[10].反应体系为:1×PCR Buffer(10 mmol/L Tris-HCl pH 8.3,50 mmol/L KCl,1.5 mmol/L MgCl2),4 μL dNTP混合物(各2.5 mmol/L),1 μL连接产物,上下游引物(20 μmol/L)各 0.5 μL,2 U rTaq(TaKaRa),补足ddH2O至50 μL.PCR循环参数为94℃预变性4 min;94℃30 s,50℃50 s,72℃90 s,40个循环;最后72℃延伸5 min,4℃ 保存.扩增产物经AXYGEN AxyPrep DNA凝胶回收试剂盒回收纯化后,由上海立菲生物技术公司广州测序部完成测序.
1.3 序列分析
先利用Bioedit软件对序列数据进行编辑,然后用ClustalX进行对位排列,经人工仔细核查后,再利用DnaSP 4.0提取变异位点.其他相关鸡种的D-loop全长序列从NCBI数据库下载(http://www.ncbi.nlm.nih.gov/).采用DNAMAN进行同源性分析,利用MEGA 5.0统计碱基组成,并计算基于Kimura双参数模型的遗传距离.以原鸡为外群(GenBank登录号: AP003322),构建NJ分子系统发生树.
2 结果与分析
2.1 mtDNA D-loop区序列变异
用Bioedit和ClustalX对原始DNA序列进行对位排列和剪切对齐后,得到五华三黄鸡两个类群——丰华和太和mtDNA控制区序列全长分别为1232、1231bp.与原鸡 (Gallusgallusgallus-AP003322)相比,发现其与五华三黄鸡的丰华类群mtDNA D-loop区序列之间共有14个变异位点,都是碱基转换,包括6次A-G间转换和8次T-C间转换;而五华三黄鸡的太和类群mtDNA D-loop区全序列在第859位点缺失,与原鸡mtDNA D-loop区序列之间共有14个变异位点,其中有一个是缺失,其他的变异位点都是碱基转换,包括5次A-G间转换和8次T-C间转换(图1).

2.2 mtDNA D-loop区碱基组成
本试验测定了五华三黄鸡的两种类群鸡——丰华和太和的mtDNA控制区D-Loop全序列,其长度分布为1232和1231 bp,碱基含量见表1.其中(A+T)含量60.3%~60.4%,明显高于(G+C)含量39.6%.
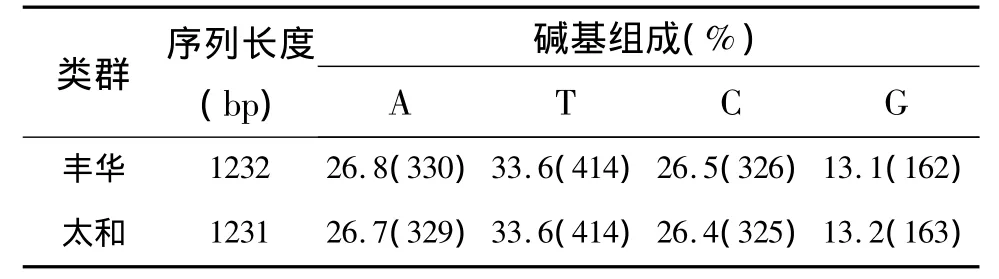
表1 五华三黄鸡D-loop序列碱基组成表
2.3 系统发生树
通过MEGA5.0软件,采用邻接法重建系统发生树,对五华三黄鸡mtDNA D-loop基因进行了遗传进化分析(图2).结果表明,五华三黄鸡两个类群--丰华和太和处于一个分支中,亲缘关系非常近.基于Kimura双参数模型计算丰华和太和的遗传距离为0.001,其中丰华类群与原鸡的遗传距离为0.012,而太和类群与原鸡的遗传距离为0.011.由图1可以清晰的看出,五华三黄鸡与原鸡来自于一个大分支,但亲缘关系较远,与中国红原鸡的亲缘关系最近.

图2 基于mtDNA D-loop序列的20种禽类NJ分子系统发生树
3 讨论
本试验通过PCR反应和测序获得了五华三黄鸡的线粒体控制区D-loop全序列1231 bp和1232 bp,共发现14个变异位点,其中丰华类群都是碱基转换,而太和类群有一个碱基缺失.A+T含量(>60%)明显大于G+C含量(<40%),这也是其他禽类mtDNA D-loop的一个明显特征[11~12].系统进化分析表明五华三黄鸡与中国红原鸡亲缘关系较近;与轱辘鸡亲缘关系较远;与泰国红原鸡和腾冲雪鸡亲缘关系最远.其中太和类群与中国红原鸡的线粒体控制区序列同源性高达100.0%,说明中国红原鸡可能是五华三黄鸡的祖先.鸟类mtDNA控制区的进化速度是2%/ Ma[13],五华三黄鸡丰华类群与原鸡全序列计算的遗传距离(0.012),它们分歧进化的时间约为60万年;五华三黄鸡丰华类群与泰国红原鸡全序列计算的遗传距离(0.013),它们分歧进化的时间约为65万年.
地方鸡经由贸易网络、人类迁移和农业扩张而遍布全世界,同时也带去了它们的基因遗产[8].长期与环境的共进化以及人工选择,形成了大量各具特色的地方品种.有些品种的历史只有几十年,而有些品种则达几千年,如乌骨鸡和杜金鸡.Razafindraibe等(2008)[14]发现马达加斯加岛乡村鸡的一个单倍群可能来自商品鸡品系的基因渗透.6个匈牙利地方鸡的线粒体D-loop分析表明主要来源于印度次大陆,部分可能来源于东南亚、中国和日本[15].宋春红等(2007)[16]探讨了中国6个地方鸡品种的母系起源,推测这些品种分别来自云南、老挝和越南附近地区的红原鸡大陆亚种,基因流是品种群体间遗传分化的主要因素.
五华三黄鸡原产地五华县,在秦汉时期隶属于龙川县,于北宋熙宁四年(1071)建县(长乐县).明宋期间,大量客家先民迁至现在的梅州各县后逐渐定居下来.在此之前,现今五华县辖区内曾先后生活着“百越”、瑶、畲等土著民族,他们曾经是这片土地的主人[17].本研究初步表明在进化关系上五华三黄鸡与中国红原鸡亲缘关系较近,但五华三黄鸡是否起源于中原地区,或是本地鸡种,抑或是融合了中原的混合鸡种,与客家其他地区如江西、福建等地方鸡的亲缘关系等科学问题还有待进一步研究.
[1]陈国宏,王克华,王金玉.中国禽类遗传资源[M].上海:上海科学技术出版社,2004.
[2]钟福生,黄勋和,周彩云,等.五华三黄鸡山地放养条件下肉用性能及内品质的研究[J].嘉应学院学报:自然科学,2013,31(5):67-72.
[3]黄族豪,刘迺发,龙进.从线粒体DNA控制区基因比较石鸡和大石鸡的遗传变异[J].江西农业大学学报,2006,28(3):420-424.
[4]廖承红,王华伟,梁浩,等.文昌鸡线粒体DNA控制区序列遗传多样性分析[J].广东农业科学,2012(12):145-147.
[5]郭亮,张娟,马建宁,等.固原鸡线粒体DNA控制区全序列测定及分析[J].中国家禽,2012,34(1):32-35.
[6]王文涛,陈彬,傅筑荫.两种贵州地方鸡线粒体DNA控制区全序列分析[J].湖北农业科学,2011,50(15):3114-3119.
[7]BERTHOULY-SALAZAR C,ROGNON X,NHU VAN T,et al.Vietnamese chickens:a gate towards Asian genetic diversity[J].BMC Genet,2010(11):53.
[8]MWACHARO J M,BJORNSTAD G,MOBEGI V,et al.Mitochondrial DNA reveals multiple introductions of domestic chicken in East Africa[J].Mol Phylogenet Evol,2011,58(2):374-382.
[9]陈美,黄勋和,王参谋,等.黄嘴白鹭遗传多样性AFLP分析方法的建立[J].厦门大学学报:自然科学版,2008,47 (增刊2):6-10.
[10]FUMIHITO A,MITAKE T,TAKADA M,et al.The genetic link between the Chinese bamboo partridge(Bambusicola thoracica)and the chicken and junglefowls of the genus Gallus[J].Proc Natl Acad Sci USA.1995,92(24): 11053-11056.
[11]李慧芳,朱文奇,杨宁,等.家鸭、媒鸭和野鸭mtDNA D-oop区的遗传变异[J].畜牧兽医学报,2011,42(9):213-219.
[12]张汤杰,李慧芳,陈宽维,等.利用线粒体D-loop区分析家鸭品种遗传多态性与系统进化[J].畜牧兽医学报,2007,38(11):1168-1175.
[13]刘益平,朱庆,曾凡同,等.原鸡线粒体DNA部分序列多态性分析[J].畜牧兽医学报,2004,35(2):134-140.
[14]RAZAFINDRAIBE H,MOBEGI V A,OMMEH S C,et al.Mitochondrial DNA origin of indigenous Malagasy chicken: implications for a functional polymorphism at the Mx gene[J].Ann NY Acad Sci,2008(1149):77-79.
[15]REVAY T,BODZSAR N,MOBEGI V E,et al.Origin of Hungarian indigenous chicken breeds inferred from mitochondrial DNA D-loop sequences[J].Anim Genet,2010,41(5):548-50.
[16]宋春红,陈红菊,马月辉,等.中国6个地方鸡品种的母系起源[J].畜牧兽医学报,2007,38(7):735-740.
[17]刘佐泉.客家历史与传统文化[M].郑州:河南大学出版社,2003.
猜你喜欢
兽医导刊(2022年4期)2023-01-05
当代水产(2022年2期)2022-04-26
当代水产(2021年7期)2021-11-04
中国民间疗法(2021年8期)2021-07-22
海洋通报(2020年5期)2021-01-14
当代工人(2020年9期)2020-08-03
兽医导刊(2019年16期)2019-02-12
中国公路(2017年18期)2018-01-23
水利科技与经济(2017年6期)2017-04-28
家禽科学(2017年3期)2017-02-02
