
美国白蛾核型多角体病毒的研究进展
2014-04-04 00:21吴立华王金昌王洪秀
江西科学 2014年2期
吴立华,靳 亮,王金昌,王洪秀
(1.天津市蓟县林业局,天津301900;2.江西省科学院微生物研究所,江西 南昌330029; 3.江西省农业科学院农业应用微生物研究所,江西 南昌330200)
0 引言
美国白蛾(Hyphantria cunea(Drury))属鳞翅目(Lepidoptera)灯蛾科(Arctiidae),是一种重要的国际性检疫害虫。原产于北美,分布范围在19°~55°N。随着人类活动、现代化交通运输工具的发展,逐渐向世界各地传播。自1979年在我国辽宁省的丹东首次发现以来,美国白蛾强大的繁殖力和适生性,在全国呈蔓延态势;已有50多种林木、果树及农作物受害,严重时有的树木叶子全被吃光,全株枯死,大片林木被毁,给疫区的经济、环境造成了巨大损失和压力[1]。
国内外的许多专家学者,结合美国白蛾疫区的实践经验提出了防治美国白蛾的方法。目前除采用人工采蛹、人工捕蛾、人工网捕及诱杀等人工物理方法以及使用仿生制剂等化学方法降低虫口密度外,较为注重和推崇的是生物防治方法。生物防治方法主要有天敌防治[2]、细菌和病毒防治(苏云金芽孢杆菌(Bt)制剂和美国白蛾核型多角体病毒)[3]以及性信息激素防治[4]。其中杨忠岐[5]等从原产地引进天敌的模式,在国内筛选出寄生率高达83.2%的天敌周氏啮小蜂(Chouioia cunea Yang),在疫区对美国白蛾的自然控制起了关键性的作用,对美国白蛾天敌的研究取得了突破性的进展。同时已开展利用其进行美国白蛾生物防治,现在室内繁殖的技术已很成功,并且取得了较好的防治效果[6~11]。
随着美国白蛾生物防治技术的建立,作为生物杀虫剂的美国白蛾核型多角体病毒虫体繁殖技术也臻于成熟,其作为有效的生物防治手段在各地取得了良好的防治效果[12],但是对于HcNPV的生物学特性及对寄主的致病性及机理等的基础应用研究,报道的尚少,还有待于进一步研究。本文从侵染特点、致病性、流行病学、基因组特点及应用前景等方面综述了美国白蛾核型多角体病毒的研究进展。
1 美国白蛾核型多角体病毒的侵染特点
1.1 感染HcNPV的美国白蛾幼虫的外部症状
美国白蛾幼虫感染美国白蛾核型多角体病毒后,其表现的症状与一般鳞翅目幼虫感染核型多角体病毒的症状相似[1]。初期不表现明显的外部特征。以1.6×106PIB/mL浓度的HcNPV病毒液接毒4龄美国白蛾幼虫,感病3-4 d后出现病症,此时幼虫食欲减退,生长缓慢,发病后幼虫体色逐渐变成浅粉色,行动异常,取食量减少,后期虫体肿胀,体表具光泽,反应呆滞,死亡后,表皮极薄稍触即破,最后虫体液化,呈浅褐色脓液。
1.2 感染HcNPV的美国白蛾幼虫的组织病变过程
取食病毒的幼虫,6-72 h中肠、脂肪体、真皮、气管未见明显病变;96 h中肠、脂肪体、气管个别细胞细胞核出现肿胀;120-144 h各组织细胞核的病变渐渐加重,但中肠病部分细胞肿胀,病变较轻,真皮未见明显病变,至后期,也出现中肠细胞破碎解体[1]。总之,经病毒处理的幼虫,脂肪体储量明显减少,周缘层脂肪体及肌肉与体壁分离;脂肪体裂解成碎片,体壁内表皮、外表皮结构变得松弛,内表皮结构异常,厚度不均匀且表皮细胞明显增厚,正常表皮细胞为单层细胞,而感病试虫增为2~4层,且排列凌乱。精巢未发现病变。
2 美国白蛾核型多角体病毒的致病性研究
有关HcNPV的致病性研究,主要集中在对寄主的毒力、传播和交叉感染等基础应用方面。
2.1 HcNPV对寄主的毒力研究
1979年,Im[13]等在韩国首次发现了美国白蛾核型多角体病毒(HcNPV),毒力测定表明,对2龄、5龄幼虫的LD50分别为8.377×104PIB/mL,4.974×105PIB/mL;在106PIB/ml浓度下,2龄、3龄、4龄、5龄的LT50分别为9.6 d、11.5 d、12 d、17 d,在野外条件下,用6.4×107PIB/mL的病毒感染,3龄时LT50为4.8 d,5龄时为14.2 d,而且第1代幼虫比第2代幼虫更为敏感。可见,随着龄期的增加,LD50增大,LT50延长。而同年,Boucias在室内对美国白蛾质型多角体病毒(Hc-CPV)进行了研究,其结果表明,致死时间与幼虫龄期和病毒的剂量有关,美国白蛾幼虫的死亡率随病毒剂量的增加而增高[14]。1995年对美国白蛾核型多角体病毒作了致病力测定,发现2、3龄幼虫的死亡率远远高于4龄幼虫,在1.5×101~1.5×109PIB/mL喂幼虫,死亡率在90%以上。对美国白蛾核型多角体病毒、斜纹夜蛾核型多角体病毒和从美国白蛾幼虫上分离出的2种颗粒体病毒感染美国白蛾幼虫,在105PIB/mL的剂量下,HcNPV和AcaNPV病毒的死亡率分别是62.5%~100%,70% ~80.5%;颗粒体病毒在5×109PIB/mL剂量下,致病死亡88.1% ~90.0%[15]。
吴戈宇[16]等研究美国白蛾核型多角体病毒(HcNPV)毒力发现,感染美国白蛾2龄幼虫,LC50=1.26×105PIB/mL,感染3龄幼虫LC50= 4.08×105PIB/mL,感染4龄幼虫LC50=5.34× 105PIB/mL。以3×106PIB/mL HcNPV悬液感染美国白蛾2龄幼虫LT50为7.625 d;3龄幼虫LT50为8.625 d;4龄幼虫LT50为9.875 d。以1 ×106PIB/mL HcNPV悬液感染美国白蛾2龄幼虫LT50为9.625 d;3龄幼虫LT50为9.625 d;4龄幼虫LT50为10.625 d。不同龄期幼虫对病毒的敏感性存在一定差异,低龄幼虫较敏感。致死时间随病毒感染浓度增加而减少,同一浓度感染不同虫龄幼虫,病毒感染潜伏期,随虫龄增加而延长。
2.2 HcNPV对寄主的传播和交叉感染研究
1966年在东京大学的林业实验站发生美国白蛾核型多角体病毒流行病。Hukuhara&Namura等随后研究了HcNPV在土壤中的分布[17]。发现在美国白蛾喜食树种附近一定深度的土壤中有HcNPV的存在,而在非喜食树种附近以及道路的土壤内,没有HcNPV的存在,说明土壤中该病毒来源于虫尸。即由宿主危害的树木上掉下来的虫尸中逸散而出于土壤中。另有研究发现,风力可以将HcNPV传播到30 m以外的地方。1977年肯塔基大学的Bouclas&Nordin研究了不同龄期的美国白蛾幼虫对核型多角体病毒NPV和颗粒体病毒GV的敏感性,发现低龄幼虫比老龄幼虫对2种病毒的敏感性差异很大,2龄幼虫的LD50值,后者比前者高100倍,5龄则高1 000倍[18]。1982年,Kunim用病毒糊、病毒悬液、病毒粉与凡士林的混合物处理未产卵的白蛾成虫。用病毒糊处理的雌蛾产卵后,幼虫可以被NPV感染致病;而用病毒悬液、病毒粉与凡士林的混合物处理后,幼虫不感染病毒,而对雌蛾交尾、产卵行为无影响,但是,卵的孵化率下降[19]。Tomita&Iwashita在1982年用HcNPV对其它鳞翅目幼虫作了交叉感染试验,发现红腹灯蛾(Spilosoma subcarnea)对它的敏感性与美国白蛾接近,但它对桑绢野螟(Diaphania pyloalis)、家蚕(Bomyx mori)、分月扇舟蛾(Clostera anastomosis tristis)、桑毛虫(Euproctis silimis)、黄斜带毒蛾(Numenea disparills albofascia)、茶黄毒蛾(Euproctis Psendoconspersa)无致病力[20]。
段彦丽[21]等研究发现用不同浓度的美国白蛾核型多角体病毒亚致死剂量感染美国白蛾4龄和5龄幼虫,并分别收集不同处理的残余带毒试虫进行室内传代饲养。结果表明:病毒对寄主昆虫不仅具有直接致死作用,而且对蛹重和雌虫产卵量均有明显影响,尤其是对亲代、子一代和子二代寄主昆虫。对病毒传播途径的研究证实Hc-NPV可通过多种途径传播到寄主种群中。
不同细胞系对美国白蛾NPV的受纳性是不同的。1987年,Lee H H在斜纹夜蛾(Spodoptera frugiperda)细胞TNM-FH-T内成功复制出美国白蛾核型多角体病毒[22]。并在1988年对其做了温度敏感性试验,在32℃的条件下,12个突变种基本上都不能形成多角体。
2.3 HcNPV制剂对美国白蛾防治研究
我国也对应用病毒制剂防治美国白蛾进行了研究:刘岱岳[23]等在1985年在陕西省用美国白蛾核型多角体病毒制剂107PIB/mL进行飞机常量低空喷雾防治美国白蛾。喷雾10 d以后调查,地面定点调查防治效果73.4%,树梢定点调查防治效果为84.6%。赵海燕[24]等在2010年在青岛城阳区开展了美国白蛾核型多角体病毒的林间防效试验;药后14 d,青岛农业大学害虫生物防治技术研究所提供HcNPV病毒液浓度分别为5×106PIB/mL、3.3×106PIB/mL和2.5×106PIB/mL,其校正防效分别为90.41%、87.67%、90.41%。
另外,HcNPV与其它细菌或者荧光染料复配后,杀虫效果明显提高。段彦丽[25]等发现与美国白蛾核多角体病毒单独使用相比,HcNPV与Bt合剂使LT50缩短0.5-2.1 d,缩短了美国白蛾幼虫死亡时间,加快美国白蛾核多角体病毒致死速度。杨唯一[26]等研究了3种荧光增白剂对Hc-NPV的增效作用,结果发现0.5%的VBL(荧光增白剂)和1%的BA(荧光增白剂)对HcNPV的增效作用明显,可作为HcNPV的增效剂加入到制剂中。
3 美国白蛾核型多角体病毒的分子生物学研究
近几十年来病毒分子生物学得到了蓬勃的发展,昆虫病毒的研究也不例外,但是HcNPV的分子生物学研究与其它NPV相比才刚刚起步,但也积累了一些研究成果。1998年,应用基因工程技术,将苏云金杆菌库尔斯塔克亚种的HD-1菌株中的杀虫蛋白基因(ICP),导入到美国白蛾核型多角体病毒中,制成了复合病毒制剂,命名为ICP-HcNPV杀虫剂。试验证明,Bt的杀虫基因通过复合病毒在昆虫细胞中以毒素蛋白的23形式表达出来,表现出很高的杀虫活性[22]。Hyung-Hoan Lee[27]等用BamHI和SmaI 2种限制性外切酶对HcNPV基因组作酶切分析,绘制了HcNPV基因组物理图。翌年用质粒载体 pUC8和pBR322克隆了部分基因组EcoRI酶切片段[28]。同年克隆和定位了多角体蛋白基因 Polyhedrin[29]。
贡成良[30]等研究报道美国白蛾(Hy-phantria cunea)核型多角体病毒(HcNPV)CP基因的核苷酸序列及蛋白质的一级结构特征;翌年测定了美国白蛾核型多角体病毒几丁质酶基因核苷酸序列[31];2000年通过对HcNPV半胱氨酸蛋白酶、几丁质酶基因失活分析研究提示:CP、ChiA两基因为病毒复制非必需基因,它们的失活不影响病毒的复制与多角体的形成,但感染细胞的存活时间比HcNPV、HcNPVPTTH+感染的多2 d。推测CP、ChiA两基因失活后,可延长细胞持续表达外源基因的时间[32]。曹广力等进行了超氧化物歧化酶基因SOD和抑制细胞凋亡基因p35的克隆和序列分析[33,34]。
近年来,Ikda[35]等研究了美国白蛾核型多角体病毒的iap基因发现:在HcNPV感染SpIm细胞的过程中,Hycu-IAP3对于阻止细胞凋亡来说是必要的。2006年,Ikda[36]等完成美国白蛾全基因组序列测定,基因组DNA结构如图1所示。HcNPV基因组序列全长132959 bp,G+C含量45.1%,有148个开放阅读框(ORFs),编码50多种多肽。基于系统学上的亲缘关系以及拥有group I NPVs中的特有基因如 gp64、ie-2、ptp-1等,推断 HcNPV属于 group I NPVs,并且与 Cf MNPV或Op MNPV非常相似。另外,美国白蛾核型多角体病毒基因组拥有6个独特ORFs,他们分别是 ORF 71、ORF 41、ORF 99、ORF 100、ORF 121、ORF 148。这6个ORFs在核苷酸序列基因库中没有找到任何可识别的同源性序列。这些序列的功能虽然还不清楚,但是可以作为美国白蛾核型多角体病毒的特异性检测序列,应用到美国白蛾NPV的流行病毒学研究。Shirata[37]等研究发现,当家蚕核型多角体病毒(BmNPV)和Hc-NPV共同感染家蚕细胞时,HcNPV的ep32编码的蛋白诱导家蚕细胞的球形蛋白合成量下降,限制了家蚕核型多角体病毒扩增。这一研究结果为理解细胞对可感染病毒的抗性机制提供了一种模式。
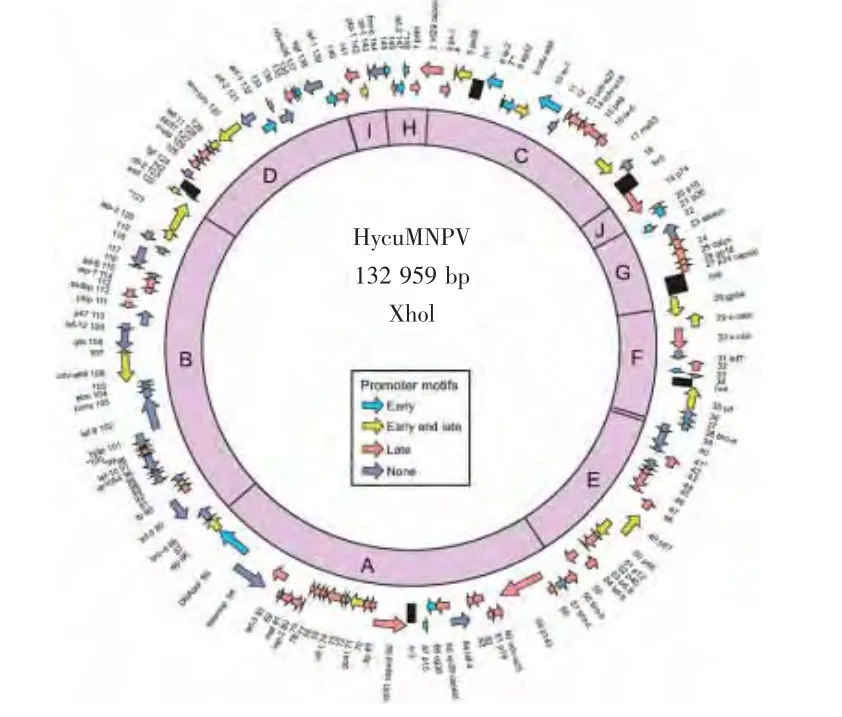
图1 HycuMNPV基因组DNA结构和限制性内切酶XhoⅠ的物理图谱(摘自Motoko Ikeda et al.,2006)
4 美国白蛾核型多角体病毒杀虫剂的产业化发展
HcNPV作为一种重要的病原微生物和生物杀虫剂,不仅具有专一性强、对环境和人畜安全、不杀伤天敌等优点,而且在其应用后能在自然界中长期存在,从而引起病毒流行病的大发生,可在较长时间内自然控制害虫种群密度的增长,在害虫的持续控制研究中发挥着重要的作用。HcNPV对防治美国白蛾幼虫具有上述优点,可有效降低化学药剂对环境的污染,可以大面积应用[5~7]。
但病毒作为杀虫剂在应用之前,首先要解决大量增殖问题。目前,采用活虫体进行增殖依然是获得大量病毒的主要途径[38],虫龄和接种浓度(剂量)是影响病毒大量增殖的主要因素[39,40]。郑桂玲[41]等对美国白蛾龄期接种剂量及其组合对该HcNPV产量的影响幼虫进行了研究,筛选出适合大量增殖的最佳条件为:在室内大量增殖HcNPV时,以1.48×106OBs/头的剂量饲喂5龄和6龄幼虫均可以获得最高的病毒产量,百头幼虫的病毒总产量可达到1.55×1011OBs和1.50 ×1011OBs,而单头幼虫的病毒产量最高可达2.319 ×109OBs,这为该病毒的大量增殖和病毒杀虫剂的生产提供理论依据。中国林业科学研究院应用微生物研究室在美国白蛾幼虫的人工大量饲养、病毒的工厂化生产以及病毒的田间应用技术方面已获得成功经验。
综上所述,从侵染特点、致病性、流行病学、基因组特点及应用前景等方面综述了美国白蛾核型多角体病毒的研究进展。随着HcNPV全基因组序列测序的完成,相关的分子生物学的研究将会蓬勃向前推进。另外,随着美国白蛾幼虫的规模化饲养技术(包括种虫车间、健康幼虫饲养车间、病毒感染增殖车间、制剂加工车间的设计和智能化;优化幼虫感染技术,稳定单位病毒产量病毒技术)的成熟,HcNPV杀虫剂虫将从实验室走向大田,这将对科研成果的推广起到良好的促进作用。
[1] 段彦丽.美国白蛾NPV和Bt混合致病机理及其对寄主种群持续控制作用[D].北京:中国林业科学研究院,2008.
[2] 杨秀卿,魏建荣,杨忠岐.大连地区美国白蛾寄生性天敌昆虫[J].中国生物防治,2001,17(1):40-42.
[3] 陈 颖,但汉斌,魏雪生,等.化学Bt-15A3防治美国白蛾的试验[J].天津农学院学报,2003,(1):24-26.
[4] Valentyuk E I,Derevyanko N M,Gensitskii I P.Polistes wasps and the fall webworm[J].Zashchita Rastenii Moskva,1989,(7):38.
[5] Bishop D H L.Genetic engineering of microbes:virus insecticides-a case study In:Darby,G.K.Hunter,P.A.,Russel,A.D.(Eds),50 Years of Microbials[M].England Cambridge:Cambridge University Press,1995:249-277.
[6] Burden J P,Hails R S,Windass J D,et al.Infectivity,speed of Kill,and productivity of a baculovirus expressing the itch mite toxin txp-1 in second and fourth instar Larvae of Trichoplusia ni[J].Journal of Invertebrate Pathology,2000,75(3):226-236.
[7] Cory J S.Field trials of genetically improveed baculovirus insecticide[J].Nature,1994,370:138-140.
[8] McCutchen B F,Choudary P V,Crenshaw R,et al.Development of a recombinant baculovirus expressing an insect-sensitive:potential for pest control[J].Biol Technology,1991,9:848-852.
[9] Keddie B A,Aponte G W,Volkman L E.The pathway of infection of Autographa californica nuclear polyhedrosis virus in an insect host[J].Science,1989,243: 1728-1730.
[10]Pringle C R.Virus taxonomy-1999.The universal system of virus taxonomy,updated to include the new proposals ratified by the International Committee on Taxonomy of Viruses during 1998[J].Archives of Virology,1999,144(2):421-429.
[11]Black B C,B rennan L A,Dierks P M,et al.Commercialization of baculoviral insecticides in baculoviruses[M].New York:Edited by Lois,K.Miller.Plenum Press,1997.
[12]杨忠岐,张永安.重大外来入侵害虫—美国白蛾生物防治技术研究[J].昆虫知识,2007,44(4):72-80.
[13]Im D J,Hyun J S,Paik W H,et al.Studies on the nature and pathogenicity of nuclear polyhedrosis virus of the fall webworm,Hyphantria cunea(Drury)[J].Korean Journal of Plant Protection,1979,18(1):1-10.
[14]Boucias D S,Nordin G L.Susceptibility of Hyphantria cunea to a cytoplasmic polyhedrosis virus[J].Journal of the Kansas Entomological Society,1979,52(4):641-647.
[15]Pritchett D W,Young S Y.Efficacy of baculoviruses against field population of fall webworm,Hyphantria cunea[J].Journal of the Georgia Entomollogical Society,1980,15(3):332-336.
[16]吴戈宇,刘玉书,李 银,等.美国白蛾核型多角体病毒毒力的生物测定[C].华中昆虫研究,2008:144-148.
[17]Hukuhara T,Namura H.Distribution of a nuclear-polyhedrosis virus of the fall webworm,hyphantria cunea,in soil[J].Journal of Invertebrate Pathology,1972,19: 308-316.
[18]Boucias D G,Nordin G L.Interinstar susceptibility of the fall webworm,hyphantria cunea to its nuclearpolyhedrosis and granulosis viruses[J].Journal of Invertebrate Pathology,1977,30:68-75.
[19]Kunimi Y.Transovum transmission of a nuclear polyhedrosis virus of the fall webworm,Hyphantria cunea DRURY(Lepidoptera:Arctiidae)[J].Applied Entomology and Zoology,1982,17(3):410-417.
[20]Tomita K.Cross-transmission of the granulosis virus of hyphantria cunea drury(Lepidoptera:Arctiidae),to other lepidopterous insect species[J].Journal of Applied Entomology and Zoology,1982,26(4):224-227.
[21]段彦丽,曲良建,王玉珠,等.美国白蛾核型多角体病毒传播途径及对寄主的持续作用[J].林业科学,2009,45(6):83-86.
[22]Lee H H,Lee M K,Cho I H,et al.Location and clo-ning of the polyhedrin gene of Hyphantria cunea nuclear polyhedrosis virus[J].Journal of the Korean Society Virology,1991,21(1):25-34.
[23]刘岱岳,居蜀生.美国白蛾病毒的繁殖与飞机喷施[J].生物防治通报,1986,2(2):74.
[24]赵海燕,王艳士,修玉义,等.美国白蛾核型多角体病毒林间药效试验[J].中国森林病虫,2012,31 (1):38-39.
[25]段彦丽,陶万强,曲良建,等.HcNPV和Bt复配对美国白蛾的致病性[J].中国生物防治,2008,24(3): 223-238.
[26]杨唯一,张永安,唐 明,等.三种荧光增白剂对Hc-NPV增效作用研究[J].林业科学研究,2009,22 (5):736-73.
[27] Lee H H,Lee H J.Reseach map of the geneme of Hyphantria cunea nuclear polyhedrosis virus[J].Kon Kun Journal of Genetic Engineering,1991,4(0):9-56.
[28] Lee H H,Kim J W,Kim H K,et al.Cloning of the Hyphantria cunea Nuclear Polyhedrosis Virus Partial EcoRI Genome DNA Fragments in Plasmid Vectors pUC8 and pBR322[J].J.of Kor.Soc.of Virology,1991,21(1):35-40.
[29]Flipsen J T,Mans R M,Kleefsman A W,et al.Deletion of the baculovirus ecdysteroid UDP-glucosyltransferase gene induces early degeneration of Malpighian tubules in infected insects[J].Journal of Virology,1995,69 (7):4529-4532.
[30]贡成良,小林淳,宫岛成寿,等.HcNPV半胱氨酸蛋白酶基因的核苷酸序列研究[J].生物化学与生物物理学报,1998,30(3):307-310.
[31]贡成良,小林淳,宫岛成寿.美国白蛾核型多角体病毒几丁质酶基因核苷酸序列研究[J].病毒学报,1999,15(3):260-269.
[32]贡成良,小林淳,金 伟,等.HcNPV半胱氨酸蛋白酶、几丁质酶基因失活分析[J].生物化学与生物物理学报,2000,32(2):187-191.
[33]曹广力,薛仁宇,朱越雄,等.美国白蛾多角体病毒超氧化物歧化酶基因的序列分析和表达[J].微生物学报,2001,41(2):173-180.
[34]曹广力,薛仁宇,朱越雄,等.美国白蛾核型多角体病毒p35基因的克隆及序列分析[J].昆虫学报,2002,45(6):711-716.
[35] Ikeda M,Yanagimoto K,Kobayashi M.Identification and functional analysis of Hyphantria cunea nucleopolyhedrovirus iap genes[J].Virology,2004,321:359-371.
[36]Ikeda M,Shikata M,Shirata N,et al.Gene organization and complete sequence of the Hyphantria cunea nucleopolyhedrovirus genome[J].J Gen Virol,2006,87: 2549-2562.
[37] Shirata N,Ikeda M,Kobayashi M.Identification of a Hyphantria cunea nucleopolyhedrovirus(NPV)gene that is involved in global protein synthesis shutdown and restricted Bombyx mori NPV multiplication in aB.moricell line[J].Virology,2010,398:149-157.
[38]陶万强,潘彦平,王金利,等.杨扇舟蛾颗粒体病毒室内增殖的研究[J].中国森林病虫,2009,28(3):9-10,18.
[39]Grzywacz D,Jones K A,Moawad G,et al.The in vivo production of Spodoptera littoralis nuclear polyhedrosia virus[J].Jounal of Virological Methods,1998,71(1): 115-122.
[40]李长友,张履鸿,李国勋,等.甘蓝夜蛾核型多角体病毒的大量增殖[J].东北农业大学学报,1995,26 (3):247-250.
[41]郑桂玲,周洪旭,耿以龙,等.美国白蛾核型多角体病毒室内大量增殖的研究[J].中国森林病虫,2011,30(5):36-38,41.
猜你喜欢
宁夏医学杂志(2020年3期)2021-01-21
现代园艺(2018年3期)2018-02-10
现代园艺(2017年19期)2018-01-19
现代园艺(2017年23期)2018-01-18
金色少年(奇趣科普)(2017年4期)2017-06-05
农村农业农民·B版(2016年7期)2016-10-21
山东林业科技(2016年5期)2016-07-05
哈尔滨医药(2015年2期)2015-12-01
中国当代医药(2015年9期)2015-03-01
应用海洋学学报(2014年3期)2014-11-22
