
中国红树林北缘区秋茄树种群空间分布格局和结构特征分析
2014-04-09 11:45何东进肖石红李晓景蔡金标游巍斌
植物资源与环境学报 2014年2期
何东进, 肖石红, 李晓景, 王 韧, 蔡金标, 游巍斌
(1. 福建农林大学, 福建 福州 350002; 2. 福鼎市林业局, 福建 福鼎 355200; 3. 霞浦县林业局, 福建 霞浦 355100)
秋茄树〔Kandeliacandel(Linn.) Druce〕为红树科(Rhizophoraceae)秋茄树属(KandeliaWight et Arn.)植物,广泛分布于广东、广西、福建和台湾等地[1]。由于秋茄树群落多处于经常性受到海水淹滞的潮间带,使其兼具陆地植物群落和海洋植物群落的双重特性,在自然生态平衡中有特殊作用[2-5]。红树林群落除了在湿地生态系统中具有促进土壤沉积物形成、过滤有机物和污染物以及净化水质等重要作用外,还有抵抗潮汐和洪水冲击、减缓风浪、调节水流以及保护堤岸等功能[6]。福建省福鼎市是中国天然红树林的北缘区,由于纬度高、温度低等原因,与中国其他红树林主要产区的生境条件存在明显差异[7-8]。因此,研究北缘区红树林的形成过程和生理生态特征具有特别重要的意义。
空间分布格局是一个种群内个体间在空间中的配置模式或分布规律,能反映种群个体在水平空间上的相互关系[9]。种群分布格局的形成与物种生理生态学特性、种内和种间关系、生物和非生物因素干扰以及生境条件密切相关[10-12]。研究种群空间分布格局不仅能了解某个物种的空间分布特点,还能通过物种不同阶段的格局变化寻求其变化规律及其关联性,了解种群进化过程以及种内和种间关系的动态变化等[13-14]。生命表分析是研究种群动态特征的重要方法,通过编制种群生命表能掌握影响种群数量变化的主要因素、判断种群动态发展趋势、分析物种生命过程中重要的死亡威胁,对进一步估算种群趋势和组建预测模式等都能起到一定的指示作用。
闽东滨海湿地不同演替阶段秋茄树林均遭严重干扰,种植历史超过150 a的近自然群落逐渐凸显生境丧失问题,与天然林或其他秋茄树林产区相比,研究区内秋茄树人工林长势不佳[8,15]。对闽东滨海湿地不同滩位的秋茄树林空间分布格局及生命过程进行研究能够了解闽东秋茄树林的动态变化特征,一方面能更有效地保护秋茄树天然林,恢复适宜的生态环境,减缓自然衰退过程;另一方面对比天然林群落自然进化机制,能够为人工林种植策略的制定提供理论依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区域的地理、气候和海洋等基本概况参见文献[15]。
1.2 研究方法
1.2.1 样地设置与径级划分 在全面勘查基础上,选择福鼎市前歧镇秋茄树天然林和点头镇秋茄树人工林2个具有代表性的地段分别设置3块样地,每个样地外滩和内滩面积各占50%,样地间沿着海岸线方向间隔30 m,样地面积20 m×20 m,共6块样地;调查记录样地内人为干扰状况、郁闭度、群落年龄和经纬度等基础数据;对样地内的乔木层种类进行每木检尺,测定树高、胸径、冠幅和基径。
依据海水到达区域的差异将秋茄树个体划分至不同滩位:高潮时海水到达的地方为内滩,低潮时海水到达的地方为外滩。对于立木级的划分方法,不同学者提出了多个标准[16-18];本研究则根据秋茄树的生活史特点,按照基径(D)将秋茄树个体划分为13个径级: D<2 cm为Ⅰ级,随后以4 cm为径阶进行划分, 2 cm≤D<6 cm为Ⅱ级、6 cm≤D<10 cm为Ⅲ级、……、D≥46 cm为ⅩⅢ级(上限排除法)。参照秋茄树的径级将其种群划分为幼苗群(Ⅰ级)、小树群(Ⅱ和Ⅲ级)、大树群(Ⅳ~Ⅶ级)和老树群(Ⅷ级及以上)4个发育阶段,分别测算聚集度指标。

1.2.3 生命过程研究方法 以上述按照径阶等级划分的秋茄树天然林个体的13个径级替代相应的龄级,即Ⅰ级为龄级1,Ⅱ级为龄级2,以此类推,以“空间替代时间”的方法编制秋茄树天然林种群的静态生命表,分析其生命过程[19-21]。计算以下各项参数:lx(x龄级开始时的标准存活数)、ax(x龄级的现有存活数)、dx(从x到x+1龄级间隔期的标准死亡数)、qx(从x到x+1龄级的期间死亡率)、Lx(从x到x+1龄级间隔期的标准存活个体数)、Tx(x龄级及以上龄级的标准个体总数)、ex(进入x龄级个体的平均期望寿命)、Kx(消失率)、px(x龄级的存活率)、Sx(生存率)和Fx(累计死亡率)。并绘制种群期间死亡率、消失率、生存率和累计死亡率曲线以及存活曲线。
2 结果和分析
2.1 不同滩位秋茄树天然林和人工林的高度结构分析
不同滩位秋茄树天然林及人工林的高度分布状况能直观地显示不同高度个体在群落垂直结构中的地位和作用。由表1可见:位于内滩的秋茄树天然林株高主要集中于0.0~0.5和1.5~2.0 m,株数合计有353株,占总数的67.0%;位于外滩的天然林株高也有类似现象,株高0.0~0.5和1.5~2.0 m 的株数合计有236株,占总数的71.1%。由于秋茄树人工林无幼苗存在,故株高0.0~0.5 m的株数为0,株高主要集中于1.5~2.0 m;位于内滩的秋茄树人工林株高1.5~2.0 m 的植株有671株,占总数的87.7%;位于外滩的人工林株高1.5~2.0 m的植株有403株,占总数的86.9%。
由表1可以看出:秋茄树天然林及人工林不同滩位间的高度结构相似,但天然林和人工林的高度结构却不同。由于天然林中存有大量幼苗,天然林内株高0.0~0.5 m的个体所占比例较大,株高1.5~2.0 m的个体也较多,相对于人工林而言,天然林的高度结构分布较为均匀。而人工林中株高1.5~2.0 m的个体数量超过80.0%,株高2.0 m以上的个体极少且无幼苗,显示秋茄树人工林的高度结构极不合理。

表1 不同滩位秋茄树天然林和人工林的高度结构
2.2 不同滩位秋茄树天然林和人工林的径级结构分析
不同滩位秋茄树天然林和人工林的径级结构见表2。由表2可见:外滩和内滩的秋茄树天然林均呈现随径级增大个体数量减少的趋势;而人工林外滩和内滩的个体数量变化均存在1个明显的峰值,即径级Ⅲ的个体数量均最多,且不存在Ⅸ级以上个体。
从表2中还可见:秋茄树天然林和人工林的径级分布规律明显不同。秋茄树天然林各径级个体数随径级增大均匀减少,符合发育良好的天然林径级结构的普遍规律;而人工林各径级个体数近似正态分布,处于径级Ⅲ的个体数量达到峰值,在群落中占有优势地位。
2.3 秋茄树天然林和人工林的空间分布格局分析

表2 不同滩位秋茄树天然林和人工林的径级结构

表3 秋茄树天然林和人工林不同发育阶段的空间分布格局1)
2.4 秋茄树天然林种群的生命过程分析
由于秋茄树人工林均为人为一次性造林,且栽种时间较短,故仅以秋茄树天然林为对象进行生命过程分析。依据“空间替代时间”方法,依据秋茄树天然林不同径级的个体数量编制静态生命表,作为秋茄树天然林多个世代重叠的某一特定时间进行分析。为了避免生命表中出现死亡率为负值的情况,采用匀滑技术对数据进行处理,其中x龄级的现有存活数ax经匀滑后得ax*。秋茄树天然林种群的静态生命表见表4。以秋茄树天然林种群的静态生命表为基础,进一步进行生存分析,其种群期间死亡率、消失率、生存率和累计死亡率曲线以及存活曲线分别见图1、图2和图3。
由图1可以看出:秋茄树天然林种群的期间死亡率与消失率的变化趋势基本趋于一致,二者在龄级1和7达到最大,在这2个龄级内有一半左右的个体死亡;除这2个龄级外,其他龄级的期间死亡率和消失率都不大;在龄级8~12等5个阶段,期间死亡率均维持在0.1以内,且变化幅度较小。从生存率与累计死亡率曲线(图2)中可以看出:秋茄树天然林个体的生存率单调下降、累计死亡率单调上升,二者互补;秋茄树天然林仅在龄级1时个体生存率高于累计死亡率,从龄级2开始均表现为累计死亡率高于生存率。
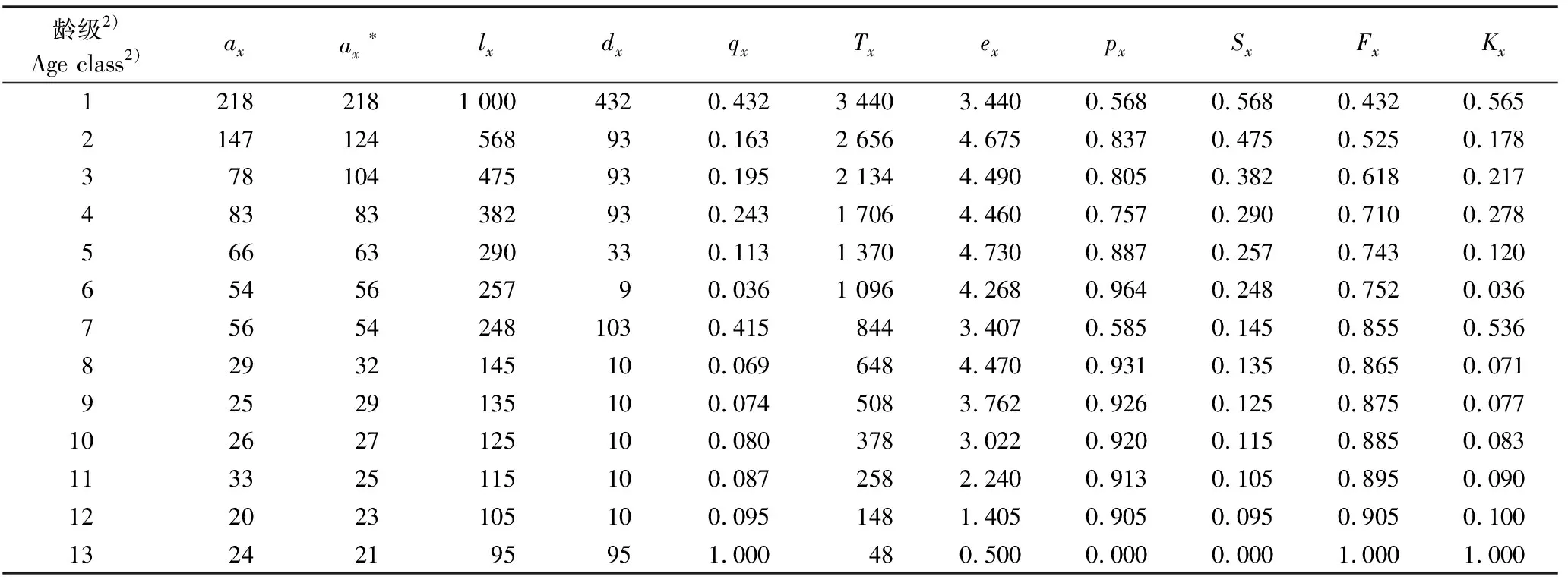
表4 秋茄树天然林种群的静态生命表1)

—●—: 期间死亡率 Interval mortality rate;

—○—: 生存率Survival rate;
种群的存活曲线分为Deevey-Ⅰ型、Deevey-Ⅱ型和Deevey-Ⅲ型3种类型,可以采用指数方程式Nx=N0e-bx描述Deevey-Ⅱ型存活曲线,采用幂指数方程式Nx=N0x-b描述Deevey-Ⅲ型存活曲线。采用Deevey-Ⅱ型和Deevey-Ⅲ型2种模型对秋茄树天然林种群的存活曲线进行检验:Nx=644.07e-0.203 1x,R2=0.891 9;Nx=1 003.5x-1.075 3,R2=0.965 4。通过比较两者的相关系数,可以认为秋茄树天然林种群的存活曲线更趋向于Deevey-Ⅲ型存活曲线。由此可见,秋茄树天然林种群整体的数量动态变化趋势及结构特征为:在幼苗时期个体死亡率较高,随个体生长至成树时期,种群内个体数量趋于稳定。

用径级替代龄级Age class is replaced by diameter class.
3 讨论和结论
秋茄树天然林经过长期演替,在高度结构上已初步形成了一定数量的适应于闽东滨海湿地特殊环境的个体,即在秋茄树天然林中株高0.0~0.5和1.5~2.0 m的个体占较大比例。与天然林相比,秋茄树人工林的高度结构不甚合理,缺乏大量幼苗以及株高超过2.0 m的高大植株,成为制约秋茄树人工林健康演替的重要因子,秋茄树人工林的径级结构也验证了该问题。另外,秋茄树天然林中径级Ⅱ~Ⅲ和Ⅶ~Ⅷ 2个阶段的个体数量变化较大,死亡率分别高达46.9%和48.2%,依据“空间替代时间”方法,可以将这2个阶段看作秋茄树天然林种群生长发育的重要过渡阶段;其中,前者可视为秋茄树幼苗向小树的过渡阶段,后者可视为秋茄树小树向成树的过渡阶段。而调查区内秋茄树人工林的林分结构不合理,作为优势种的秋茄树大部分个体仅处于小树阶段,群落结构不稳定,不能充分发挥其生态作用。
从空间分布格局来看,秋茄树天然林除在幼苗发育阶段为聚集分布格局外,其他发育阶段都属于随机分布格局,这主要与秋茄树本身特殊的生殖繁育方式有关。通常,秋茄树幼苗先在母体上生活一段时间,在母体上发育到一定程度后幼苗从母株脱落并落入湿地淤泥中逐渐发育成熟,秋茄树的这种特殊繁殖方式提高了其后代在滨海湿地特殊环境下的生存能力。从高度结构和径级结构中也可推断,秋茄树是一种典型的依靠r对策生存的物种,每一个重要的发育过渡阶段都将依靠“高死亡率换取竞争力”的方式存活[22]。秋茄树幼苗的“胎生”方式以及r对策使得大量幼苗分布在母树四周,形成聚集分布。而从幼苗过渡到小树阶段后,一方面研究区属滨海湿地地带,光照、地势和水分等主要生境因子差别不大;另一方面,秋茄树本身生境偏好相对不明显,因此小树群、大树群和老树群随机分布。在秋茄树人工林中,小树群呈均匀分布,大树群呈聚集分布;一般情况下,自然界中绝对呈均匀分布的种群极为罕见,研究区内秋茄树人工林之所以呈均匀分布且小树群占极大比例,其原因可能为过度人工密植,经过长期演替仍未恢复自然状态;而少量小树个体生长为大树个体,随着株高和径级的增大对空间和养分的竞争加剧,呈现聚集分布状态。
从秋茄树天然林种群的静态生命表与存活曲线、期间死亡率及消失率曲线可以看出:秋茄树天然林种群中幼苗丰富但期间死亡率较高,成树期间死亡率相对较低且生长稳定。在整个生活史中,秋茄树个体需要穿越2个死亡率较高的阶段,即幼苗发育成小树和小树发育成成树的阶段,这一点与其径级结构分布一致。由于秋茄树采用的r对策生存方式,大量的幼苗通过环境的过滤和筛选才能进入下一阶段,随着个体进一步发育,由于空间和资源的有限性以及种群内部的自疏现象,必然阻止大量个体进入生殖生长阶段。否则,秋茄树较强的繁育幼苗能力加之数量较多的具有繁殖能力的个体将会加剧种群内部对资源的竞争,从而破坏现有的种群结构。
对不同滩位秋茄树天然林及人工林的研究结果表明:秋茄树人工林及天然林的高度及径级结构在内滩和外滩差异不大。外滩频繁受到潮汐和海浪冲刷,秋茄树植株数量相对少于内滩。但内滩与外滩秋茄树的高度结构与径级结构中各等级比例相似,说明内滩和外滩间的水分和盐分差异对秋茄树生长发育的影响并不显著,也侧面反映了秋茄树本身生境偏好相对不明显。与秋茄树人工林相比,秋茄树天然林的林分密度和郁闭度都小于人工林;在相同的调查面积内,天然林和人工林的总株数分别为859和1 229,林分郁闭度分别约为70%和90%,人工林的过度人工密植是出现这一状况的主要原因。但是,实地调查结果表明:秋茄树天然林与人工林群落的物种多样性均较差。在秋茄树的其他分布地,秋茄树种群形成后能够自然扩散海莲〔Bruguierasexangula(Lour.) Poir.〕、桐花树〔Aegicerascorniculatum(Linn.) Blanco〕和木榄〔Bruguieragymnorrhiza(Linn.) Savigny〕等红树林树种,然而这些树种在本研究区域中出现频率相对较少,而且在本研究区域中秋茄树本身的长势也明显比其他分布地差,如闽东秋茄树人工林平均株高为1.79 m,远低于深圳福田相同龄级秋茄树人工林平均株高2.89 m[23],也明显低于海南琼山12年生秋茄树人工群落(平均高度2.4~3.0 m)[24]。究其原因主要是本研究区域纬度高、温度低,是中国秋茄树红树林天然林生长的北缘区,对秋茄树而言这一区域的生境条件并非最佳。
参考文献:
[1] 林 鹏. 中国红树林生态系统[M]. 北京: 科学出版社, 1997: 42-66.
[2] ZHANG Y H, WANG W Q, WU Q C, et al. The growth ofKandeliacandelseedlings in mangrove habitats of the Zhangjiang estuary in Fujian, China[J]. Acta Ecologica Sinica, 2006, 26(6): 1648-1655.
[3] 何东进, 游巍斌, 王 韧, 等. 中国红树林天然分布北缘区不同起源秋茄林湿地沉积物重金属空间分布特征比较[J]. 应用与环境生物学报, 2013, 19(6): 945-951.
[4] 何东进, 郑开基, 王 韧, 等. 闽东滨海湿地不同起源秋茄林重金属元素Zn、Cd、Cu的累积与分布特征比较[J]. 福建农林大学学报: 自然科学版, 2012, 41(2): 187-192.
[5] 何东进, 谭 勇, 廖小娟, 等. 闽东滨海湿地重金属生态风险评价方法选择与比较研究[J]. 福建林学院学报, 2014, 34(2): 97-103.
[6] 苏少川, 何东进, 王 韧, 等. 闽东滨海湿地生态系统服务功能价值评估[J]. 湿地科学与管理, 2012(3): 14-18.
[7] 吴桂觉. 福鼎滨海湿地及红树林调查研究初报[J]. 林业勘察设计, 2006(1): 142-144.
[8] 王 韧, 李晓景, 蔡金标, 等. 闽东沿海秋茄天然林与人工林生物量比较[J]. 西南林学院学报, 2010, 30(1): 16-20.
[9] 蒋雪琴, 刘艳红, 赵本元. 湖北神农架地区巴山冷杉(Abiesfargesii)种群结构特征与空间分布格局[J]. 生态学报, 2009, 29(5): 2211-2218.
[10] 何小娟, 何东进, 洪 伟, 等. 天宝岩天然猴头杜鹃种群空间分布格局研究[J]. 云南农业大学学报: 自然科学版, 2009, 24(5): 734-738.
[11] 李晓景, 何东进, 洪 伟, 等. 天宝岩自然保护区不同海拔天然柳杉种群空间分布格局研究[J]. 江西农业大学学报, 2009, 31(3): 422-426.
[12] 周建云, 李 荣, 张文辉, 等. 不同间伐强度下辽东栎种群结构特征与空间分布格局[J]. 林业科学, 2012, 48(4): 149-155.
[13] 范 娟, 赵秀海, 汪金松, 等. 江西九连山亚热带常绿阔叶林优势种空间分布格局[J]. 生态学报, 2012, 32(9): 2729-2737.
[14] 汤孟平, 周国模, 施拥军, 等. 天目山常绿阔叶林优势种群及其空间分布格局[J]. 植物生态学报, 2006, 30(5): 743-752.
[15] 何东进, 郑开基, 王 韧, 等. 闽东湿地重金属污染特征与生态风险评价[J]. 武夷科学, 2009, 25(12): 44-49.
[16] 王伯荪, 李鸣光, 彭少麟. 植物种群学[M]. 广州: 广东高等教育出版社, 1995.
[17] 赵学农, 刘伦辉. 版纳青梅种群结构动态与分布格局[J]. 植物学报, 1993, 35(7): 552-560.
[18] BRODIE C, HOULE G, FORTIN M-J. Development of aPopulusbalsamiferaclone in subarctic Québec reconstructed from spatial analyses[J]. Journal of Ecology, 1995, 83(2): 309-320.
[19] NUSKE R S, SPRAUER S, SABOROWSKI J. Adapting the pair-correlation function for analysing the spatial distribution of canopy gaps[J]. Forest Ecology and Management, 2009, 259(1): 107-116.
[20] OTTO R D. An evaluation of forest landscape spatial pattern and wildlife community structure[J]. Forest Ecology and Management, 1996, 89(1/3): 139-147.
[21] 巨天珍, 米彩燕, 师贺雄, 等. 小陇山国家级自然保护区油松种群更新及生存[J]. 山地学报, 2012, 30(1): 36-40.
[22] 方炎明, 张晓平, 王中生. 鹅掌楸生殖生态研究: 生殖分配与生活史对策[J]. 南京林业大学学报: 自然科学版, 2004, 28(3): 71-74.
[23] 昝启杰, 王勇军, 廖宝文, 等. 深圳福田无瓣海桑+海桑-秋茄人工林结构的研究[J]. 林业科学研究, 2001, 14(6): 610-615.
[24] 陈玉军, 廖宝文, 郑松发, 等. 无瓣海桑、海桑、秋茄红树人工林群落动态及物种多样性研究[J]. 应用生态学报, 2004, 15(6): 924-928.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
乡村科技(2021年1期)2021-12-03
山西林业(2021年2期)2021-07-21
园艺与种苗(2021年3期)2021-05-26
农村百事通(2020年21期)2020-11-23
农民致富之友(2020年4期)2020-04-03
绿色中国(2019年19期)2019-11-26
绿色中国(2019年19期)2019-11-26
中国水土保持科学(2019年6期)2019-04-26
