
亚硝酸盐对虾类毒性影响的研究进展
2014-06-26 09:54寇红岩冼健安苗玉涛叶超霞潘训彬王安利
海洋科学 2014年2期
寇红岩,冼健安,郭 慧,钱 坤,苗玉涛,叶超霞,潘训彬,王安利
(华南师范大学 生命科学学院,广东省水产健康安全养殖重点实验室,生态与环境科学广东普通高校重点实验室,广东 广州 510631)
1 水体亚硝酸盐的来源
亚硝酸盐是水产养殖系统中最常见的污染物之一。虾类主要以氨的形式进行氮废物的排泄,亚硝酸盐是由氨转变而来的,这过程由亚硝化细菌所完成。硝化细菌可进一步将亚硝酸盐转变为硝酸盐,可见亚硝酸盐是养殖水体中脱氮作用的中间产物,但由于硝化细菌的生长速度较亚硝化细菌慢很多,这一转变过程容易失衡,亚硝酸盐就被积累在养殖水体中[1]。因此随着养殖周期的延长,亚硝酸盐的浓度也会越来越高。在高密度对虾养殖池中,亚硝酸氮的质量浓度可达到20 mg/L[2]。
2 亚硝酸氮对虾类的半致死浓度
2.1 亚硝酸氮对不同虾种的半致死浓度
多种虾类的亚硝酸氮对不同虾种的半致死浓度(median lethal concentration,LC50)已有研究报道,结果见表1。对于同种虾类,不同学者的研究结果有一定的差异,可能是由于影响亚硝酸氮毒性的因素较多,不同研究条件有一定的差异所致。但总体上看,海水虾类比淡水虾类的亚硝酸氮耐受能力要高得多,主要与水体的盐度有关。
2.2 亚硝酸氮对不同虾龄的半致死浓度
斑节对虾(Penaeus monodon)[3]和长毛对虾(P.penicillatus)[4]的研究结果显示,各时期幼虫的亚硝酸盐耐受能力依次为: 仔虾>糠虾>溞状幼虫>无节幼虫。在凡纳滨对虾(Litopenaeus vannamei)的研究中,仔虾的 24h耐受力不如糠虾和溞状幼体,但仔虾的96 h耐受力则远高于糠虾和溞状幼体[5]。对于日本囊对虾(Marsupenaeus japonicus),溞状幼体的 24 h耐受力高于糠虾和仔虾,但96h时仔虾的耐受力最高[5]。刀额新对虾(Metapenaeus ensis)则表现为: 仔虾>糠虾>无节幼虫>溞状幼虫[6]。对于不同虾龄的成体,目前仍缺乏系统的比较研究,而不同学者研究的条件也存在一定的差异,较难进行准确的比较。
2.3 影响亚硝酸盐毒性的环境因素
目前研究结果显示,影响亚硝酸盐毒性的环境因素主要有盐度、Cl-含量、溶氧和 pH等。盐度对亚硝酸盐毒性的影响十分显著,盐度越高,虾类对亚硝酸盐的耐受能力就越强[7-9]。盐度影响其毒性可能主要与 Cl-含量有关,主要与 Cl-竞争鳃上的吸收位点,盐度越高,Cl-的含量就越高,竞争吸收进入体内的就越少。研究证实水环境中Cl-含量越高,虾类对亚硝酸盐的耐受能力也越强[10-12]。研究表明水体溶氧量越高,亚硝酸盐的毒性越低[13-14]。破坏虾类血蓝蛋白的正常携氧能力,导致机体缺氧是亚硝酸盐的毒性机制之一[15-18],因此水体溶氧量的提高可缓解亚硝酸盐的毒性作用。在水环境中,亚硝酸盐以亚硝酸根离子()和亚硝酸(HNO2)两种形式存在,后者能自由扩散通过鳃进入机体,毒性较大。在天然水体pH条件下,只有很少一部分亚硝酸盐以HNO2的形式存在,当水体pH下降时,以HNO2形式存在的亚硝酸盐的比例上升,因此在低pH条件下,亚硝酸盐的毒性较强。在pH为6.8、8.2和9.8的水体中,斑节对虾经0.72 mmol/L亚硝酸盐胁迫48 h后,血淋巴的亚硝酸盐浓度分别上升至3.70、2.87和1.91 µmol/L[19]。温度也是十分重要的环境因子,但目前仍未见关于温度影响亚硝酸盐对虾类的毒性的研究报道。
在实际养殖过程中,亚硝酸盐并不是单一存在的,氨氮、亚硝酸氮和硝酸氮可相互转化,各组分的比例会受到环境因子(溶氧、pH、盐度等)的影响,主要是由于这些环境因子改变了硝化或反硝化的方向和速率[20-22]。另外,养殖模式也会影响总氮和亚硝酸氮的含量[23]。因此,在池塘养殖中,亚硝酸盐毒性的影响因素和过程要复杂得多。
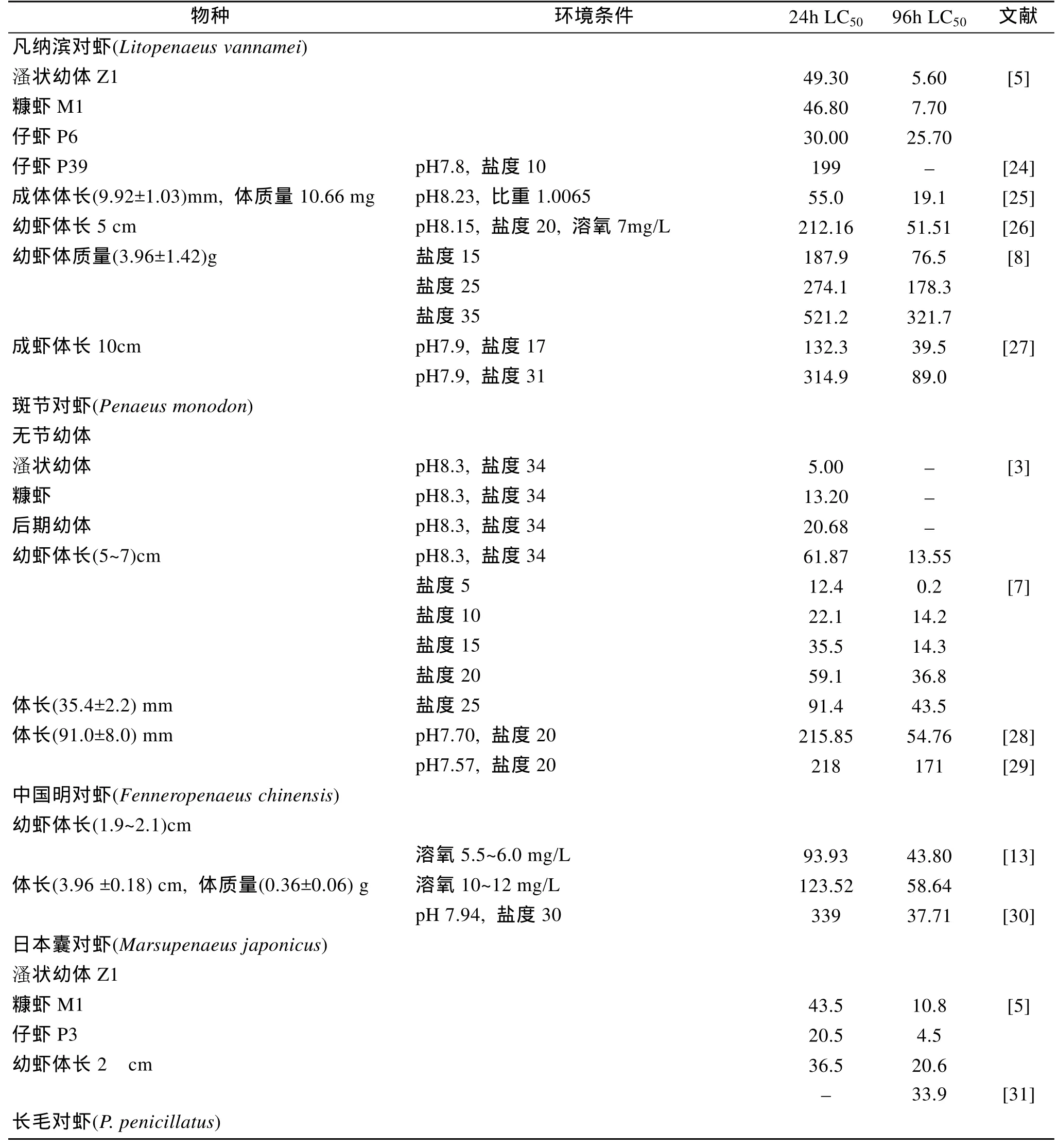
表1 亚硝酸氮对不同虾类的半致死浓度(LC50,mg/L)
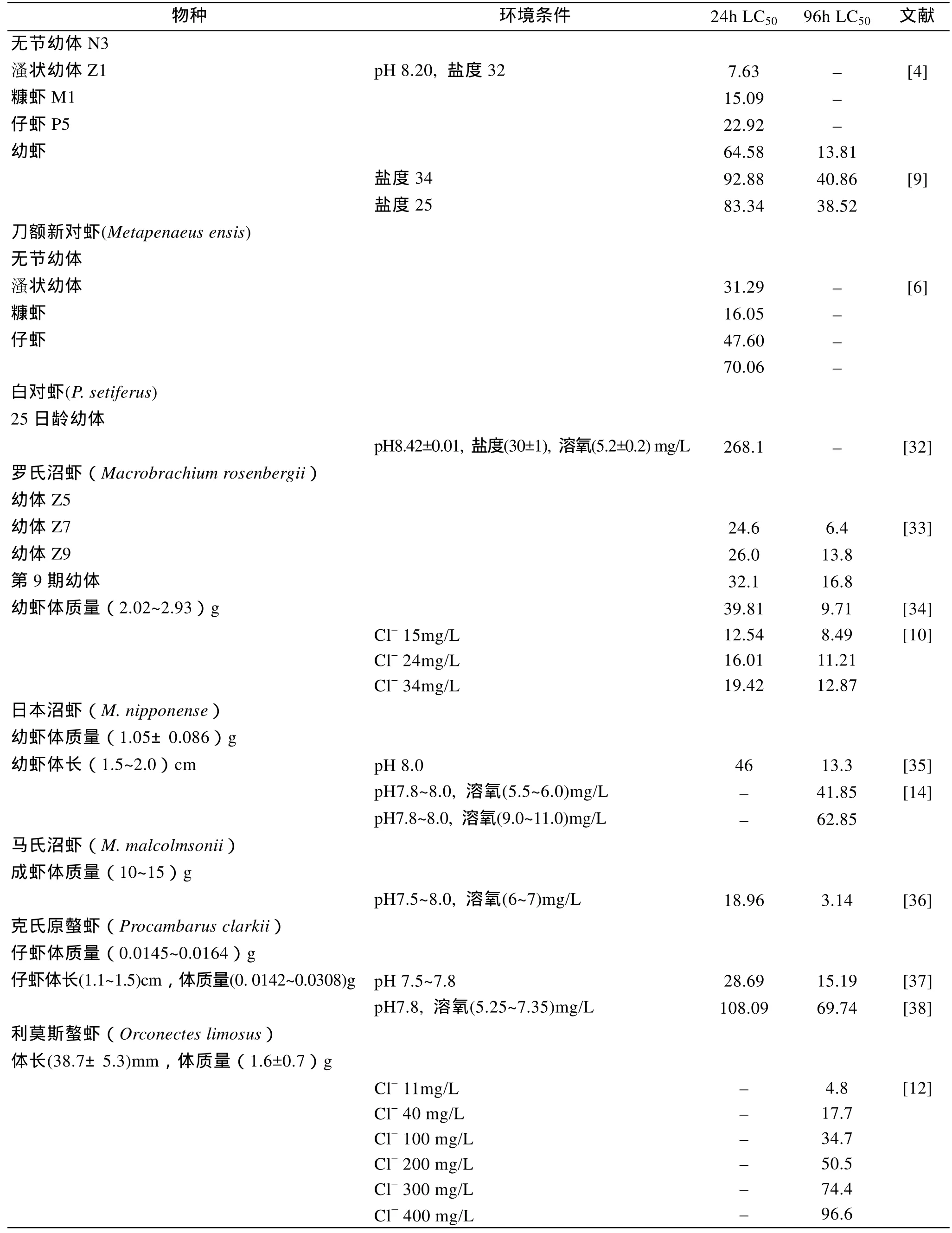
续表
3 亚硝酸盐的吸收和积累
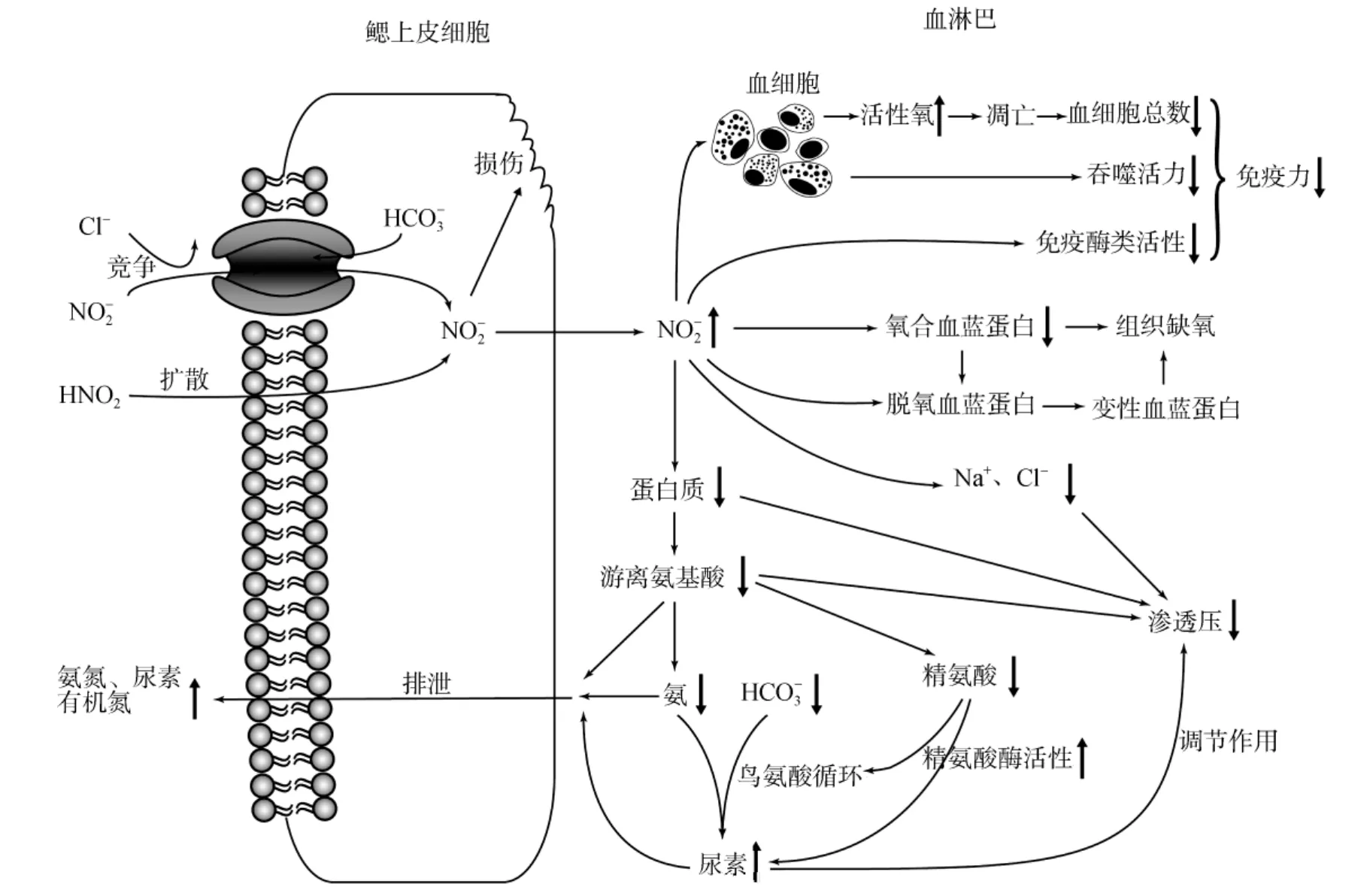
图1 亚硝酸盐对虾类血淋巴的影响
亚硝酸盐进入血淋巴后,被运送到其他组织,并有不同程度的积累。斑节对虾暴露于0.36 mmol/L的-N中48h后,肌肉、肝胰腺、鳃、前肠、血淋巴、心脏、眼柄和中肠的-N浓度分别上升至外环境浓度的 1.1、3.4、5.0、5.6、6.6、6.8、8.5和11.4倍,此研究结果显示肌肉基本不积累-N,而中肠积累的-N最多[42]。海水鱼类除通过鳃吸收亚硝酸盐外,还可以通过肠道进行吸收[1],斑节对虾的中肠是积累-N最多的组织,可能意味着海水虾类也有着相同的吸收机制,即在吞饮水的同时可能通过中肠吸收亚硝酸盐。
4 亚硝酸盐对虾类的毒性影响
4.1 对虾类生长和变态的影响
一定浓度的亚硝酸盐会抑制虾类的生长和变态发育。凡纳滨对虾置于(2.5~5.0)mg/L-N中养殖14 d,其体长、体质量增长率和存活率均低于对照组[25]。罗氏沼虾幼体经-N亚急性毒性作用,出现发育变态缓慢,增质量率和存活率下降等现象[33,43]。因此,控制水体中亚硝酸盐浓度对虾类健康苗种的培育至关重要的一环[44-45]。
4.2 对虾类组织结构的影响
血淋巴是虾体积累亚硝酸盐的主要部位之一,如前所示,血浆中的-N浓度可以累积达到外环境的6.6倍,因此在亚硝酸盐胁迫过程中,血细胞将遭受胞外高浓度-N的刺激。罗氏沼虾经-N应激后,血细胞总数(Total haemocyte count,THC)不断下降,血细胞的超微结构观察结果显示透明细胞出现核变小,常染色质、粗面内质网、核糖体和线粒体数量减少等现象,而半颗粒细胞和颗粒细胞除出现上述现象外,细胞质中的高电子致密颗粒数量也减少[46],这些结果表明-N导致了血细胞结构异常,这将进一步导致各种生理和免疫功能的异常。
4.3 对虾类血蓝蛋白及其载氧能力的影响
亚硝酸盐对血蓝蛋白载氧能力的破坏被认为是其对虾类产生毒性的主要机制之一。众多研究显示,虾类受到亚硝酸盐胁迫后,随着血淋巴亚硝酸盐浓度的上升,氧合血蓝蛋白和血清总蛋白含量、氧合血蓝蛋白占血清总蛋白含量的比例、血淋巴的氧亲和力均显著下降[15-18]。血蓝蛋白是甲壳动物机体内的载氧蛋白,亚硝酸盐进入血淋巴后,导致氧合血蓝蛋白转变为脱氧血蓝蛋白,并抑制脱氧血蓝蛋白与氧结合[15];另一方面,与亚硝酸盐破坏血红蛋白的机制类似,亚硝酸盐可能进一步导致脱氧血蓝蛋白氧化为变性血蓝蛋白,研究发现亚硝酸盐与脱氧血蓝蛋白的反应速率比其与氧合血蓝蛋白的反应速率快15倍[49]。因此,亚硝酸盐导致血淋巴的氧亲和力下降,破坏了其正常的携氧能力,最终导致虾类窒息死亡[15](图1)。
4.4 对虾类血淋巴生化组成和生理状态的影响
4.4.1 离子含量
正常状态下,鳃细胞中 Cl-的流入和的排出调节了Cl-转运和血淋巴的酸碱平衡。亚硝酸盐胁迫下,在 Cl-/交换过程中,由于的竞争,部分Cl-被替代,进入虾体的Cl-减少。小龙虾暴露于1 mmol/L亚硝酸盐1周后,血淋巴的Cl-含量下降了40%[41]。日本对虾暴露于0.36 mmol/L亚硝酸盐24 h后,血淋巴Cl-减少了58 mmol/L[50]。亚硝酸盐胁迫下,血淋巴中的也会下降[51],一方面可能由于与进行交换而排出体外,另一方面有研究者认为与氨通过鸟氨酸循环合成了尿素[52](图1)。
日本囊对虾的研究显示,亚硝酸盐胁迫对血淋巴中 K+、Ca2+和 Mg2+的含量没有显著影响,而 Na+含量则显著下降,作者认为 Na+含量的下降可能是虾体水分含量上升所致[50]。小龙虾的研究也发现亚硝酸盐胁迫导致血淋巴 Na+含量下降,另外还发现其肌肉的 K+含量大大下降,而在中肠腺则有升高的趋势,肌肉流失的部分 K+可能转移到了中肠腺内[53]。罗氏沼虾在低Cl-含量的水体中受到亚硝酸盐胁迫时,血淋巴的Na+、K+和Mg2+的含量均有显著下降[10]。
4.4.2 蛋白质和游离氨基酸含量
虾类血淋巴中含有大量的蛋白质,其中80%~95%为血蓝蛋白[15],剩下的部分主要是与免疫和凝集相关的因子,如酚氧化酶、凝集素、β-1,3-葡聚糖结合蛋白和抗菌肽等。血清总蛋白含量在亚硝酸盐胁迫下呈现显著的下降[15,17,50],其下降一方面可能由于部分蛋白质被加速代谢为氨氮;另一方面,在亚硝酸盐胁迫下,血清中多种免疫酶类的活性下降[27],血细胞产生各类功能蛋白的能力可能受到了抑制。与总蛋白含量的变化相同,血清游离氨基酸含量也呈现下降现象,另外还伴随着氨氮排泄量的显著上升[50](图 1),表明其下降可能是代谢速度加快所致。
4.4.3 氮代谢
在亚硝酸盐胁迫下,虾类的总氮、氨氮、尿素氮和有机氮的排泄量均显著提高[52,54]。日本对虾经0.076和1.433 mmol/L亚硝酸盐胁迫48 h后,其氨氮的排泄量分别是对照组对虾的1.7和2.9倍,尿素氮的排泄量分别是对照组对虾的263和694倍,有机氮的排泄量分别是对照组对虾的49和832倍[52]。这些结果表明,亚硝酸盐胁迫导致虾体的生理代谢过程加快。
研究表明,在十足目甲壳动物的氮排泄物中,氨氮占 60%~70%,氨基酸占 10%,另外还有少量的尿素、尿酸和氧化三甲胺[55]。亚硝酸胁迫下,氮代谢途径也发生了改变。日本对虾在正常生理状态下,氨氮和尿素氮分别占排泄总氮的 41.7%~90.8%和2.8%~10.5%;暴露于0.076和1.433 mmol/L亚硝酸盐 24 h后,氨氮占排泄总氮的比例分别下降至10.0%和3.8%,其尿素氮的比例分别上升至58.7%和21.4%,而有机氮的比例则分别上升至 31.3%和74.8%[52]。
尿素可来源于3种途径: (1)鸟氨酸循环;(2)尿酸分解;(3)精氨酸水解[52]。日本对虾经亚硝酸盐胁迫后,血淋巴中的氨氮、和精氨酸含量均下降,而尿素含量上升[52]。日本沼虾经1.39 mmol/L亚硝酸盐暴露 24 h后,肝胰腺和血淋巴中的精氨酸酶活性显著升高[54],这些结果表明尿素含量的升高可能来源于鸟氨酸循环(氨与的合成)或者精氨酸水解[50,52,54]。尿素合成的加强一方面可能为了降低血氨的含量,从而降低血氨与外界摄入的亚硝酸盐的联合毒性[54];另一方面尿素可能起着调节渗透压的作用[50,56](图 1)。
4.4.4 渗透压和pH
虾血清渗透压随着亚硝酸盐胁迫时间的延长呈现逐渐下降的现象,且胁迫浓度越大,下降速度越快。日本囊对虾经0.72 mmol/L亚硝酸盐胁迫48 h后,渗透压下降至 884.5 mOsm/kg,为对照组的96.8%[50]。如前所述,在亚硝酸盐的作用下,虾血淋巴中的 Cl-、Na+、蛋白质和游离氨基酸等含量均下降,是导致血清渗透压下降的主要原因;另外,虾体水分含量的升高也是造成渗透压下降的原因之一[50](图1)。多种虾类的研究均发现亚硝酸盐会导致血淋巴pH下降[10,16,51,57],含量的下降可能主要原因之一。
4.5 对虾类免疫功能的影响
凡纳滨对虾置于含有亚硝酸氮 4.0和 8.0 mg/L的水体中养殖 18 d后,其 THC、超氧化物歧化酶(SOD)活力、PO活力、抗菌活力、溶菌活力和血清蛋白含量均显著下降,亚硝酸氮浓度越高,对虾的免疫功能越低[27]。红螯螯虾置于(0.3~6.0)mg/L亚硝酸盐下饲养2周后,螯虾肝胰腺、鳃和肌肉中的酸性磷酸酶(ACP)、碱性磷酸酶(AKP)、SOD和谷胱甘肽过氧化物酶(GPx)活性均显著下降,鳃组织 Na+,K+-ATPase和Ca2+,Mg2+-ATPase的活性也呈现下降的趋势[48]。将凡纳滨对虾暴露于5.15,11.06和21.40 mg/L的亚硝酸氮中 48~168 h,会导致弧菌(Vibrio alginolyticus)注射后的对虾的死亡率上升;对虾暴露于4.94,9.87 和19.99 mg/L的亚硝酸氮中,其THC和PO显著下降。在马氏沼虾[36]和罗氏沼虾[58]的研究中也得到类似的结果。这些结果表明亚硝酸盐胁迫抑制了对虾的免疫功能,增加了其对病原体的易感性。
凡纳滨对虾在9.87 和19.99 mg/L的亚硝酸氮暴露96 h后,血细胞的超氧阴离子()含量显著上升,这些升高的可能对机体产生氧化毒性[2]。日本沼虾暴露于亚致死浓度的亚硝酸氮水体中24 h后,活性氧(ROS)含量增加 97%,而 SOD、过氧化氢酶(CAT)和GPx活力则分别下降了67%、80.6%和82.7%。ROS含量过高会导致机体受到氧化损伤[59],亚硝酸盐胁迫使 ROS的产生和清除失衡,从而导致组织受到氧化损伤可能是其毒性机制之一[35]。这一氧化毒性机制在斑节对虾血细胞的研究中得到了证实,将斑节对虾置于 20 mg/L的亚硝酸氮水体中应激 48 h后,其THC显著下降,同时ROS含量显著提高,DNA损伤细胞和凋亡细胞的比例均显著上升,表明亚硝酸盐诱导了血细胞产生过量的ROS,这些ROS对血细胞造成了氧化伤害并进一步诱导了细胞凋亡,导致了THC的下降,而THC下降到一定的阀值将会导致虾的死亡[60](图1)。
5 展望
上述研究结果显示,虾类的存活、生长发育、生理代谢和免疫功能等均会受到水体亚硝酸盐的毒性影响。但是,这些研究仍不够深入,其毒性影响的作用机制仍不明确。今后可从以下几个方面开展进一步的研究:
(1)温度是近年来十分受关注的环境因子,研究温度对亚硝酸盐毒性的影响及其机制对虾类的健康养殖有着重要指导意义;(2)近年来的研究认为虾类存在类免疫球蛋白[61],可进一步开展亚硝酸盐对这些类免疫球蛋白影响的研究;(3)利用最新的组学技术,研究亚硝酸盐对虾类毒性影响的分子机制。
这些研究也表明,亚硝酸盐是虾类健康养殖过程中不可忽视的环境因子,必须进行严格的监控。但是,亚硝酸盐的控制是多方面的,其中健康合理的营养投喂是控制亚硝酸盐的重要因素之一。营养搭配合理的配合饲料不但可以减少虾类氮废物的排放,而且可以提高虾类的免疫功能和抗胁迫能力。但目前针对虾类抗亚硝酸盐胁迫能力的研究仍甚少[62,63],未来的研究可更加关注营养物质和各类添加剂[64]在虾类抗亚硝酸盐胁迫能力方面的作用。另外,通过现代遗传育种手段选育抗性强的品系也是提高养殖虾类抗亚硝酸盐胁迫能力的重要途径之一[65]。
[1]高明辉,马立保,葛立安,等.亚硝酸盐在水生动物体内的吸收机制及蓄积的影响因素[J].南方水产,2008,4(4): 73-79.
[2]Tseng I.The immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus under nitrite stress[J].Fish &Shellfish Immunology,2004,17(4): 325-333.
[3]周光正.氨和亚硝酸盐对于对虾幼虫的毒性[J].海洋湖沼通报,1991,2: 95-98.
[4]高淑英,邹栋梁.亚硝酸盐对长毛对虾幼体的毒性[J].台湾海峡,1994,13(3): 236-239.
[5]姚庆祯,徐桂荣.亚硝酸盐和氨对凡纳对虾和日本对虾幼体的毒性作用[J].上海水产大学学报,2002,11(1): 21-26.
[6]Chen J C,Nan F H.Lethal effect of nitrite on Metapenaeus ensis larvae[J].Journal of the World Aquaculture Society,1991,22(1): 51-56.
[7]胡贤德,孙成波,王平,等.不同盐度条件下亚硝酸氮对斑节对虾的毒性影响[J].沈阳农业大学学报,2009,40(1): 103-106.
[8]Lin Y C,Chen J C.Acute toxicity of nitrite on Litopenaeus vannamei (Boone)juveniles at different salinity levels[J].Aquaculture,2003,224(1-4): 193-201.
[9]Chen J C,Lin C Y.Lethal effects of ammonia and nitrite on Penaeus penicillatus juveniles at two salinity levels[J].Comparative Biochemistry and Physiology C,1991,100(3): 477-482.
[10]Chen J C,Lee Y.Effects of nitrite on mortality,ion regulation and acid-base balance of Macrobrachium rosenbergii at different external chloride concentrations[J].Aquatic Toxicology,1997,39(3-4): 291-305.
[11]Yildiz H Y,Benli A.Nitrite toxicity to crayfish,Astacus leptodactylus,the effects of sublethal nitrite exposure on hemolymph nitrite,total hemocyte counts,and hemolymph glucose[J].Ecotoxicology and Environmental Safety,2004,59(3): 370-375.
[12]Kozak P,Machova J,Policar T.The effect of chloride content in water on the toxicity of sodium nitrite for spiny-cheek crayfish (Orconectes limosus RAF.)[J].Bull Fr Pêche Piscic,2005,376-377: 705-714.
[13]王娟,曲克明,刘海英,等.不同溶氧条件下亚硝酸盐和氨氮对中国对虾的急性毒性效应[J].海洋水产研究,2007,28(6): 1-6.
[14]朱大世,罗静波,吴宇略.不同溶氧条件下 NO2--N和 NH3-N 对青虾的急性毒性效应[J].长江大学学报(农学卷),2010(3): 40-42.
[15]Chen J C,Cheng S Y.Changes of oxyhemocyanin and protein levels in the hemolymph of Penaeus japonicus exposed to ambient nitrite[J].Aquatic Toxicology,1995,33(3-4): 215-226.
[16]Chen J,Cheng S.Hemolymph oxygen content,oxyhemocyanin,protein levels and ammonia excretion in the shrimp Penaeus monodon exposed to ambient nitrite[J].Journal of Comparative Physiology B,1995,164(7): 530-535.
[17]Cheng S Y,Chen J C.Hemocyanin oxygen affinity,and the fractionation of oxyhemocyanin and deoxyhemocyanin for Penaeus monodon exposed to elevated nitrite[J].Aquatic Toxicology,1999,45(1): 35-46.
[18]Cheng S Y,Chen J C.Study on the oxyhemocyanin,deoxyhemocyanin,oxygen affinity and acid-base balance of Marsupenaeus japonicus following exposure to combined elevated nitrite and nitrate[J].Aquatic Toxicology,2002,61(3-4): 181-193.
[19]Chen J C,Cheng S Y.Recovery of Penaeus monodon from functional anaemia after exposure to sublethal concentration of nitrite at different pH levels[J].Aquatic Toxicology,2000,50(1-2): 73-83.
[20]王伟良,李德尚,董双林,等.养虾围隔中无机氮浓度与放养密度及环境因子的关系[J].海洋科学,2000,24(10): 44-47.
[21]孙舰军.虾池生态系统中诸因子对虾体的影响[J].海洋科学,1997,21(2): 24-25.
[22]查广才,麦雄伟,周昌清,等.凡纳滨对虾淡化高产虾池水体生态特征[J].海洋科学,2006,30(9): 58-62.
[23]夏新建,许忠能,林小涛,等.不同池塘养殖模式的环境氮磷负荷及其水质特征[J].海洋科学,2012,36(5): 87-92.
[24]Schuler D J,Boardman G D,Kuhn D D,et al.Acute toxicity of ammonia and nitrite to Pacific white shrimp,Litopenaeus vannamei,at low salinities[J].Journal of the World Aquaculture Society,2010,41(3): 438-446.
[25]彭自然,臧维玲,高杨,等.氨和亚硝酸盐对凡纳滨对虾幼虾的毒性影响[J].上海水产大学学报,2004,13(3): 274-278.
[26]孙国铭,汤建华.氨氮和亚硝酸氮对南美白对虾的毒性研究[J].水产养殖,2002,1: 22-24.
[27]黄翔鹄,李长玲,郑莲,等.亚硝酸盐氮对凡纳滨对虾毒性和抗病相关因子影响[J].水生生物学报,2006,30(4): 466-471.
[28]Chen J C,Lei S C.Toxicity of ammonia and nitrite to Penueus monodon juveniles[J].Journal of the World Aquaculture Society,1990,21(4): 300-306.
[29]Chen J C,Liu P C,Lei S C.Toxicities of ammonia and nitrite to Penaeus monodon adolescents[J].Aquaculture,1990,89(2): 127-137.
[30]Chen J C,Ting Y Y,Lin J N,et al.Lethal effects of ammonia and nitrite on Penaeus chinensis juveniles[J].Marine Biology,1990,107(3): 427-431.
[31]曾红,林能锋,张文焕,等.亚硝酸氮对日本对虾(Penaeus japonicus)幼虾的急性毒性试验[J].福建畜牧兽医,2006,28(5): 13-14.
[32]Alcaraz G,Chiappa-Carrara X,Espinoza V,et al.Acute toxicity of ammonia and nitrite to white shrimp Penaeus setiferus postlarvae[J].Journal of the World Aquaculture Society,1999,30(1): 90-97.
[33]臧维玲,沈林华.亚硝酸盐和氨对罗氏沼虾幼体的毒性[J].上海水产大学学报,1996,5(1): 15-22.
[34]刘淑梅,吴维宁.亚硝酸盐氮对罗氏沼是幼体的毒性试验[J].水产科技情报,1999,26(6): 281-283.
[35]Wang W N,Wang A L,Zhang Y J,et al.Effects of nitrite on lethal and immune response of Macrobrachium nipponense[J].Aquaculture,2004,232(1-4): 679-686.
[36]Chand R K,Sahoo P K.Effect of nitrite on the immune response of freshwater prawn Macrobrachium malcolmsonii and its susceptibility to Aeromonas hydrophila[J].Aquaculture,2006,258(1-4): 150-156.
[37]罗静波,曹志华,温小波,等.亚硝酸盐氮对克氏原螫虾仔虾的急性毒性效应[J].长江大学学报(自然科学版),2005,2(11): 64-66.
[38]於叶兵,陆伟,黄金田,等.亚硝酸盐和硫化物对克氏原螯虾幼虾的毒性效应研究[J].水生态学杂志,2011,32(1): 111-114.
[39]Chen J C,Chen S F.Accumulation of nitrite in hemolymph of Penaeus japonicus[J].Marine Ecology Progress Series,1992,83(2): 305-308.
[40]Chen J C,Chen S F.Accumulation of nitrite in the haemolymph of Penaeus monodon exposed to ambient nitrite[J].Comparative Biochemistry and Physiology C,1992,103(3): 477-481.
[41]Jensen F B.Uptake,elimination and effects of nitrite and nitrate in freshwater crayfish (Astacus astacus)[J].Aquatic Toxicology,1996,34(2): 95-104.
[42]Cheng S Y,Chen J C.Accumulation of nitrite in the tissues of Penaeus monodon exposed to elevated ambient nitrite after different time periods[J].Archives of Environmental Contamination and Toxicology,2000,39(2): 183-192.
[43]Mallasen M,Valenti W C.Effect of nitrite on larval development of giant river prawn Macrobrachium rosenbergii[J].Aquaculture,2006,261(4): 1292-1298.
[44]林更铭,杨清良,许章程.对虾无公害生态育苗[J].海洋科学,2007,31(10): 11-14.
[45]徐琴,李健,刘淇,等.4种微生态制剂对对虾育苗水体主要水质指标的影响[J].海洋科学,2009,33(3):10-15.
[46]胡义波,王玥,姜乃澄.氨态氮、亚硝态氮对罗氏沼虾血细胞及超微结构的影响[J].浙江大学学报(理学版),2005,32(6): 691-697.
[47]吴中华,刘昌彬.中国对虾慢性亚硝酸盐和氨中毒的组织病理学研究[J].华中师范大学学报(自然科学版),1999,33(1): 119-122.
[48]吕晓燕,李嘉尧,方燕,等.亚硝酸盐对红螯光壳螯虾不同组织免疫相关酶活性及超微结构的影响[J].水产学报,2010,34(12): 1812-1820.
[49]Tahon J P,Van H D,Vinckier C,et al.The reaction of nitrite with the haemocyanin of Astacus leptodactylus[J].Biochem J,1988,249(3): 891-896.
[50]Cheng S Y,Chen J C.Effects of nitrite exposure on the hemolymph electrolyte,respiratory protein and free amino acid levels and water content of Penaeus japonicus[J].Aquatic Toxicology,1998,44(1-2):129-139.
[51]Chen J C,Cheng S Y.Hemolymph osmolality,acid-base balance,and ammonia excretion of Penaeus japonicus bate exposed to ambient nitrite[J].Archives of Environmental Contamination and Toxicology,1996,30(2): 151-155.
[52]Cheng S Y,Chen J C.The time-course change of nitrogenous excretion in the Kuruma shrimp Penaeus japonicus following nitrite exposure[J].Aquatic Toxicology,2001,51(4): 443-454.
[53]Jeberg M V,Jensen F B.Extracellular and intracellular ionic changes in crayfish Astacus astacus exposed to nitrite at two acclimation temperatures[J].Aquatic Toxicology,1994,29(1-2): 65-72.
[54]Lee W C,Chen J C.Nitrogenous excretion and arginase specific activity of kuruma shrimp Marsupenaeus japonicus exposed to elevated ambient nitrite[J].Journal of Experimental Marine Biology and Ecology,2004,308(1): 103-111.
[55]Regnault M.Nitrogen excretion in marine and freshwater crustacea[J].Biol Rev,1987,62: 1-24.
[56]Chen J C,Cheng S Y.Accumulation of urea in the haemolymph and ammonia excretion of Penaeus japonicus exposed to ambient nitrite[J].Comparative Biochemistry and Physiology C,1995,110(1): 1-6.
[57]Jensen F B.Sublethal physiological changes in freshwater crayfish,Astacus astacus,exposed to nitrite:haemolymph and muscle tissue electrolyte status,and haemolymph acid-base balance and gas transport[J].Aquatic Toxicology,1990,18(1): 51-60.
[58]Cheng W,Liu C H,Chen J C.Effect of nitrite on interaction between the giant freshwater prawn Macrobrachium rosenbergii and its pathogen Lactococcus garvieae[J].Diseases of Aquatic Organisms,2002,50(3): 189-197.
[59]冼健安,苟妮娜,陈晓丹,等.流式细胞术检测虾类血细胞活性氧含量方法的建立 [J].海洋科学,2012,36(2): 29-33.
[60]Xian J A,Wang A L,Chen X D,et al.Cytotoxicity of nitrite on haemocytes of the tiger shrimp,Penaeus monodon,using flow cytometric analysis[J].Aquaculture,2011,317: 240-244.
[61]章跃陵,刘文杰,王三英,等.凡纳滨对虾类 Ig定性和定位的初步研究[J].海洋科学,2005,29(9): 31-35.
[62]Chand R K,Sahoo P K,Kumari J,et al.Dietary administration of bovine lactoferrin influences the immune ability of the giant freshwater prawn Macrobrachium rosenbergii (de Man)and its resistance against Aeromonas hydrophila infection and nitrite stress[J].Fish Shellfish Immunol,2006,21(2): 119-129.
[63]杨丽坤,蔡端波,黄增瑞,等.硒对日本沼虾耐受亚硝酸钠胁迫的作用[J].安徽农业科学,2010,38(34):19547.
[64]张海波,吴嘉敏,谭洪新,等.溶菌酶和抗菌肽在对虾养殖中的应用[J].海洋科学,2008,32(6): 61-65.
[65]张灵侠,沈琪,胡超群.凡纳滨对虾的遗传育种研究现状[J].海洋科学,2008,32(2): 91-95.
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年8期)2022-09-20
当代水产(2022年5期)2022-06-05
当代水产(2021年8期)2021-11-04
学苑创造·A版(2021年10期)2021-10-30
当代水产(2021年4期)2021-07-20
农民致富之友(2020年27期)2020-09-12
湖北大学学报(自然科学版)(2020年3期)2020-06-18
饲料工业(2019年24期)2019-12-31
物理化学学报(2017年8期)2017-12-18
