
黑尾叶蝉精氨酸激酶和一个谷胱甘肽-S-转移酶基因的克隆和发育表达特性研究
2014-08-10 12:29侯吉祥孟盼盼马伟华林拥军
植物保护 2014年1期
侯吉祥,孟盼盼,马伟华,周 菲,林拥军
(华中农业大学作物遗传改良国家重点实验室, 武汉 430070)
黑尾叶蝉精氨酸激酶和一个谷胱甘肽-S-转移酶基因的克隆和发育表达特性研究
侯吉祥,孟盼盼,马伟华,周 菲,林拥军*
(华中农业大学作物遗传改良国家重点实验室, 武汉 430070)
黑尾叶蝉(Nephotettixcincticeps)是一种能传播病毒的水稻害虫,对水稻生产有着巨大的威胁。本文在黑尾叶蝉中克隆得到了一个精氨酸激酶基因(NcAK)和一个delta类型的谷胱甘肽-S-转移酶基因(NcGST),并对两个基因进行了序列分析和表达特性检测。结果表明,NcAK包含的开放阅读框长度为1 068 bp,编码一个含356个氨基酸的蛋白。进化研究表明该蛋白序列与玻璃翅叶蝉(Homalodiscavitripennis)的精氨酸激酶的序列亲缘关系最近。Real-time结果表明NcAK在黑尾叶蝉的雄成虫中表达量高于在卵、1龄若虫、5龄若虫和雌成虫中的表达量。NcGST的开放阅读框由651个碱基组成,编码一个包含217氨基酸的蛋白,进化研究表明该蛋白与美国牧草盲蝽(Lyguslineolaris)GSTs编码蛋白亲缘关系最近。表达量检测表明NcGST在5龄若虫和雌雄成虫中的表达量要高于在卵和1龄的表达量。
黑尾叶蝉; 精氨酸激酶; 谷胱甘肽-S-转移酶
黑尾叶蝉[Nephotettixcincticeps(Uhler)]是一种主要分布在长江中上游和西南各省的水稻害虫,它不仅通过吸取水稻汁液为害水稻,同时能够传播水稻黄矮病、水稻矮缩病等水稻病毒病,一旦病害暴发将对水稻产量造成极大的影响[1]。目前对黑尾叶蝉主要采用农药防治。但是由于农药的过量使用,黑尾叶蝉已经对有机磷类和有机氯类农药产生了一定的抗药性,需要寻找发展更安全高效的抗虫方法。
精氨酸激酶是仅存于无脊椎动物体内的磷酸原激酶,主要功能是通过催化精氨酸和ATP之间的可逆反应将能量存储于磷酸精氨酸的高能磷酸键中,磷酸精氨酸是昆虫肌肉中唯一有效形成ATP的磷酞基供体[2]。不仅如此,精氨酸激酶在甲壳动物的免疫中也起着一定的作用[3]。目前已经在多个物种中都得到了精氨酸激酶的cDNA序列[4-5]。
谷胱甘肽-S-转移酶Glutathione-S-transferase (GSTs)是多个基因编码、具有多种功能的超基因家族酶,分子量23~29 ku。根据亚细胞定位,谷胱甘肽-S-转移酶可以分为微粒体型(microsomal GSTs)、线粒体型(mitochondrial GSTs)和胞质型(cytosolic GSTs)三大类型。其中胞质谷胱甘肽-S-转移酶是研究最多的类型,通常所指的谷胱甘肽-S-转移酶即为此类。昆虫胞质谷胱甘肽-S-转移酶包括omega、sigma、theta、zeta、delta和epsilon 6个已知的家族[6]。其中delta家族和epsilon家族是昆虫生物体特有的家族。谷胱甘肽-S-转移酶对昆虫的对农药的敏感性降低有着重要的意义。研究表明,谷胱甘肽-S-转移酶在有机磷杀虫剂尤其是二甲基膦酸酯和二甲基硫代膦酸酯的代谢中起着相当重要的作用,对保棉磷、碘甲烷、3,4-二氯硝基苯也都表现出很高的代谢活性[7]。在对有机磷农药有抗性的德国小蠊(BlattellagermanicaLinnaeus)的研究中,吴刚等证明抗性品系德国小蠊的谷胱甘肽-S-转移酶酶活力相对于敏感品系明显提高[8]。Zhu等发现GSTs抑制剂能够降低美国牧草盲蝽对农药原料1-氯-2,4-二硝基苯和马拉硫磷的抗性[9],提高农药对盲蝽的致死率。目前已经有研究表明黑尾叶蝉对敌敌畏等有机磷农药具有很高的抗药性[10],而GSTs对于有机磷类和有机氯农药在昆虫体内的代谢中起着很重要的作用,因此研究黑尾叶蝉的GSTs对研究黑尾叶蝉的解毒能力有着重要的意义。本文克隆了一个编码黑尾叶蝉的精氨酸激酶的NcAK基因和一个编码黑尾叶蝉谷胱甘肽转移酶的NcGST基因的CDS序列,对序列进行了保守结构域预测和系统进化树分析, 并检测它们mRNA的表达特性,为以后深入研究精氨酸激酶和谷胱甘肽转移酶在黑尾叶蝉体内的表达调控及谷胱甘肽转移酶的解毒机制打下了基础。
1 材料与方法
1.1 供试昆虫
黑尾叶蝉于2009年8月采集于江西南昌,然后于本实验室内用水稻感虫品种‘TN1’保存。培养条件为:温度(26±2)℃,相对湿度50%,光照周期L∥D=14 h∥10 h。
1.2 材料
限制性内切酶、Taq酶、TA克隆试剂盒、Trizol试剂、DEPC为上海生工产品;反转录试剂盒购自Roche公司;特异引物由武汉安基生物有限公司合成。
1.3 方法
1.3.1 总RNA的提取和第一链cDNA的合成
取黑尾叶蝉在液氮中研磨成粉末,用Trizol 试剂盒提取总RNA,用DNA酶消化痕量DNA,按反转录试剂盒说明合成第一链cDNA。
1.3.2 基因的扩增
设计NcAK和NcGST的扩增引物见表1,以黑尾叶蝉的cDNA为模板,进行扩增,扩增条件为:95 ℃ 5 min;94 ℃ 30 s,58 ℃30 s,72 ℃ 1 min,32 个循环;终延伸72 ℃ 7 min。
表1NcAK和NcGST的扩增引物序列
Table1TheprimersofNcAKandNcGST

引物名称Primername引物序列Primersequence(5′→3′)NcAKFCTCCTAGCGTCACTCCGTTCNcAKRTGAAAATAGGAGCCGCCGTGNcGSTFCTGATGCGTAGTGAGGTGTGANcGSTRTGCAGTCACTTCGGTTGTGT
1.3.3 基因的克隆与测序
取4μL 的PCR产物,用p-EASY T3 clone kit进行TA克隆;从蓝白斑平板中挑取单个白色菌落扩大培养,重组质粒用EcoR I 酶切鉴定,选择其中4个阳性克隆,用引物SP6/T7测序。
1.3.4 序列分析、同源性分析和进化树的构建
利用DNAman软件对两个基因进行翻译,利用软件DNAman对蛋白质等电点和分子量进行预测。利用NCBI在线工具Conserved Domains Search(www.ncbi.nlm.gov/structure/cdd/wrpsb.cgi)对两个基因进行保守结构域的搜索。利用NCBI在线工具Blastx对两个基因进行同源基因的寻找。采用Mega5.0软件的邻接法(NJneighbor-joining)构建进化距离分析的P-distance模型,系统树每个分支的统计学显著性分析以bootstrap 进行检验,重复次数为1 000 次。
1.4 NcAK和NcGST的表达特性研究
分别提取不同发育阶段和性别黑尾叶蝉(24 h内产下的虫卵、24 h内蜕皮的1龄若虫、5龄若虫和雌雄成虫)的总RNA,取2 μg 总RNA 反转录成cDNA 模板,再设计引物(表2),检测基因表达量。使用TaKaRa公司的real-time试剂盒20 μL体系。扩增条件:95 ℃预变性30 s;然后95 ℃ 35 s,60 ℃36 s。共40个循环;利用Excel软件使用 2-△△Ct法将获得的数据进行方差分析,利用SPSS软件中Duncan 氏新复极差法进行差异显著性分析。
表2NcAK和NcGST的RT引物序列
Table2TheprimersforRT-PCRofNcAKandNcGST
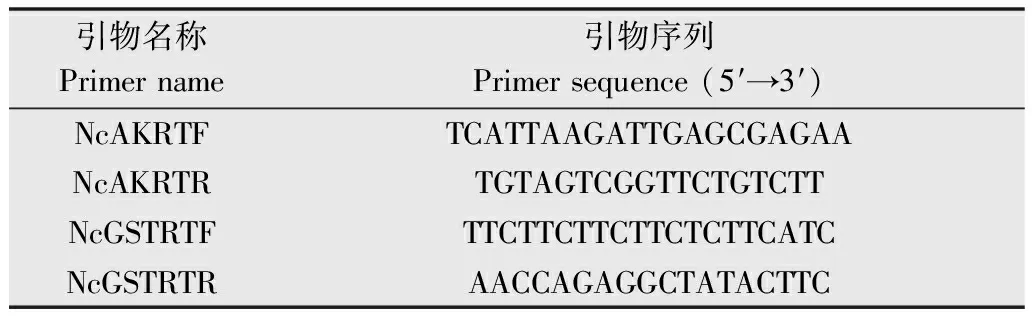
引物名称Primername引物序列Primersequence(5′→3′)NcAKRTFTCATTAAGATTGAGCGAGAANcAKRTRTGTAGTCGGTTCTGTCTTNcGSTRTFTTCTTCTTCTTCTCTTCATCNcGSTRTRAACCAGAGGCTATACTTC
2 结果与分析
2.1 NcAK和NcGST序列分析
扩增得到两条长度分别为799 bp和1 144 bp的cDNA片段,NcAK基因的开放阅读框由1 068个碱基组成,编码356个氨基酸,分子量为 40 134.59,推测蛋白序列等电点为5.98。NcGST的开放阅读框由651个碱基组成,编码217个氨基酸,推测蛋白序列分子量为24 168.7,等电点为6.74。其核酸序列及按三联密码子推导出的氨基酸序列见图1和图2。用NCBI在线工具 Conserved Domains Search(www.ncbi.nlm.gov/structure/cdd/wrpsb.cgi)预测发现NcAK具有精氨酸激酶的保守结构域,NcGST具有delta和epsilon类型的GSTs保守结构域。BlastX比对分析显示,NcAK基因推测编码蛋白与其他物种的精氨酸激酶蛋白序列相似性高达90%~97%,而NcGST基因推测编码蛋白与其他物种GSTs基因蛋白序列相似性达到60%~74%,其中与delta类型的GSTs基因相似程度最高。因此推测,NcAK基因编码精氨酸激酶而NcGST基因编码一个delta类型的谷胱甘肽转移酶。

图1 NcAK的开放阅读框推测得到的蛋白序列Fig.1 The ORF and the deduced protein of NcAK

图2 黑尾叶蝉谷胱甘肽-S-转移酶NcGST的开放阅读框和推测得到蛋白序列Fig.2 The ORF and the deduced protein of NcGST
2.2 序列的同源性分析和进化树构建
为了解NcAK和NcGST与其他物种精氨酸激酶和GSTs的亲缘关系,利用Mega5.0软件NcAK和NcGST预测的蛋白序列与其他物种同源蛋白序列进行进化分析。结果(图3和图4)显示,在选取的昆虫物种中NcAK与玻璃翅叶蝉(Homalodiscavitripennis)的精氨酸激酶和叶蝉(OncometopianigricansWalker)的精氨酸激酶同缘关系最近。而NcGST与半翅目的美国牧草盲蝽(LyguslineolarisGoeze)、半翅目的白背飞虱(SogatellafurciferaHorváth)和褐飞虱(NilaparvatalugensStål)的GSTs亲缘关系最近。
2.3 基因表达特性研究
以核蛋白L19为内参,检测了NcAK和NcGST在黑尾叶蝉24 h内产的卵,24 h内蜕皮产生的1龄和5龄若虫以及雌雄成虫中的表达情况,结果发现NcAK基因在黑尾叶蝉的几个龄期中都有表达,在雄成虫的表达量最高,差异性显著(P<0.05),在其他龄期表达量差异不大。而NcGST在5龄若虫和雌雄成虫中的表达量要高于在卵中和1龄若虫中的表达量。
3 讨论
本研究克隆得到了黑尾叶蝉精氨酸激酶的CDS序列,并进行了序列分析和表达特性研究。经过序列分析和同源性比对发现,NcAK推测得到的蛋白序列和其他昆虫中得到的精氨酸激酶蛋白序列氨基酸一致性高达80% ~ 97%。这证明克隆得到的确实是精氨酸激酶,氨基酸相似性高度一致说明精氨酸激酶在进化中是高度保守的。这也证明了该基因的重要性。表达量检测表明,该基因在黑尾叶蝉的不同龄期都有表达,在雄成虫中表达量最高,和其余龄期差异显著(P<0.05),这也许与雄成虫的发育特性有关。在其余的龄期中表达量差异不显著。这表明NcAK的表达在成虫中表现出了性别差异而没有表现出不同龄期之间的差异。部分昆虫如家蚕存在精氨酸表达量随龄期的增加而增加的现象[4],这也许与家蚕是完全变态昆虫而黑尾叶蝉不是完全变态昆虫有关。
人们已经以精氨酸激酶为靶标进行了大量抗虫研究,徐秀凤等利用饲喂以精氨酸激酶为靶标的dsRNA的方式提高了小菜蛾(PlutellaxylostellaLinnaeus)的死亡率[11]。苏晓峰等发现利用RNAi的方法沉默精氨酸激酶基因可以有效延缓棉铃虫的生长发育[12]。而在黄曲条跳甲(PhyllotretastriolataFabricius)精氨酸激酶的研究中,Zhao通过RNAi方式沉默精氨酸激酶的表达,结果提高了黄曲条跳甲的死亡率,并降低了雌性黄曲条跳甲的产卵量[13]。这些试验证明,以精氨酸激酶为靶标进行抗虫是一种可行的方式。在黑尾叶蝉的抗虫研究中,NcAK可以作为备选的抗虫靶标。

图3 精氨酸激酶蛋白质序列构建的进化树Fig.3 The phylogenetic tree of the deduced protein of NcAK and homologous gene sequences
本试验分离得到了黑尾叶蝉谷胱甘肽-S-转移酶基因NcGST的CDS序列,利用NCBI在线工具 Conserved Domains Search(www.ncbi.nlm.gov/structure/cdd/wrpsb.cgi)预测得到该基因编码的蛋白拥有delta类型的NcGST的结构域,推测NcGST是delta类型的谷胱甘肽-S-转移酶基因。预测的NcGST蛋白序列和美国牧草盲蝽内得到的谷胱甘肽-S-转移酶的氨基酸有74%的相似性。美国牧草盲蝽内的GSTs具有解毒功能,由此推测NcGST也有类似功能。检测表明NcGST在卵和1龄若虫中表达量与在5龄若虫和雌雄成虫中表达量差异显著(P<0.05)。由此可以推测,黑尾叶蝉对有机磷和有机氯类农药的敏感性会随着龄期的增加而降低,这在之前未见报道。而黑尾叶蝉对吡虫啉、阿维菌素和噻嗪酮的敏感性变化中据报道有类似趋势,林雪梅等发现,随着龄期的增长,黑尾叶蝉对吡虫啉、阿维菌素的敏感性会降低[14],在其他昆虫已经证明GSTs的活性与这两种农药在昆虫体内的代谢有关[15-16],所以这种现象也许与NcGST的表达量增加有关,推测NcGST对这类杀虫剂也有解毒能力。黑尾叶蝉对噻嗪酮的敏感性同样也随着龄期的增加而降低,但是在褐飞虱抗性品系的研究中林友伟发现GSTs的活性与这种敏感性的降低无关[17]。黑尾叶蝉对噻嗪酮敏感性的降低可能有其他原因,需要进一步研究。
现在昆虫对农药的敏感性越来越低,发展安全高效的抗虫策略已经迫在眉睫。本次研究中克隆到了黑尾叶蝉的精氨酸激酶和一个谷胱甘肽-S-转移酶的CDS,分析了其序列特征和表达特征,为以后对黑尾叶蝉的研究提供了有用的信息,同时也为以后的抗虫研究打下了基础。

图4 NcGST及其同源基因蛋白质序列构建的进化树Fig.4 The phylogenetic tree of the deduced protein of NcGST and homologous gene sequences

图5 NcAK在不同龄期的表达量Fig.5 The expression levels of NcAK during different stage
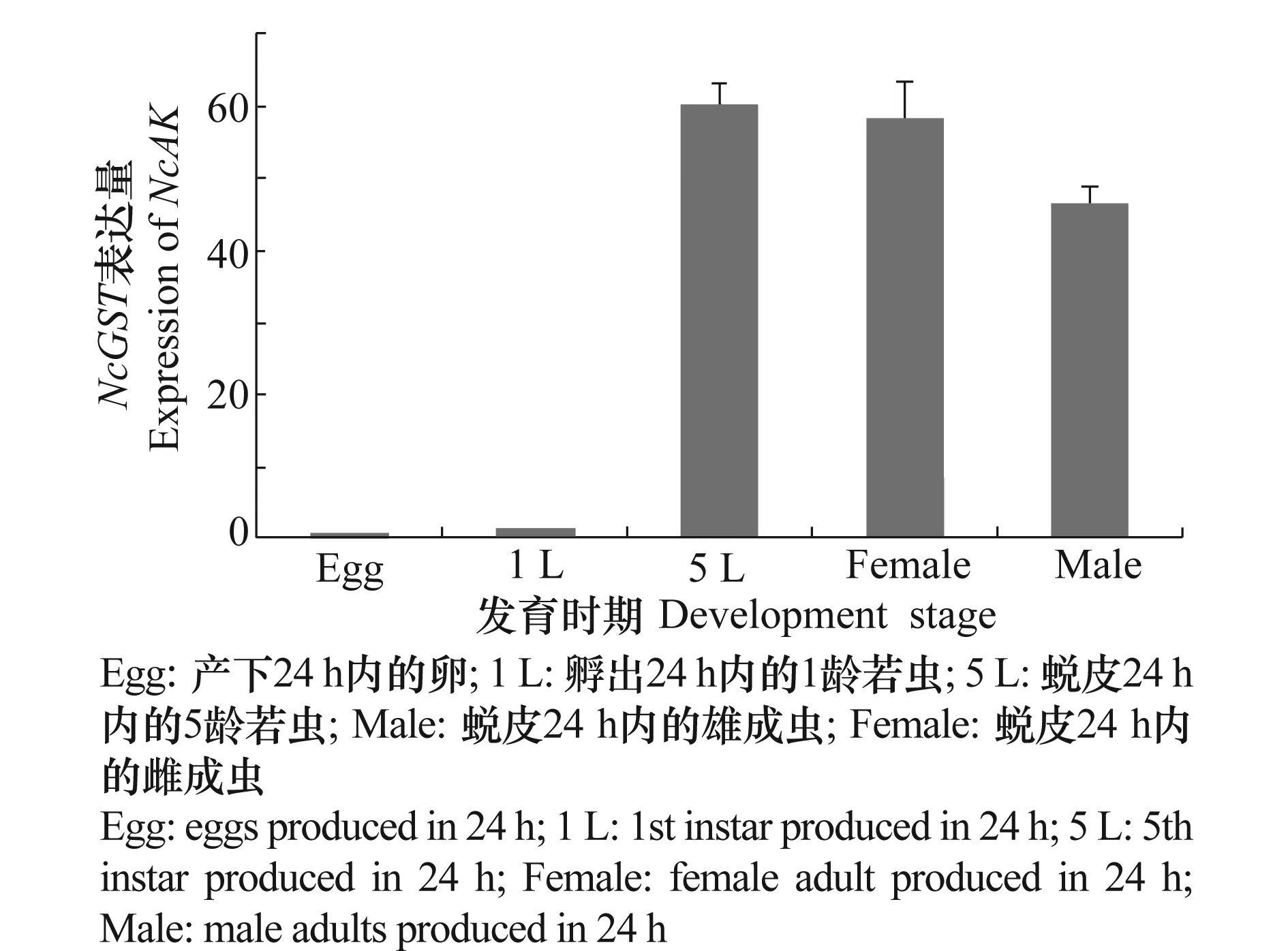
图6 NcGST在不同龄期的表达量Fig.6 The expression levels of NcGST during different stage
[1] 阮义理, 陈声详, 金登迪.传播水稻病毒病的介体黑尾叶蝉发生动态及化学防治[J].昆虫知识, 1985,22(2): 54-57.
[2] Uda K, Fujimoto N, Akiyama Y, et al.Evolution of the arginine kinase gene family[J].Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 2006, 1(2): 209-218.
[3] 姚翠鸾, 王志勇, 相建海.甲壳动物精氨酸激酶的结构与功能[J].中国生物化学与分子生物学报, 2008, 24(3): 203-208.
[4] 王华兵, 徐豫松.家蚕精氨酸激酶基因的克隆, 基因结构与表达分析[J].中国农业科学, 2006, 39(11): 2354-2361.
[5] Dumas C, Camonis J.Cloning and sequence analysis of the cDNA for arginine kinase of lobster muscle[J].Journal of Biological Chemistry, 1993, 268(29): 21599-21605.
[6] Ding Y, Ortellif, Rossiter L C, et al.TheAnophelesgambiaeglutathione transferase supergene family: annotation, phylogeny and expression profiles[J].BMC genomics, 2003, 4(1): 35.
[7] Motoyama N, Dauterman W.Role of nonoxidative metabolism in organophosphorus resistance[J].Journal of Agricultural and Food Chemistry, 1974, 22(3): 350-356.
[8] 陈东亚, 褚宏亮, 杨维芳, 等.德国小蠊谷胱甘肽 S-转移酶和乙酰胆碱酯酶活力与抗药性的关系[J].中国媒介生物学及控制杂志, 2009, 20(6): 522-524.
[9] Zhu Y C, Snodgrass G L, Chen M S.Comparative study on glutathione S-transferase activity, cDNA, and gene expression between malathion susceptible and resistant strains of the tarnished plant bug,Lyguslineolaris[J].Pesticide Biochemistry and Physiology, 2007, 87(1): 62-72.
[10]陈巧云, 姜家良, 林浩, 等.黑尾叶蝉对有机磷的抗性及增效机制的研究[J].昆虫学报, 1978, 21(4): 360-367.
[11]徐秀凤.小菜蛾精氨酸激酶基因 dsRNA 在大肠杆菌中的表达及其效应[D].福州:福建农林大学, 2012.
[12]苏晓峰.精氨酸激酶在棉铃虫中的表达及调控研究[D].北京:中国农业科学院, 2011.
[13]Zhao Y, Yang G, Wang P G, et al.Phyllotretastriolata(Coleoptera: Chrysomelidae): Arginine kinase cloning and RNAi-based pest control[J].European Journal of Entomology, 2008, 105(5): 815-822.
[14]林雪梅, 乔广行.不同龄期稻黑尾叶蝉对杀虫剂的敏感性测定[J].湖南化工, 2000, 30(5): 53-54.
[15]潘文亮, 党志红, 高占林.棉蚜抗吡虫啉品系和敏感品系主要解毒酶活性比较[J].昆虫学报, 2003, 46(6): 793-796.
[16]孟和生, 王开运, 姜兴印, 等.桔全爪螨的抗药性选育及其解毒酶活力变化[J].昆虫学报, 2002, 45(1): 58-62.
[17]林友伟.两种水稻飞虱对吡虫啉和噻嗪酮的抗性及治理研究[D].南京: 南京农业大学, 2004.
CloningandexpressionprofilesofargininekinaseandGSTsofNephotettixcincticeps
Hou Jixiang,Meng Panpan,Ma Weihua,Zhou Fei,Lin Yongjun
(NationalKeyLaboratoryofCropGeneticImprovementinWuhan,HuazhongAgriculturalUniversity,Wuhan430070,China)
The green leafhopperNephotettixcincticepsis a rice insect that can transmit plant viruses.We cloned and analyzed an arginine kinase gene namedNcAKand a delta class GSTs gene namedNcGSTinNephotettixcincticeps.NcAKcontained an open reading frame (ORF) of 1 068 bp in length, and encodes a protein of 356 amino acids.Phylogenetic analysis showed that the deduced protein ofNcAKwas closely related to the arginine kinase ofHomalodiscavitripennisandOncometopianigricans.The real-time PCR results indicated that theNcAKwas expressed higher in male adults than in the eggs, 1st instars, 5th instars, and female adults.NcGST, 899 bp in length, contained an ORF of 651 bp and encoded a protein of 217 amino acids.Phylogenetic analysis showed that the deduced protein ofNcGSTwas closely related to the GSTs ofLyguslineolaris.The real-time PCR results indicated thatNcGSTwas expressed higher in 5th instars, male adults and female adults than in eggs and 1st instars.
Nephotettixcincticeps; arginine kinase; glutathione-S-transferase
2013-05-17
: 2013-06-08
国家重大科技专项(2011ZX08001-001)
S 433.3
: ADOI: 10.3969/j.issn.0529-1542.2014.01.014
* 通信作者 E-mail:yongjunlin@mail.hzau.edu.cn
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
今日农业(2021年15期)2021-11-26
现代养生·上半月(2021年10期)2021-09-24
中外葡萄与葡萄酒(2020年5期)2020-09-25
北京农学院学报(2019年3期)2019-10-14
中国药剂学杂志(网络版)(2019年4期)2019-07-17
水产科学(2019年3期)2019-05-30
分析化学(2018年4期)2018-11-02
四川动物(2018年5期)2018-10-29
猪业科学(2018年7期)2018-01-22
