
外源茉莉酸和水杨酸甲酯对西花蓟马生长发育和种群动态的影响
2014-08-10 12:29郅军锐从春蕾廖启荣
植物保护 2014年6期
牟 峰, 郅军锐, 从春蕾, 廖启荣, 田 甜
(贵州大学昆虫研究所,贵州山地农业病虫害重点实验室,贵阳 550025)
外源茉莉酸和水杨酸甲酯对西花蓟马生长发育和种群动态的影响
牟 峰, 郅军锐*, 从春蕾, 廖启荣, 田 甜
(贵州大学昆虫研究所,贵州山地农业病虫害重点实验室,贵阳 550025)
为明确不同外源因子对菜豆的诱导抗性及对西花蓟马的影响,研究了茉莉酸、水杨酸甲酯、西花蓟马为害、机械损伤诱导处理的菜豆对西花蓟马生长发育和种群动态的影响。结果表明外源茉莉酸、水杨酸甲酯、西花蓟马为害处理菜豆植株显著延长了西花蓟马未成熟期的发育时间,较健康植株分别延长4.72、2.87和4.95 d,也造成西花蓟马的存活率不同程度的下降。西花蓟马在机械损伤处理和健康植株发育时间差异不显著。不同处理均使西花蓟马种群数量明显下降,并且茉莉酸的抑制作用最明显。因此,外源茉莉酸和水杨酸甲酯诱导的菜豆可对西花蓟马生长发育和种群动态产生不同程度的影响,且茉莉酸的诱导抗性要优于水杨酸甲酯。本结果对探索西花蓟马的防治新策略有一定的参考价值。
西花蓟马; 诱导抗性; 发育历期; 种群动态
西花蓟马[Frankliniellaoccidentalis(Pergande)]是原产于北美洲的一种世界著名的危险性害虫[1]。自2003年7月首次在北京发现后,现已分布在我国10多个地区,造成危害的面积和程度不断加重[2-7]。西花蓟马寄主植物广泛,能直接取食60多科500多种植物,而且还能传播番茄斑萎病毒(TSWV)、凤仙花坏死斑病毒(INSV)在内的多种病毒[8-9]。由于西花蓟马具有个体小,隐蔽性强,寄主范围广,繁殖力强,孤雌生殖等生物学特性,造成防治困难[10]。因此对其有效治理必须寻找新的方法。
昆虫取食对植物造成损伤,使植物产生十分复杂的系统传导信号,其中许多信号都可以激发植物的防御反应,茉莉酸和水杨酸信号传导途径是虫害诱导主要防御途径[11-12]。研究表明,在植物上施用外源茉莉酸和水杨酸可以影响昆虫的生长发育[13-14],如喷有茉莉酸的甘蓝植株对小菜蛾发育历期、蛹重以及产卵量有显著的影响[15];外源茉莉酸诱导枸杞苗木后,枸杞蚜若虫发育历期延长,成虫寿命缩短、产仔量下降、蚜虫体重减轻[16];杨扇舟蛾取食经水杨酸甲酯处理的黑杨,其生长发育受阻[17]。同时也有研究表明虫害后的寄主植物对后取食的植食性昆虫的生长发育也有不同程度的影响,如虫害杨树可延长分月扇舟蛾发育历期、降低雌雄性比和产卵量[18],B型烟粉虱为害后的烟草植株对后取食的烟蚜生长发育和繁殖有显著的不利影响[19]。
有关外源信号物质诱导抗性对西花蓟马的影响也有一些报道。如杨帆[20]研究发现,西花蓟马取食茉莉酸甲酯、水杨酸等诱导处理的黄瓜叶片后,发育时间显著延长,蛹重下降,西花蓟马的生存适合度降低。目前有关不同诱导因子如外源信号物质、虫伤、机械损伤等对西花蓟马生长发育、种群动态的影响的研究还不是很全面,而且试验多数是在离体叶片上进行的。本试验以整株菜豆为供试植物,研究茉莉酸、水杨酸甲酯等不同处理的菜豆植株对西花蓟马生长发育和种群动态的影响,为害虫新的防治措施提供依据。
1 材料与方法
1.1 供试昆虫和植物
西花蓟马:采自贵阳市白云区玫瑰园切花月季上,并于实验室内利用菜豆(PhaseolusvulgarisLinn.)饲养。在条件为(25±1)℃,湿度(70±1)%,光照14 L∥10D的人工气候箱中饲养多代后供试。
菜豆植株:在相同条件下的人工气候箱内培养菜豆植株,以营养钵单株培育清洁苗,待菜豆长至2片真叶期时作为供试寄主植物,生长期间不使用任何农药。
1.2 菜豆植株处理
供试植物参照Thaler[21]和van Dam等[22]的方法,设置如下5个不同处理:
(1)虫伤,记为DTF:在供试菜豆植株每片叶接种10头虫龄一致的西花蓟马2龄若虫,每株共20头,作为西花蓟马为害后的植株,接虫1 d后剔除若虫。
(2)机械损伤,记为MD:供试菜豆植株每叶用针头(直径0.29 mm)从正面穿透10次,1 d后用于试验。
(3)茉莉酸诱导,记为JA:在供试菜豆植株上喷洒浓度为1.0 mmol/L的茉莉酸,喷至叶片全湿为准,1 d后用于试验。
(4)水杨酸甲酯诱导,记为MeSA:在供试菜豆植株上喷洒浓度为5 mmol/L的水杨酸甲酯,喷至叶片全湿为准,1 d后用于试验。
(5)健康植株,记为CK:健康菜豆植株作为对照。
1.3 试验方法
1.3.1 不同处理菜豆植株对西花蓟马生长发育的影响
将已羽化3 d并交配的西花蓟马雌成虫分别接在不同处理菜豆植株上(10头/株),用养虫笼罩住处理植株,任其取食,1 d后移去成虫。每日早、晚06:30观察植株叶片中卵的孵化情况,发现初孵若虫后,立即将其移到叶子正面,并用加拿大树脂和蓖麻子油(V∶V=1.5∶1)把若虫圈起来,留有适当空间供其取食。叶片上1头若虫作为一个重复,每个处理设置30个重复,试验在相同条件的人工气候箱中进行。每日早、晚06:30观察记录各种处理中若虫的发育情况,记录从初孵若虫至羽化所需时间(h),并记载其发育为雌雄的情况。
1.3.2 不同处理菜豆植株对西花蓟马种群动态的影响
选取羽化且交尾3 d的西花蓟马雌成虫,每株10头,分别接于已处理好的不同处理菜豆植株上,于接虫后第10天进行害虫种群调查。调查时将菜豆植株整株剪下,立即放入自封袋(40 cm×60 cm)中,计数菜豆上西花蓟马成虫和若虫的数量。另外在接入西花蓟马的菜豆植株剪下后在营养钵中插一张黄色粘虫板,将笼罩扣回营养钵上方,7 d后收集土壤中的蛹羽化出来的西花蓟马成虫,计入西花蓟马的总数量中。试验在相同条件的人工气候箱中进行,设置3次重复。
1.4 数据统计分析
试验数据采用Excel 2003和SPSS 19进行统计和分析,利用Duncan氏新复极差法检验不同诱导因子对西花蓟马生长发育和种群动态影响的差异显著性。
2 结果与分析
2.1 不同诱导因子对西花蓟马生长发育的影响
从表1可以看出不同诱导处理的菜豆植株对西花蓟马各虫态及未成熟期的发育时间有不同程度的影响。西花蓟马的卵期和2龄若虫期在虫害植株的发育时间最长,1龄若虫以茉莉酸诱导处理的发育时间最长。预蛹期和蛹期是不食不动的时期,但不同处理对它们的生长发育也有影响,是前期处理的累积效应所致,但不同处理间的差异明显变小,茉莉酸、水杨酸甲酯、虫伤3种处理之间没有显著差异,但均高于健康植株和机械损伤植株。茉莉酸、水杨酸甲酯、虫伤处理的菜豆植株上的西花蓟马未成熟期的历期分别为17.68、15.83和17.91 d,均显著长于健康植株上西花蓟马的发育时间,说明茉莉酸、水杨酸甲酯、虫伤3种处理均能够有效抑制西花蓟马的发育,又以虫伤和茉莉酸的抑制效果高于水杨酸甲酯,而机械损伤与健康植株相比,发育历期差异不大。

表1 不同外源诱导因子处理下西花蓟马的发育历期1)Table 1 The development durations of Frankliniella occidentalis under different treatments with exogenous factorsd
1) 表中数据为平均值±标准误,同一列中小写字母不同表示发育历期在不同处理之间达到显著水平(P<0.05;Duncan氏新复极差测验法)。
Data in the table are mean±SE; different lowercase letters in the same column indicate the significant level at 5%.
西花蓟马各个虫期在不同处理菜豆植株上存活率也有差异,如图1所示。西花蓟马各龄期的存活率,以健康植株和机械损伤处理最高,在健康植株上最低的存活率也高达96.67%,水杨酸甲酯处理次之,虫伤和茉莉酸处理各龄期存活率最低。不同虫态在不同处理之间也有一定的差异,西花蓟马1龄和2龄若虫在茉莉酸诱导处理的植株上的存活率高于虫伤处理,但在预蛹期与虫伤处理存活率差异不大,在蛹期又低于虫伤处理的。
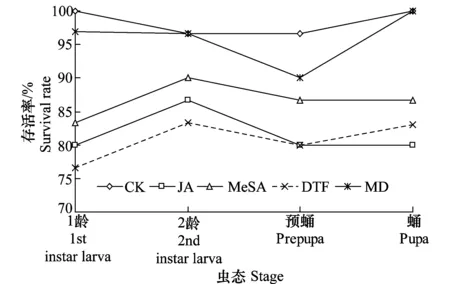
图1 西花蓟马在不同外源诱导因子处理菜豆上的存活率Fig.1 The survival rate of Frankliniella occidentalis on treated bean plants with different exogenous factors
2.2 不同诱导因子对西花蓟马种群数量的影响
虫伤、茉莉酸和水杨酸甲酯、机械损伤都能有效地抑制西花蓟马的种群数量(表2),其中尤以虫伤和茉莉酸处理的西花蓟马种群数量最低,分别只有健康植株的25.68%和27.03%。机械损伤处理种群数量也显著低于健康植株上西花蓟马的数量,但又显著高于其他3种处理。
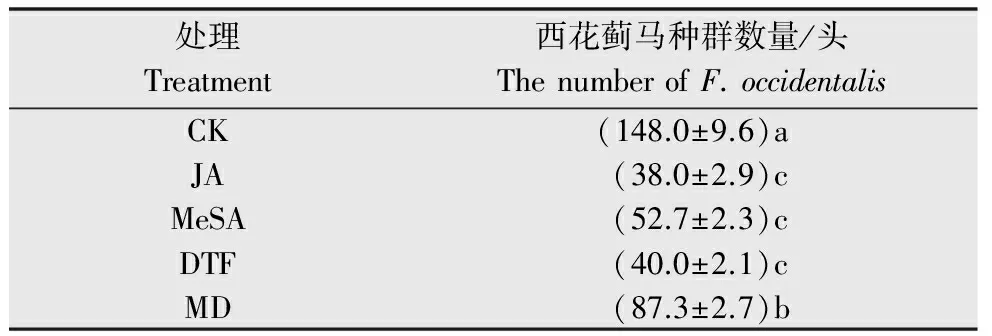
表2 不同外源诱导因子处理下西花蓟马种群动态1)Table 2 Population dynamics of Frankliniellaoccidentalis under different treatments with exogenous factors
1) 表中数据为平均值±标准误,同一列中小写字母不同表示种群数量在不同处理下达到显著水平(P<0.05;Duncan氏新复极差测验法)。
Data in the table are mean±SE; different lowercase letters in the same column indicate the significant level at 5%.
3 讨论
本试验研究结果表明,茉莉酸、水杨酸甲酯以及虫伤处理的菜豆植株使西花蓟马生长发育时间延长,存活率下降。说明茉莉酸、水杨酸甲酯以及虫伤处理的菜豆对西花蓟马产生了明显的防御反应,证明了茉莉酸、水杨酸甲酯作为外源化学物质都能够诱导植物产生抗性,从而使植物对西花蓟马的生长发育产生显著的影响,且茉莉酸诱导的植物抗性强于水杨酸甲酯诱导的抗性,这与杨帆[20]的研究结论基本一致。这种诱导抗性可能是外源信号物质茉莉酸、水杨酸甲酯激发了菜豆叶片防御基因的表达,改变了次生代谢过程,产生了对西花蓟马有毒、抗营养或抗消化作用的化合物,这些化合物会对害虫的生长发育等产生不利影响[23-24]。从春蕾[25-26]研究表明西花蓟马取食为害后菜豆植株叶片内营养物质有不同程度的下降,防御酶活性、防御基因表达有不同程度的升高。
茉莉酸、水杨酸甲酯和虫伤处理的种群数量在接虫后10 d明显低于健康对照的,说明外源茉莉酸、水杨酸甲酯及虫害植株对西花蓟马繁殖力产生了影响,降低了西花蓟马的种群数量。国外学者也有相关报道,如Abe等[27]的研究发现,西花蓟马取食后野生型拟南芥茉莉酸的含量明显升高,并且在西花蓟马为害野生型拟南芥之前喷洒茉莉酸会增加植物对蓟马取食的耐受性。Abe等[28]的研究还发现在西花蓟马为害野生型拟南芥之前喷洒茉莉酸会减少该虫的种群数量。De Vos等[29]的研究也证明西花蓟马的为害促进茉莉酸的合成。因此,外源信号物质的诱导抗性对抑制西花蓟马的产卵量、降低其种群密度具有重要意义。
茉莉酸、水杨酸诱导植物抗虫性还与喷施浓度有关。田旭涛等[30]研究了4种浓度茉莉酸处理菜豆对美洲斑潜蝇的影响,发现高浓度茉莉酸处理菜豆使美洲斑潜蝇幼虫发育历期延长、取食量降低、蛹重减轻,而低浓度茉莉酸处理菜豆影响较小,仅对美洲斑潜蝇发育历期有影响。杨帆[20]研究发现,茉莉酸甲酯处理对西花蓟马发育的抑制作用随浓度增加而增加,水杨酸处理在一定浓度范围内对西花蓟马发育的抑制作用也是随浓度增加而增加。
害虫为害密度不同对其他害虫的影响程度也不同,Lynch等[31]研究发现马铃薯小绿叶蝉为害程度越高对后取食的马铃薯甲虫幼虫的发育和产卵影响越大;李海林[32]的研究表明不同受害程度的马尾松对马尾松毛虫生长发育影响不同。本试验只研究一个浓度茉莉酸、水杨酸甲酯以及一个蓟马密度为害条件下对西花蓟马生长发育和种群动态的影响,不同浓度下茉莉酸、水杨酸甲酯以及不同为害条件下对西花蓟马生长发育以及种群动态的影响还有待进一步研究。另外,西花蓟马取食不同处理诱导抗性植株后,对西花蓟马体内相关物质如解毒酶、保护酶、消化酶有何影响,也需进一步研究与探讨。
[1]Kirk W D J, Terry L I. The spread of the western flower thripsFrankliniellaoccidentalis(Pergande)[J]. Agricultural and Forest Entomology, 2003, 5(4):301-310.
[2]张友军, 吴青君, 徐宝云, 等. 危险性外来入侵生物——西花蓟马在北京发生危害[J]. 植物保护,2003,29(4):58-59.
[3]袁成明, 郅军锐, 郑珊珊, 等. 西花蓟马在贵阳地区发生危害调查研究[J]. 西南师范大学学报(自然科学版), 2010, 35(1):142-145.
[4]杨华, 崔元玗, 张升,等. 外来入侵害虫——西花蓟马在新疆的发生为害[J]. 新疆农业科学, 2010, 47(11):2252-2253.
[5]郑长英, 刘云虹, 张乃芹, 等. 山东省发现外来入侵有害生物—西花蓟马[J]. 青岛农业大学学报(自然科学版), 2007(3):172-174.
[6]任洁, 雷仲仁, 张令军, 等. 北京地区西花蓟马发生为害调查研究[J]. 中国植保导刊, 2006, 26(5):5-7.
[7]王海鸿, 雷仲仁, 李雪, 等. 西藏发现重要外来入侵害虫——西花蓟马[J]. 植物保护, 2013,39(1):181-183.
[8]Moritz G. The biology of thrips is not the biology of their adults:a developmental view[C]∥Marullo R, Mound L A,eds. Thrips and tospoviruses. Proceeding of the 7th international symposium on Thysanoptera. Canberra:Australian National Insect Collection, Regio Calabria Italy,2002:259-267.
[9]Ullman D E, Meideros R, Campbell L R, et al. Thrips as vectors of tospoviruses[J]. Advances in Botanical Research, 2002, 36:113-140.
[10]Contreras J, Espinosa P J, Quinto V, et al. Stability of insecticide resistance inFrankliniellaoccidentalisto acrinathrin, formetanate and methiocarb[J]. Agricultural and Forest Entomology, 2008, 10(3):273-278.
[11]Kessler A, Baldwin I T. Plant responses to insect herbivory:the emerging molecular analysis[J]. Annual Review of Plant Biology, 2002, 53(1):299-328.
[12]Zhu-Salzman K, Saizman R A, Ahn J E, et al. Transcriptional regulation of sorghum defense determinants against a phloem-feeding aphid[J]. Plant Physiology, 2004, 134(1):420-431.
[13]Baldwin I T. Jasmonate-induced responses are costly but benefit plants under attack in native populations[J]. Proceedings of the National Academy of Sciences, 1998, 95(14):8113-8118.
[14]Sano H, Seo S, Orudgev E, et al. Expression of the gene for a small GTP binding protein in transgenic tobacco elevates endogenous cytokinin levels, abnormally induces salicylic acid in response to wounding, and increases resistance to tobacco mosaic virus infection[J]. Proceedings of the National Academy of Sciences of the United States of America, 1994, 91(22):10556-10560.
[15]吕要斌, 刘树生. 外源茉莉酸诱导植物反应对小菜蛾生长发育的影响[J].应用生态学报, 2005, 16(1):193-195.
[16]宫玉艳, 段立清, 王爱清, 等.外源茉莉酸诱导枸杞对枸杞蚜生长发育和繁殖的影响[J]. 昆虫学报,2010,53(6):670-674.
[17]赵玲, 胡增辉, 赵风君, 等. 水杨酸甲酯诱抗黑杨对杨扇舟蛾生长发育的影响[J].北京林业大学学报, 2005,27(1):75-78.
[18]周艳琼, 钱为, 郝德君, 等. 虫害诱导黑杨对分月扇舟蛾生长发育的影响[J]. 昆虫知识, 2010, 47(6):1150-1156.
[19]毕明娟. B型烟粉虱诱导的烟草防御信号途径及B型烟粉虱和烟蚜对烟草防御反应的生理适应性差异[D]. 泰安:山东农业大学, 2010.
[20]杨帆. 水杨酸、茉莉酸甲酯诱导黄瓜对西花蓟马(缨翅目:蓟马科)的抗性[D]. 武汉:华中农业大学, 2009.
[21]Thaler J S, Stout M J, Karban R, et al. Jasmonate‐mediated induced plant resistance affects a community of herbivores[J]. Ecological Entomology, 2001, 26(3):312-324.
[22]van Dam N M, Oomen M. Root and shoot jasmonic acid applications differentially affect leaf chemistry and herbivore growth[J]. Plant Signaling & Behavior, 2008, 3(2):91-98.
[23]Agrawal A A, Karban R. Specificity of constitutive and induced resistance:pigment glands influence mites and caterpillars on cotton plants[J]. Entomologia Experimentalis et Applicata, 2000, 96(1):39-49.
[24]桂连友, 刘树生, 陈宗懋. 外源茉莉酸和茉莉酸甲酯诱导植物抗虫作用及其机理[J]. 昆虫学报, 2004, 47(4):507-514.
[25]从春蕾, 郅军锐, 谢路飞, 等. 西花蓟马为害对菜豆叶绿素及营养物质含量的影响[J]. 植物保护, 2013, 39(2):20-24.
[26]从春蕾, 郅军锐, 谢路飞, 等. 西花蓟马取食对菜豆防御基因表达的诱导作用[J]. 昆虫学报, 2013(10):1174-1180.
[27]Abe H, Ohnishi J, Narusaka M, et al. Function of jasmonate in response and tolerance of Arabidopsis to thrip feeding[J]. Plant and Cell Physiology,2008, 49(1):68-80.
[28]Abe H, Shimoda T, Ohnishi J, et al. Jasmonate-dependent plant defense restricts thrips performance and preference[J]. BMC Plant Biology, 2009, 9(1):97.
[29]de Vos M, van Oosten V R, van Poecke R M P, et al. Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack [J]. Molecular Plant-microbe Interactions,2005, 18(9):923-937.
[30]田旭涛, 张箭, 李丹, 等. 茉莉酸处理菜豆对美洲斑潜蝇抗性的影响[J]. 植物保护学报, 2013, 40(4):345-349.
[31]Lynch M E, Kaplan I, Dively G P, et al. Host-plant-mediated competition via induced resistance:interactions between pest herbivores on potatoes[J]. Ecological Applications, 2006, 16(3):855-864.
[32]李海林. 不同受害程度的马尾松对马尾松毛虫种群数量影响的研究[D]. 北京:北京林业大学, 2006.
EffectsofexogenousjasmonicacidandmethylsalicylateondevelopmentaldurationandpopulationdynamicsofFrankliniellaoccidentalis
Mou Feng, Zhi Junrui, Cong Chunlei, Liao Qirong, Tian Tian
(InstituteofEntomology,GuizhouUniversity,GuizhouProvincialKeyLaboratoryforAgriculturalPestManagementoftheMountainousRegions,Guiyang550025,China)
In order to explore the induced resistance of bean plants by different exogenous factors and its effects onFrankliniellaoccidentalis, the influences of bean plants treated with jasmonic acid(JA), methyl salicylate(MeSA), thrips-infested and mechanical damage on the developmental duration and population dynamics ofF.occidentaliswere studied. The results showed that the developmental duration of immature stage ofF.occidentaliswas significantly prolonged when treated with exogenous JA, MeSA and thrip-infestation, with 4.72, 2.87 and 4.95 d longer than that of the healthy plant, respectively, and the survival rate ofF.occidentalisdecreased to different extents. The difference in the development duration ofF.occidentalisbetween healthy and mechanically damaged plants was insignificant. The population dynamics ofF.occidentalissignificantly decreased under different treatments, and JA had the most significant inhibitory effect. Therefore, the bean plants induced by exogenous JA and MeSA had effects on the developmental duration and population dynamics ofF.occidentalis, and the effect of induced resistance by JA was bigger than that of MeSA.These results could provide guide for exploring new control strategies forF.occidentalis.
Frankliniellaoccidentalis; induced resistance; developmental duration; population dynamics
2013-12-05
:2014-04-07
国家自然科学基金(31160367)
S 433.89
:ADOI:10.3969/j.issn.0529-1542.2014.06.013
* 通信作者 E-mail:zhijunrui@126.com
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
今日农业(2021年11期)2021-11-27
农业资源与环境学报(2021年5期)2021-10-06
生物安全学报(2021年3期)2021-09-06
河南农业大学学报(2020年3期)2020-07-09
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
中国烟草学报(2016年3期)2016-11-23
故事大王(2014年3期)2014-05-21
探索财富(2013年10期)2013-11-18
