
巨大芽胞杆菌B196菌株抑菌物质的分离纯化
2014-08-10 12:29袁高庆黎起秦彭好文
植物保护 2014年2期
廖 庭,秦 健,袁高庆,黎起秦,林 纬,彭好文
(广西大学农学院,南宁 530005)
研究报告
巨大芽胞杆菌B196菌株抑菌物质的分离纯化
廖 庭,秦 健,袁高庆,黎起秦*,林 纬,彭好文
(广西大学农学院,南宁 530005)
巨大芽胞杆菌(Bacillusmegaterium)B196菌株产生的拮抗物质对水稻纹枯病菌(Rhizoctoniasolani)的生长具有较强的抑制作用,明确该菌产生拮抗物质的种类是进一步研究该菌的抑菌机制及其应用的基础。本文采用盐酸沉淀B196菌株的去菌体培养液,再用甲醇抽提获得拮抗物质的粗提物。利用反相HPLC系统,将粗提物过C18柱,收集有抑制水稻纹枯病菌生长作用的活性化合物。运用质谱测得其分子量分别为1042.592 7 u,再利用碰撞诱导解离(CID)技术获得化合物的典型结构特征离子碎片,结果表明其一级结构为Pro-Asn-Ser-βAA-Asn-Tyr-Asn-Gln (βAA为14个碳原子的氨基脂肪酸)。综合以上信息将该化合物鉴定为Iturin A2。
巨大芽胞杆菌; iturin A2; 分离纯化
巨大芽胞杆菌(Bacillusmegateriumde Bary)是一类好氧、革兰氏阳性、可生长在土壤和植物体内的芽胞杆菌细菌,它可以被用来吸附金属Au3+[1]、净化水体[2]、降解农药[3]、解磷解钾[4]等,被广泛地应用在环境保护方面。此外,巨大芽胞杆菌还可以产生抑制植物病原菌的抗菌物质[5],因而在植物病害防治方面也受到人们的关注。目前已有报道分离出一些对植物病原菌有较强拮抗作用的菌株[6],一些菌株对一些植物病害有较好的防治作用[7],巨大芽胞杆菌可产生的抑菌物质有蛋白类[5]、挥发性物质[8]、细菌素[9]和脂肽类物质等[10]。
巨大芽胞杆菌B196菌株是广西大学农学院植物病理研究室从水稻上分离出来的拮抗细菌,在离体、盆栽和大田试验中,该菌株对水稻纹枯病均表现出较好的防效[11]。前期工作发现,该菌株的发酵滤液对玉米小斑病菌[Bipolarismaydis(Nisik & Miyake)Shoem.] 和水稻纹枯病菌(RhizoctoniasolaniKühn)具有较强的抑菌活性[12]。为了明确该菌株的发酵滤液所含的拮抗物质种类,本研究采用盐酸沉淀、甲醇提取和高效液相色谱分离等方法对抑菌物质进行分离纯化和鉴定,为进一步研究该菌的抑菌机制和开发防治水稻纹枯病的新型药剂打下基础。
1 材料与方法
1.1 材料
1.1.1 菌株
拮抗菌:巨大芽胞杆菌(B.megaterium)B196菌株;植物病原菌:稻瘟病菌[Magnaporthegrisea(Hebert) Barr.]、烟草赤星病菌[Alternariaalternata(Fries) Keissler ]、烟草灰霉病菌(BotrytiscinereaPers.)、水稻纹枯病菌(R.solani)、玉米白绢病菌(SclerotiumrolfsiiSacc.)、西瓜枯萎病菌[Fusariumoxysporumf.sp.niveum(E.F.Smith) SnyderetHansen]和黄瓜绵腐病菌[Pythiumaphanidermatum(Edson) Fitzpatrick],以上病原菌均来源于广西大学农学院植病研究室。
1.1.2 色谱分离材料
分析柱C18(Jupiter,5 μm,250mm×4.6mm)、制备柱C18(PRC-ODS,20mm×250mm)、离心浓缩机(Labconco CentriVap)、高效液相色谱仪(Shimadzu LC-6AD)为岛津公司生产,乙腈、乙酸铵均为色谱级。
1.2 拮抗物质的分离纯化
1.2.1 B196菌株的培养及其抑菌物质的提取
以水稻纹枯病菌为靶标,采用活性跟踪分离纯化B196菌株产生的拮抗物质[13]。将保存在4 ℃的B196菌株接种于NA培养基上,在30℃培养48 h后取一环B196菌株接种至种子培养基(葡萄糖10g、大豆蛋白胨10g、牛肉浸膏3g、蒸馏水1000mL、pH7.0)中,30℃、130r/min下培养12 h后再转移至发酵培养基(蔗糖10g、大豆蛋白胨20g、NaCl 7 g、K2HPO45 g、蒸馏水1000mL、pH 7.0),27 ℃,130r/min,振荡培养5 d。之后将B196菌株的发酵液6000r/min离心20min,去除菌体,收集上清液,用6mol/L的HCl将上清液的pH调为2.0,4 ℃下静置24 h后,于8 000r/min离心25 min,去除上清液,收集沉淀。合并所有的盐酸沉淀,加入适量的甲醇,将pH调为7.0,静置2 h,抽提甲醇,于60℃下进行减压浓缩,获得深黄色的粗提物,备用。
1.2.2 抗菌物质的抑菌活性测定方法
采用菌丝生长速率法:将分离纯化获得的各馏分于1/15 mol/L磷酸缓冲液(pH6.81)中溶解,取0.1mL各馏分与5 mL的PSA培养基(45 ℃左右)混匀,倒入6cm的培养皿中,冷却后备用。用直径为7 mm的打孔器在水稻纹枯病菌菌落边缘切取菌饼,将病菌反向接入培养皿中央,在28 ℃下恒温培养24 h后测量菌落直径,并计算抑菌率,以磷酸缓冲液和清水处理为对照,每处理3次重复。
1.2.3 HPLC分析
用流动相溶解粗提物,采用等度洗脱,液相色谱条件:色谱柱C18(Jupiter, 5 μm,250mm×4.6mm),流动相含乙腈10mmol/L,V(乙酸铵)∶V(水溶液)=40∶60,波长为214 nm,流速为0.8 mL/min,室温,样品浓度为10mg/mL,进样量为10μL。收集各色谱峰样品,用离心浓缩机浓缩和干燥,以水稻纹枯病菌为靶标菌,采用菌丝生长速率法检测各色谱峰样品的抑菌活性。
1.2.4 HPLC分离纯化
分离纯化条件:流动相含乙腈10mmol,V(乙酸铵)∶V(水溶液)=40∶60,波长214 nm,流速为8 mL/min,制备柱C18(PRC-ODS,20mm×250mm),样品用流动相液溶解,浓度为20mg/mL,进样量1mL,集中收集活性峰,用离心机浓缩和冷冻干燥后获得活性物质。
1.3 活性组分的鉴定
样品送至广西大学生命科学与技术学院,采用反射模式采集每个样品的一级质谱信息(MS),激光强度4 000,质量范围800~4 000u,一级质谱扫描完成后,选取7个信号强度最高的母离子进行二级质谱分析(MS/MS),加速电压2 kV,采用CID碰撞裂解母离子,获取每个母离子的离子碎片指纹谱。
1.4 纯化合物对植物病原菌的毒力测定
采用菌丝生长速率法测定化合物对植物病原真菌的毒力:用1/15 mol/L磷酸缓冲液(PBS)溶解纯品后分别配制成5种浓度,取0.1mL纯品溶液与5 mL的PSA培养基混匀,倒入6cm的培养皿中,冷却后接入植物病原菌,以磷酸缓冲液和清水为对照,在28 ℃下恒温培养。待对照菌落基本长满培养皿后测量菌落直径,并计算抑菌率,每处理3次重复。
2 结果与分析
2.1 HPLC分析
粗提物经分析柱C18(Jupiter,5 μm,250mm×4.6mm)分离的色谱图见图1。在8.4 min时出现一个呈正态分布的峰形,从该峰收集的样品对水稻纹枯病菌有抑菌活性。
2.2 HPLC分离纯化
样本经HPLC分离纯化后,分别在3.9~6.2 min和16.4~17.2 min集中出现洗脱峰(图2),抑菌活性检测结果表明,在16.4~17.2 min出现的洗脱峰物质,在浓度为100μg/mL时,对水稻纹枯病菌的抑菌率为81.03%,分离纯化的物质经离心机浓缩和冷冻干燥后得到的白色粉末即为纯品,且纯拮抗物质的提取率为5%。

图1 B196菌株产生的活性物质在分析柱中的色谱图Fig.1 HPLC chromatograms of the active substances produced by Bacillus megatherium strain B196

图2 B196菌株产生的抑菌物质在HPLC上的分离纯化Fig.2 Purification of the active substance produced by the strain B196on HPLC
2.3 活性物质的纯度
已分离纯化的物质在分析色谱柱C18(Jupiter,5 μm,250mm×4.6mm),流动相含乙腈10mmol/L,V(乙酸铵)∶V(水溶液)=40∶60,波长为214 nm,流速为0.8 mL/min 时检测其纯度,结果见图3。在8.4 min时出现一个呈正态分布的峰形,从该峰收集的样品对水稻纹枯病菌有抑菌活性。该洗脱峰与2.1HPLC分析中在8.4 min时出现的呈正态分布峰形一致。根据面积归一法获知其面积大于94%,说明分离获得的拮抗物质纯度已达94%以上,可以进行质谱鉴定。

图3 分离纯化的拮抗物质色谱图Fig.3 HPLC chromatograms of the active substance purified from the strain B196
2.4 活性组分的鉴定
利用基质辅助激光解析电离飞行时间质谱(MALDI-TOF-MS)对化合物进行一级质谱分析(图4),获知该化合物的质荷比(m/z)为1043.592 7、1065.579 1和1081.554 1,它们分别是化合物的质子化(M+H+)峰、钠离子(M+Na+)和钾离子(M+K+)加合峰,结果表明该化合物的分子量为1042.592 7 u。将此质谱图结果与前人研究结果[14-16]进行比对时,可以初步估计分子量为1042.592 7 u的化合物是伊枯草菌素A2。
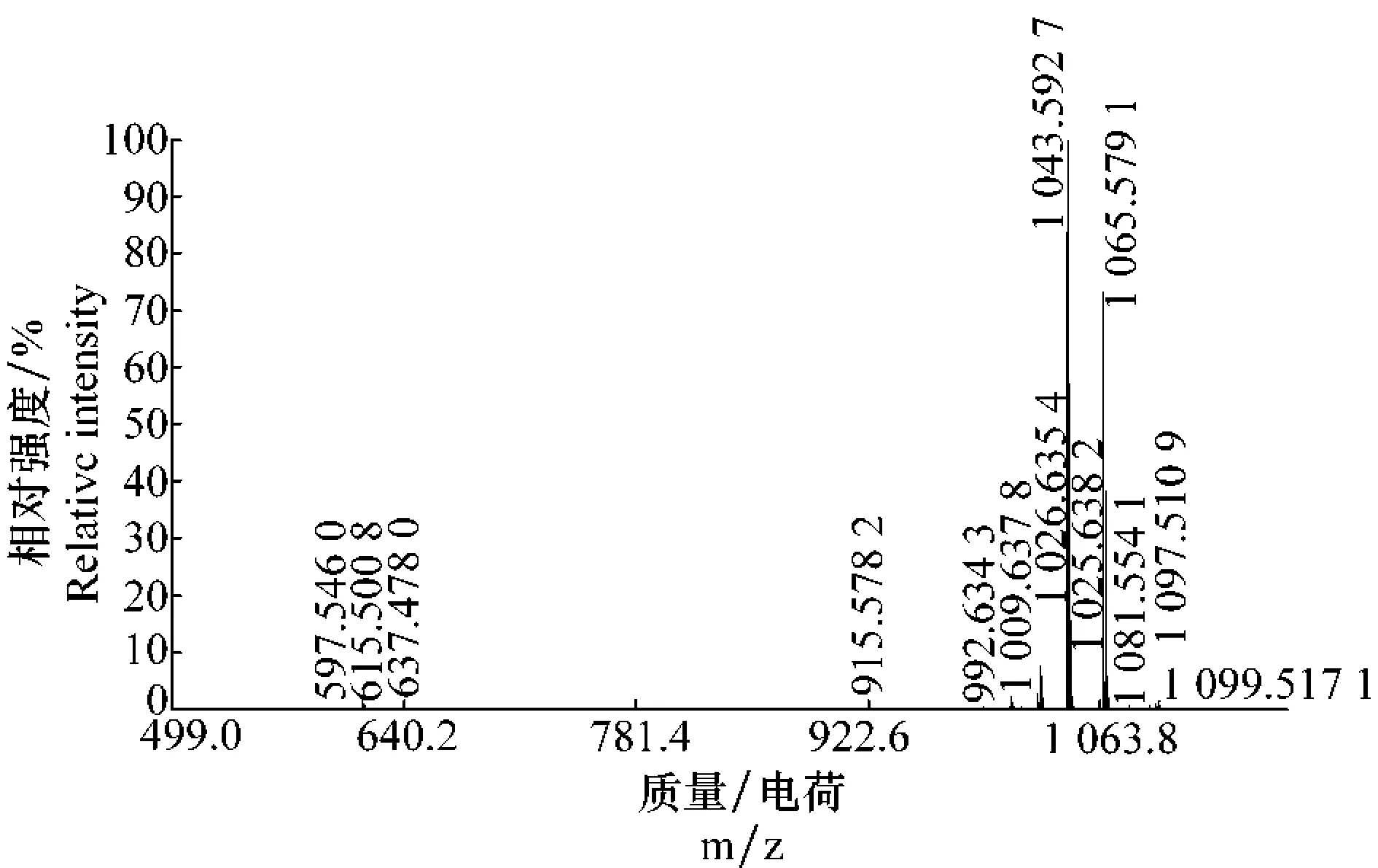
图4 纯品的基质辅助激光解析电离飞行时间质谱图Fig.4 MALDI-TOF-MS spectrum of the purified substance

图5 纯品的MALDI-TOF-MS/MS CID色谱图Fig.5 MALDI-TOF-MS/MS CID spectrum of the purified substance
多肽类物质被电离后,会形成多种强度不同的离子,据Roepstorff P等[17]和Biemann K等[18]的报道:当电荷保留在N-端时,会从多肽类物质N-端3个裂解点开始分别形成an、bn、cn系列离子,且bn系列离子的质量数,是断裂后的基团的质量数;当电荷保留在N-端时,会从多肽类物质N-端3个裂解点开始依次形成zn、yn和xn系列离子,且yn系列离子的质量数,是断裂后的基团的质量数加上2H。其中bn系列和yn系列离子是确定氨基酸序列的关键离子,其他离子属于离子片段。
纯品的二级质谱(图5),参照Yu G Y等[16]描绘的模式,先将伊枯草菌素A2的一级结构排列出来,然后分成a型离子、b型离子、c型离子、x型离子、y型离子和z型离子(图6)。从图5中可以获得的a型离子、b型离子、c型离子、x型离子、y型离子和z型离子数据(表1)。因为b类型是从左至右将结构残基切断,而y类型则是由右到左将结构残基切断,所以通过图5中b、y型离子之间的差值与氨基酸残基分子量的比较,可以获得片段结构分别为Ser-βAA-Asn-Tyr-Asn-Gln、Pro-Asn-Ser-βAA-Asn-Tyr。而Ser-βAA-Asn-Tyr为这两个结构片段的共同部分,所以结合这两个片段结构推断出纯品的一级结构为Pro-Asn-Ser-βAA-Asn-Tyr-Asn-Gln。因为βAA的质荷比m/z=b4-b3=y5-y4=225,故可以推断此氨基脂肪酸链应该含有14个碳原子,且其质子化结构为-H2N+=CH-C12H25-CO-。
综合纯品的分子量和MALDI-TOF-MS/MS CID色谱图等信息,确认其为伊枯草菌素家族的iturin A2。

图6 Iturin A2质子化(M+H+)的诱导碰撞解离碎片[16]Fig.6 Possible fragments of iturin A2(M+H)+by CID
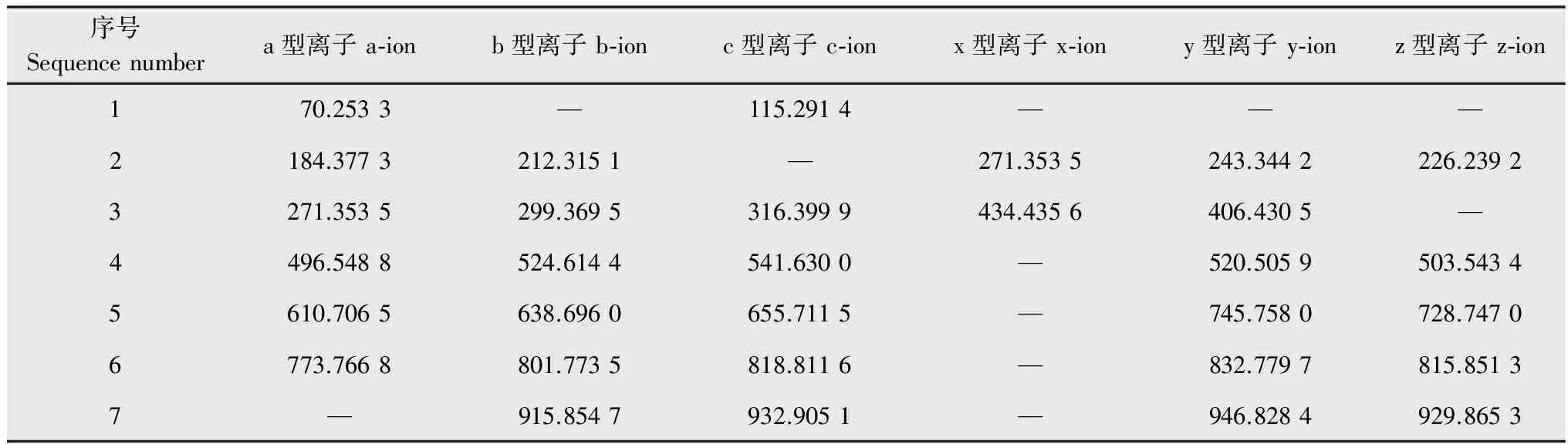
表1 纯品的MS/MS数据Table 1 Mass spectrometry/mass spectrometry (MS/MS) data of the purified substance
2.5 纯化合物对植物病原菌的毒力
采用菌丝生长速率法测定纯化合物对7种植物病原菌的毒力结果见表2。结果表明:纯化合物对植物病原菌普遍具有较高的抑菌作用。其中,纯品对稻瘟病菌的毒力最强,EC50达到了11.9700μg/mL;其次为烟草赤星病菌、烟草灰霉病菌、水稻纹枯病菌、玉米白绢病菌和西瓜枯萎病菌;对黄瓜绵腐病菌的毒力最弱,EC50为102.059 2 μg/mL。
表2纯化物质对植物病原菌的毒力1)
Table2Toxicityofthepurifiedsubstanceagainstplantpathogenicfungi

植物病原菌Plantpathogenicfungi毒力回归方程(Y=)Regressionequationoftoxicity抑菌中浓度(EC50)/μg·mL-1Medianeffectiveconcentration95%置信区间/μg·mL-195%Confidenceinterval相关系数Correlationcoefficient稻瘟病菌Magnaporthegrisea2.6980X+2.091411.9700 8.3459~17.1678 0.9533烟草赤星病菌Alternariaalternata1.3295X+1.329512.38169.2100~16.6455 0.9674烟草灰霉病菌Botrytiscinerea1.5111X+2.970222.048015.8952~30.5826 0.9626水稻纹枯病菌Rhizoctoniasolani2.8910X+0.382339.562132.6826~47.8897 0.9863玉米白绢病菌Sclerotiumrolfsii1.9371X+1.872041.186928.7190~59.0674 0.9554西瓜枯萎病菌Fusariumoxysporum1.9605X+1.537558.359242.6268~79.9166 0.9736黄瓜绵腐病菌Pythiumaphanidermatum1.9711X+1.0404102.059269.0827~150.7769 0.9767
1) 数据为3次重复的平均值;EC50为根据毒力方程的计算值。 The data were the mean of 3replicates.EC50was the value calculating from the virulence equation.
3 结论与讨论
对于脂肽化合物的分离纯化,一般都要经过盐酸沉淀、甲醇提取、硅胶柱层析、凝胶柱层析和高效液相色谱等方法才能获得纯品[13,16,19],而本研究直接采用高效液相色谱对拮抗菌代谢产物的甲醇提取物进行分离纯化,并获得纯度较高的iturin A2,简化了步骤。陈华等[20]也是通过高效液相色谱获得了iturin A2,但是他们采用梯度洗脱,流动相采用0.1%三氟乙酸的乙腈:0.1%三氟乙酸的超纯水,25 min内由30∶70逐渐过渡为55∶45,且在18.5 min左右才能开始收集iturin A2。而本研究是通过等度洗脱,在图1中,iturin A2在8.4 min即可收集,说明此条件可以更加快速、高效地获得纯品。
飞行时间质谱技术是近年发展较快的一门技术,它在生命科学研究领域得到了广泛的应用,尤其是在蛋白质的鉴定方面,不少研究者[20-22]通过飞行时间质谱技术对肽类物质进行鉴定,并获知了该物质的结构。本研究亦利用此技术将巨大芽胞杆菌B196菌株分泌的拮抗物质进行鉴定,并确定其一级结构为Pro-Asn-Ser-βAA-Asn-Tyr-Asn-Gln。

Iturin A2能够抑制立枯丝核菌(R.solani)[16]、玉米小斑病菌(B.maydis)[13]和小麦赤霉病菌(Fusariumgraminearum)[19]等植物病原真菌的生长,也能够抑制水稻白叶枯病菌(Xanthomonasoryzaepv.oryzae)和黄瓜细菌性角斑病菌(Pseudomonassyringaepv.lachrymans)[25]等植物病原细菌的生长。同时,iturin A2在田间可有效地防治玉米小斑病[13]。在本研究中,iturin A2虽然对卵菌的抑制作用相对较差,但是对植物病原菌普遍都具有较高活性,这一点跟陈华等[19]的研究结果相似。可见,巨大芽胞杆菌(B.megaterium)B196菌株分泌的iturin A2对植物病害具有较好的生防潜力,具有开发成防治植物病害生物型农药的前景。
据报道,巨大芽胞杆菌作为生防菌,可防治南方根结线虫[Meloidogyneincognita(Kofoid and White) Chitwood][26]、小麦全蚀病[Gaeumannomycesgraminisvar.tritici(Sacc.) Walker]、小麦纹枯病(Rhizoctoniacerealisvan der Hoeven)、棉花立枯病(RhizoctoniasolaniKühn)和棉花枯萎病[Fusariumoxysporumf.sp.vasinfectum(Atk.)Snyder et Hansen][27]等植物病害,但普遍存在着对植物病害防治效果低、防效单一、防效易受菌体在土壤中定殖能力的影响等缺点[27]。本研究已确定巨大芽胞杆菌(B.megaterium)B196菌株分泌出的抗菌物质是iturin A2,但在田间对植物病害的防效如何,有待试验证实。
[1] 刘月英,傅锦坤,陈平,等.巨大芽胞杆菌D01吸附金(Au3+)的研究[J].微生物学报,2000,40(4): 425-429.
[2] 王琳,李季,张鹏岩.巨大芽胞杆菌对富营养化景观水体的净化效果[J].生态环境学报,2009,18(1): 75-78.
[3] 周军英,林玉锁,徐亦刚,等.巨大芽胞杆菌LY-4对土壤中杀虫单农药的降解[J].中国环境科学,2000,20(6): 511-514.
[4] 郑传进,黄林,龚明.巨大芽胞杆菌解磷能力的研究[J].江西农业大学学报(自然科学版),2002,24(2): 190-192.
[5] 曹燕鲁,吴琼,刘矫.巨大芽胞杆菌B1301分泌物的抑菌作用及其特性的研究[J].山东轻工业学院学报,2004,18(2): 51-55.
[6] 张鑫,王婷,崔灵芝,等.苔藓内生菌Bacillusmegateriumz12的分离、鉴定和抗菌活性筛选[J].中国抗生素杂志,2012,37(6): 401-405,420.
[7] 杨合同,唐文华,迟建国,等.植病生防菌株B1301的种类鉴定及其对生姜青枯病的作用机理和防治效果[J].中国生物防治,2002,18(1): 21-23.
[8] Ko W H,Hora F K,Herlicska E.Isolation and identification of a volatile fungistatic substance from alkaline soil[J].Phytopathology,1974,64: 1398-1400.
[9] Khalil R,Elbahloul Y,Djadouni F,et al.Isolation and partial characterization of a bacteriocin produced by a newly isolatedBacillusmegaterium19 strain[J].Pakistan Journal of Nutrition,2009,8(3): 242-250.
[10]Pueyo M T,Jr C B,Carmona-Carlos A M,et al.Lipopeptides produced by a soilBacillusmegateriumstrain[J].Microbial Ecology,2009,57: 367-378.
[11]农倩,陈雪凤,黎起秦,等.水稻内生细菌B196的鉴定及其对水稻纹枯病的防治作用[J].中国生物防治学报,2011,27(1): 99-103.
[12]秦健,袁高庆,王雅,等.巨大芽孢杆菌B196菌株发酵滤液的抑菌谱及其稳定性[J].西南农业学报,2012,25(5): 1694-1697.
[13]Ye Yunfeng,Li Qiqin,Fu Gang,et al.Identification of antifungal substance (iturin A2)produced byBacillussubtilisB47 and its effect on southern corn leaf blight[J].Journal of Integrative Agriculture,2012,11(1): 90-99.
[14]Isogai A,Takayama S,Murakoshi S,et al.Structure of β-amino acids in antibiotics iturin A[J].Tetrahedron Letters,1982,23: 3065-3068.
[15]Hiradate S,Yoshida S,Sugie H,et al.Mulberry anthracnose antagonists (iturins)produced byBacillusamyloliquefaciensRC-2[J].Phytochemistry,2002,61: 693-698.
[16]Yu G Y,Sinclair J B,Hartman G L,et al.Production of iturin A byBacillusamyloliquefacienssuppressingRhizoctoniasolani[J].Soil Biology & Biochemistry,2002,34: 955-963.
[17]Roeptorff P,Fohlman J.Proposal for a common nomenclature for sequence ions in mass spectra of peptides[J].Biomedical Mass Spectrometry,1984,11(11): 601.
[18]Biemann K.Mass spectrometry of peptides and proteins[J].Annual Review of Biochemistry,1992,61: 977-1010.
[19]江木兰,王国平,胡小加,等.油菜内生菌BY-2的脂肽类抑菌物质的分子结构鉴定[J].中国油料作物学报,2010,32(2): 279-284.
[20]陈华,袁成凌,蔡克周,等.枯草芽孢杆菌JA 产生的脂肽类抗生素-Iturin A 的纯化及电喷雾质谱鉴定[J].微生物学报,2008,48(1): 116-120.
[21]颜再生.本土液化淀粉芽胞杆菌含伊枯草菌素A同分异构体[D].朝阳:朝阳科技大学,2007.
[22]Cho S J,Lee S K,Cha B J,et al.Detection and characterization of theGloeosporiumgloeosporioidesgrowth inhibitory compound iturin A fromBacillussubtilisstrain KS03[J].FEMS Microbiology Letters,2003,223: 47-51.
[23]Gong M,Wang J D,Zhang J,et al.Study of the antifungal ability ofBacillussubtilisstrain PY-1in vitro and identification of its antifungal substance (Iturin A)[J].Acta Biochimica et Biophysica Sinica,2006,38(4): 233-240.
[24]Arrebola E,Jacobs R,Korsten L.Iturin A is the principal inhibitor in the biocontrol activity ofBacillusamyloliquefaciensPPCB004 against postharvest fungal pathogens[J].Journal of Applied Microbiology,2010,108: 386-395.
[25]Phae C G,Shoda M,Kubota H.Suppressive effect ofBacillussubtilisand it’s products on phytopathogenic microorganisms[J].Journal of Fermentation and Bioengineering,1990,69(1): 1-7.
[26]梁建根,郑经武,郝中娜,等.生防菌K-8对南方根结线虫的防治及其鉴定[J].中国农学通报,2011,27(21): 282-286.
[27]杨合同,陈凯,李纪顺,等.重组巨大芽孢杆菌在小麦根际的定殖及其对植物真菌病害的防治效果[J].山东科学,2003,16(3): 12-17.
IsolationandpurificationofanantimicrobialsubstanceproducedbyBacillusmegateriumstrainB196
Liao Ting, Qin Jian, Yuan Gaoqing, Li Qiqin, Lin Wei, Peng Haowen
(CollegeofAgriculture,GuangxiUniversity,Nanning530005,China)
An antimicrobial substance produced byBacillusmegateriumstrain B196can strongly inhibit the growth ofRhizoctoniasolani.Identification of the antimicrobial substance is very important for further research on the inhibition mechanism and its application.Crude antimicrobial substances were extracted with methanol from the precipitate which was obtained by adding HCl to the cell-free culture broth and then stored at 4 ℃ overnight.The crude extract was further purified by reversed-phase HPLC with a C18 column(PRC-ODS,20×250nm)to separate the substances.An antifungal compound with strong inhibitory activity againstR.solaniwas purified.The molecular weight of the compound was 1042.592 7 u by Matrix-assisted laser desorption ionization time-of-flight mass spectrometer(MALDI-TOF-MS).TESI collision induced dissociation mass spectrometry analysis demonstrated that the compound had a primary structure of Pro-Asn-Ser-βAA-Asn-Tyr-Asn-Gln.Therefore,the compound was identified as Iturin A2.
Bacillusmegaterium; iturin A2; isolation and purification
2013-05-31
:2013-09-27
广西自然科学基金项目(2011GXNSFA018064)
S482.292
:ADOI:10.3969/j.issn.0529-1542.2014.02.004
* 通信作者 E-mail: qqli5806@gxu.edu.cn
猜你喜欢
中国生物防治学报(2022年3期)2022-07-09
科学与财富(2021年33期)2021-05-10
微生物学杂志(2020年2期)2020-12-31
农民致富之友(2019年21期)2019-08-08
中国酿造(2019年5期)2019-06-11
特别健康(2018年3期)2018-07-04
小猕猴智力画刊(2017年12期)2017-12-27
儿童故事画报·智力大王(2016年5期)2016-07-04
第二课堂(课外活动版)(2015年5期)2015-10-21
