
蝴蝶兰倍性与叶部和花部性状的相关性
2014-08-31 07:17易懋升曾瑞珍黎杨辉张志胜
华南农业大学学报 2014年5期
谢 利,刘 芳,易懋升,曾瑞珍,夏 晴,黎杨辉,张志胜
(1 华南农业大学,广东省植物分子育种重点实验室,广东 广州 510642;2 广州花卉研究中心,广东 广州 510360)
蝴蝶兰倍性与叶部和花部性状的相关性
谢 利1,刘 芳1,易懋升2,曾瑞珍1,夏 晴2,黎杨辉2,张志胜1
(1 华南农业大学,广东省植物分子育种重点实验室,广东 广州 510642;2 广州花卉研究中心,广东 广州 510360)
【目的】研究蝴蝶兰Phalaenopsis倍性与主要性状的相关性.【方法】采用压片法对85份蝴蝶兰资源的倍性进行了鉴定,测量了这些资源的气孔长度、气孔密度、叶长、叶宽、叶厚、花朵直径和花朵数.【结果和结论】85份蝴蝶兰资源中3份为二倍体,占3.5%;9份为三倍体,占10.6%;73份为四倍体,占85.9%.四倍体蝴蝶兰的气孔长度和花朵直径显著大于二倍体和三倍体,而气孔密度和花朵数显著少于二倍体和三倍体(P<0.05).三倍体蝴蝶兰的气孔长度、花朵直径和花朵数与二倍体蝴蝶兰差异均不显著(P>0.05),但气孔密度显著小于二倍体(P<0.05).四倍体蝴蝶兰的叶长显著大于二倍体(P<0.05),但二倍体与三倍体、三倍体与四倍体在叶长上差异不显著(P>0.05).二倍体、三倍体和四倍体在叶宽和叶厚上差异均不显著(P>0.05).蝴蝶兰倍性与气孔长度、叶长和花朵直径呈极显著的正相关关系,与气孔密度和花朵数呈极显著的负相关关系.
蝴蝶兰; 倍性; 气孔性状; 花部性状
蝴蝶兰Phalaenopsis是兰科植物中栽培最广泛、产业化技术最成熟和最受欢迎的种类之一,具有广阔的市场前景.新品种选育是蝴蝶兰产业的基础和可持续发展的动力,多倍体育种是蝴蝶兰育种的主要方法[1],研究蝴蝶兰资源的倍性及其与形态学性状的相关性对建立蝴蝶兰高效多倍体育种技术体系具有十分重要意义.
有关蝴蝶兰多倍体育种已有一些报道[2- 6].庄东红等[7]对蝴蝶兰属38个品种(系)的染色体数和形态的分析表明,供试蝴蝶兰品种(系)均为多倍体,且不同品种(系)的染色体长度和形态差异较大,染色体大小组成与花色有一定相关性.周建金等[8-9]研究了不同倍性蝴蝶兰杂交种子的萌发及杂交后代的染色体倍性,结果表明,不同倍性蝴蝶兰杂交的种子萌发率不同,除三倍体和三倍体杂交未能产生杂交后代外,利用二倍体、三倍体和四倍体蝴蝶兰杂交可产生二倍体、三倍体、四倍体、五倍体、六倍体、八倍体和非整倍体.Chen等[3]发现菲律宾白花蝴蝶兰P.aphrodite和巴拉旺蝴蝶兰P.hieroglyphica四倍体的气孔密度显著小于二倍体,且四倍体的花朵更大花瓣更厚.但对商业化蝴蝶兰品种倍性与其植物学和观赏性状之间相关性的研究少见报道.本研究在对85份蝴蝶兰资源进行倍性鉴定的基础上,分析了倍性与叶部和花部性状之间的相关性,为蝴蝶兰高效多倍体育种技术体系的建立提供指导.
1 材料与方法
1.1 材料
供试材料为2个蝴蝶兰原生种和83个栽培品种,其中包括31个朵丽蝶兰Doritaenopsis品种,选择种植2年左右的开花植株.所有材料种植于广州花卉研究中心花卉种质资源圃中,常规方法管理.
1.2 方法
1.2.1 染色体倍性鉴定 参考张志胜等[10]的压片法,根据蝴蝶兰根尖的特点进行了改进.具体方法如下:切取蝴蝶兰植株幼嫩的根尖2~3 mm;洗净后,于15 ℃黑暗条件下,在0.002 mol·L-1的8-羟基喹啉中浸泡6~7 h;清水洗净后,用新配制的卡诺氏液[V(乙醇)∶V(冰醋酸)=3∶1,其中乙醇的体积分数为95%]在4 ℃冰箱内固定12~24 h,然后用1 mol·L-1的HCl在60 ℃恒温水浴锅中酸解3~4 min,蒸馏水洗净;切取根尖的分生组织放在预先滴1滴卡诺氏液的载玻片上,用镊子压碎,弃去残渣,快速经过酒精灯的火焰2~3次;滴1滴石碳酸品红进行染色,染料快干时盖上盖玻片,用镊子轻压几下,快速经过火焰6~7次.在Olympus-IX71倒置显微镜下观察,选取染色体分散好的细胞拍照,统计染色体数目.
1.2.2 气孔鉴定 气孔长度和密度的测量参考张秀芳等[11]的印迹法.具体方法为:用蒸馏水擦洗健康成熟叶片下表面,干燥后,在叶片下表面的中部约1 cm×3 cm的面积上均匀涂上一层透明指甲油(Maypelline),待指甲油干燥后,将指甲油膜取下.加1滴蒸馏水在载玻片上,将与叶片接触的一面向上置于载玻片有水的地方,用吸水纸在上面压一下,使膜平整,然后在Motic-B1显微镜下观察,随机取10个视野拍照,统计气孔数,每个视野随机取1个气孔,并用标定好的测微尺测量其长度,重复3次.气孔密度的计算公式如下:
气孔密度(个/mm2)=各视野中的气孔数/ 视野的真实面积.
1.2.3 叶片长度、宽度和厚度的测量 叶片基部离层至叶尖的距离为叶长, 叶片最大宽幅为叶宽,叶长和叶宽用直尺测量.叶厚是每片叶中部靠近叶脉处的厚度,用游标卡尺测量.每株测量2片健康成熟的叶片,分别是从顶部倒数的第2和第3片叶,每品种测量10株,5株为1个重复.
1.2.4 花朵数和花朵直径的鉴定 参照国际植物新品种保护联盟(UPOV)制定的蝴蝶兰属新品种DUS测试指南中的赋值标准对花朵数和花朵直径进行鉴定.每品种统计10株健康植株的花朵数,并测量每株第1~3朵花的花朵直径.
1.3 统计分析
利用SPSS 13.0对数据进行统计分析.
2 结果与分析
2.1 蝴蝶兰资源的倍性鉴定
经染色体数目分析发现,在供试的85份材料中,有3个为二倍体,2n=2x=38,占3.5%;9个为三倍体,2n=3x=57,占10.6%;73个为四倍体,2n=4x=76,占85.9%(表1,图1).其中,2个原生种均为二倍体;商业化的蝴蝶兰栽培品种中除P.‘Timothy Christopher’外,其他均为多倍体.
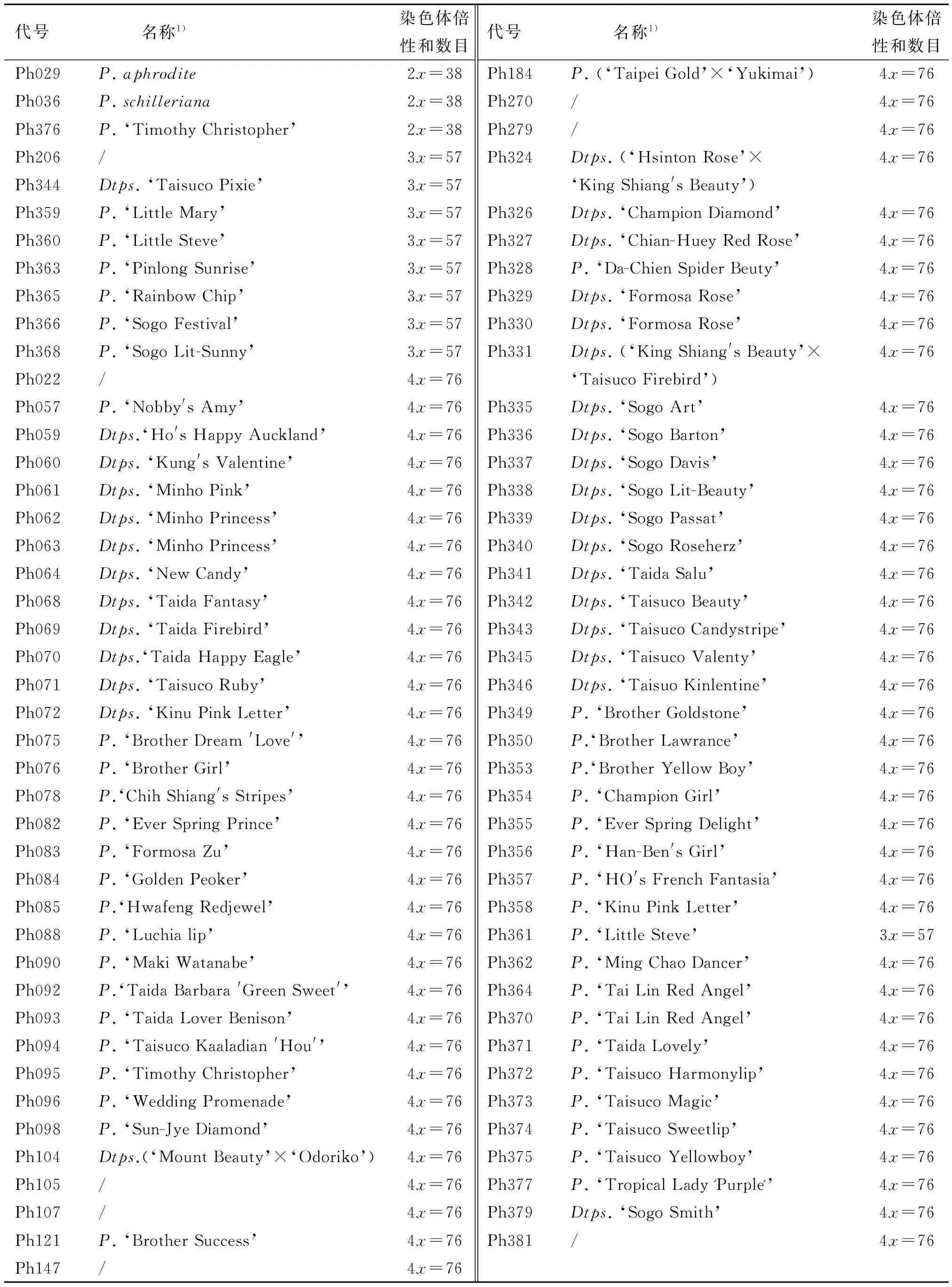
表1 蝴蝶兰资源的根尖染色体倍性和数目Tab.1 The chromosome ploidy level and the number of root tips of tested Phalaenopsis
1) 该列名称为英国皇家学会登录的品种名;/ 表示材料的名称尚未明确.

a:二倍体,P. aphrodite,2n=2x=38;b:三倍体,P. ‘Little Mary’,2n=3x=57;c:四倍体,P. ‘Ever Spring Prince’,2n=4x=76.
2.2 倍性与叶片性状的相关性
2.2.1 倍性与气孔性状的相关性 不同倍性蝴蝶兰的气孔长度和气孔密度均存在一定的变异范围,随着倍性的增加气孔长度逐渐增大,气孔密度逐渐减小.四倍体的气孔长度显著大于二倍体和三倍体,而气孔密度显著小于二倍体和三倍体(P<0.05);三倍体的气孔长度与二倍体差异不显著(P>0.05),但气孔密度显著小于二倍体(P<0.05)(表2,图2).
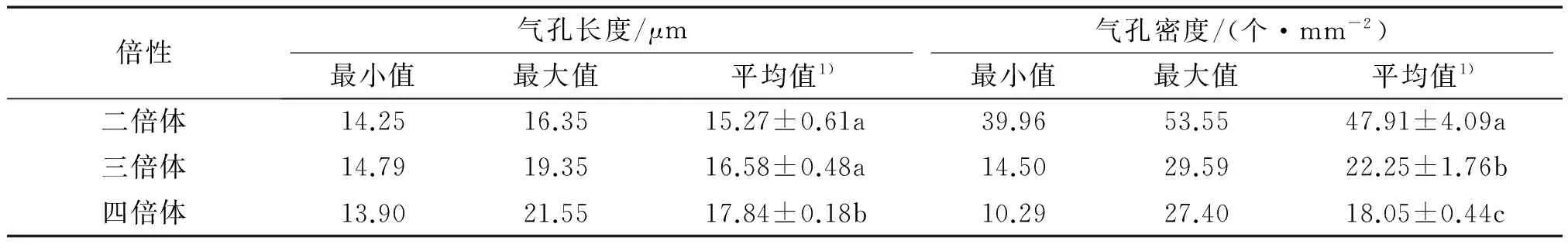
表2 不同倍性蝴蝶兰的气孔长度和气孔密度Tab.2 The length and density of stomatal cell of different ploidy Phalaenopsis
1)该列数据为2次重复的平均值±标准误,该列数据后凡是有一个相同小写字母者,表示差异不显著(Duncan’s法,P>0.05).

a、b: 二倍体P. aphrodite;c、d: 三倍体P. ‘Little Mary’;e、f: 四倍体,P. ‘Taida Lover Benison’.
对蝴蝶兰的倍性与气孔长度和气孔密度进行了相关性分析.结果表明,蝴蝶兰的气孔长度与倍性的相关系数为0.372(t=3.65>t(0.01,84)=2.64),呈极显著的正相关关系;而气孔密度与倍性的相关系数为-0.719(t=9.42>t(0.01,84)=2.64),呈极显著的负相关关系.2.2.2 倍性与叶长、叶宽和叶厚的相关性 不同倍性蝴蝶兰的叶长、叶宽和叶厚均存在一定的变异范围,随着倍性的增加,蝴蝶兰叶片变长变宽(表3).四倍体的叶长显著大于二倍体,但二倍体与三倍体、三倍体与四倍体在叶长上差异不显著(P>0.05);二倍体、三倍体和四倍体在叶宽和叶厚上均差异不显著(P>0.05).
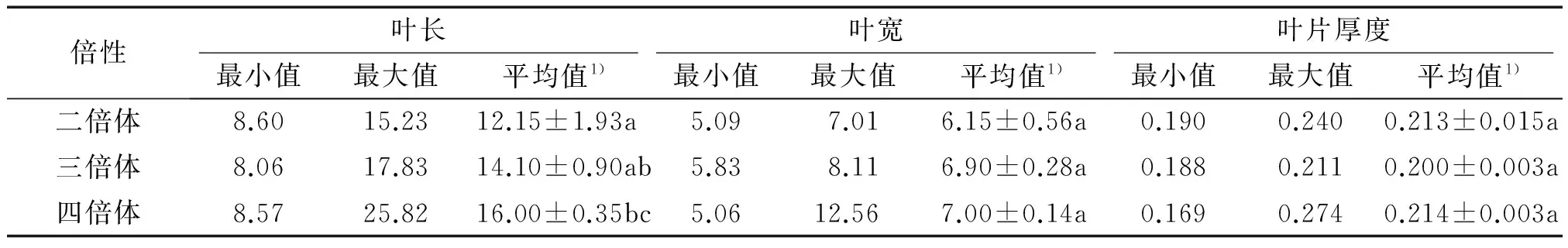
表3 不同倍性蝴蝶兰的叶片长度、宽度和厚度Tab.3 The length, width and thickness of leaf of different ploidy Phalaenopsis cm
1)该列数据为2次重复的平均值±标准误,该列数据后凡是有一个相同小写字母者,表示差异不显著(Duncan’s法,P>0.05).
对蝴蝶兰倍性与叶长、叶宽和叶厚进行了相关性分析,叶长与倍性的相关系数为0.292(t=2.78>t(0.01,84)=2.64),呈极显著的正相关,但叶宽和叶厚与倍性的相关关系不显著.
2.3 倍性与花部性状的相关性
不同倍性蝴蝶兰的花朵直径和花朵数存在一定的变异范围(表4).随着倍性的增加花朵直径逐渐增大,四倍体的花朵直径显著大于二倍体和三倍体(P<0.05),但二倍体和三倍体的花朵直径差异不显著(P>0.05).三倍体蝴蝶兰的花朵数最多,四倍体的花朵数显著少于二倍体和三倍体(P<0.05),但二倍体和三倍体的花朵数差异不显著(P>0.05).
对蝴蝶兰倍性与花朵直径和花朵数进行了相关性分析,相关系数分别为0.552(t=6.03>t(0.01,84)=2.64)和-0.509(t=5.39>t(0.01,84)=2.64),表明蝴蝶兰倍性与花朵直径呈极显著的正相关,而与花朵数呈极显著的负相关.
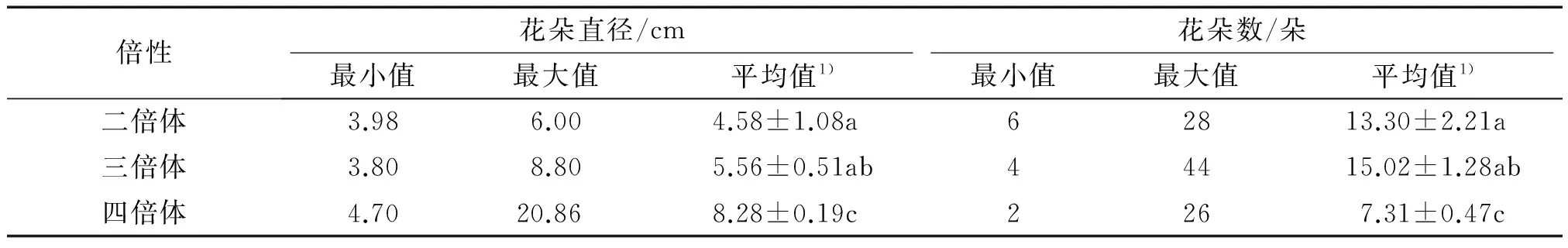
表4 不同倍性蝴蝶兰的花朵直径和花朵数
1)该列数据为2次重复的平均值±标准误,该列数据后凡是有一个相同小写字母者,表示差异不显著(Duncan’s法,P>0.05).
3 讨论与结论
多倍体具有营养体巨大、叶片宽厚和花大等特点[12].本研究结果表明,随着倍性的增加,蝴蝶兰叶片逐渐变大,四倍体的叶长显著大于二倍体;花朵直径逐渐增大,四倍体的花朵直径显著大于三倍体和二倍体,但花朵数量显著少于二倍体和三倍体,三倍体花朵数最多.因此在进行蝴蝶兰育种时,如果以大花为目标,应选育四倍体;如果以小花多花为目标,可培育二倍体;如果以小花多花或中花多花为目标,可选育三倍体.
倍性鉴定是倍性育种的重要环节.因此,寻找一种简单有效的倍性鉴定方法,对于提高蝴蝶兰倍性育种效率至关重要.Stebbins[13]认为在不便直接测定染色体数目时,单用测量植物体的某些细胞如气孔保卫细胞的方法,就能估计其为二倍体还是多倍体.刘奕清等[14]、刘静等[15]、廖道龙等[16]、庄东红等[17]研究发现多倍体气孔明显比二倍体大,气孔密度明显比二倍体小.张志胜等[10]对红掌Anthuriumandraeanum多倍体的研究表明,通过气孔长度和密度鉴定红掌四倍体是可靠的.利用气孔性状进行倍性鉴定操作简单、易行,已在许多植物中得到了应用[18- 21].本研究表明,随着蝴蝶兰倍性的增加,气孔长度呈增加趋势,气孔密度呈减少趋势,气孔长度与倍性呈极显著的正相关关系,而气孔密度与倍性呈极显著的负相关关系,这与Chen等[3]的研究结果一致.但因同倍体蝴蝶兰的气孔长度和密度也存在一定的变异范围,使不同倍性间的气孔长度和密度存在重叠的可能,因此在进行蝴蝶兰多倍体育种时,可先通过气孔长度和密度进行初步筛选,然后再用压片法进行准确鉴定.
不同倍性蝴蝶兰杂交亲和性、种子萌发率和后代倍性多样性不同[8-9],因此,为了提高杂交育种的效率,可将染色体的倍性做为亲本选择的依据之一.本研究通过采用根尖压片法对85份供试蝴蝶兰资源的倍性鉴定表明,现有蝴蝶兰栽培品种多为三倍体和四倍体,这为更好利用这些资源开展蝴蝶兰育种提供了依据.
[1] TANG Chingyan, CHEN WenHuei. Breeding and development of new varieties inPhalaenopsis[M]∥CHEN Wenhuei,CHEN Honghwa. Orchid biotechnology. Singapore:World Scientific Publishing Co.Pte.Ltd.,2007: 1- 22.
[2] GRIESBACH R J. Polypioidy inPhalaenopsisorchid improvement[J]. J Hered, 1985, 76(1): 74-75.
[3] CHEN Wenhuei, TANG Chingyan, KAO Yulin. Ploidy doubling by in vitro culture of excised protocorms or protocorm-like bodies inPhalaenopsisspecies[J]. Plant Cell Tiss Organ Cult, 2009, 98(2): 229- 238.
[4] CHEN Wenhuei, TANG Chingyan, LIN Tsaiyun, et al. Changes in the endopolyploidy pattern of different tissues in diploid and tetraploidPhalaenopsisaphroditesubsp.formosana(Orchidaceae) [J]. Plant Sci, 2011, 181(1): 31-38.
[5] MIGUEL T P, LEONHARDT K W.Invitropolyploid induction of orchids using oryzalin[J]. Sci Hortic, 2011, 130(1): 314-319.
[6] 崔广荣,张子学,胡能兵,等. 蝴蝶兰类原球茎液体培养中用秋水仙素诱导多倍体[J]. 浙江大学学报:农业与生命科学版,2010,36(1):49-55.
[7] 庄东红,曲莹,许大熊,等.蝴蝶兰若干品种(系)的染色体数和形态分析[J].园艺学报,2007,34(5):1257-1262.
[8] 周建金,谢利,黎杨辉,等.不同倍性蝴蝶兰杂交种子的离体培养[J].种子,2009,28(7):45- 49.
[9] 周建金,曾瑞珍,刘芳,等.不同倍性蝴蝶兰杂交后代的染色体倍性研究[J].园艺学报,2009,36(10):1491-1497.
[10]张志胜,黎杨辉,姜蕾,等.红掌四倍体的离体诱导及其鉴定[J]. 园艺学报,2007,34 (3):729-734.
[11]张秀芳,石东里,张 兰.观察植物气孔结构的简易方法[J].生物学通报,2002,37 (6):42.
[12]雷家军,王冲.观赏植物多倍体诱导研究进展[J].东北农业大学学报,2012,43(1):18- 24.
[13]STEBBINS G L. Variation and evolution in plants [M]. New York: Columbia University Press, 1950.
[14]刘奕清,吴林,陈泽雄,等.秋水仙素离体诱导金银花多倍体及倍性鉴定[J].中药材,2012,35(11):1729-1732.
[15]刘静,赵庆芳,丁兰.兰州百合多倍体诱导及鉴定[J].北方园艺,2011,(18):141-143.
[16]廖道龙,谢利,曾瑞珍,等.石斛属植物倍性与形态学性状的相关性研究[J].西北植物学报,2012,32(10):2023- 2029.
[17]庄东红,宋娟娟.木槿属植物染色体倍性与花粉粒、叶片气孔器性状的关系[J].热带亚热带植物学报,2005,13 (1):49-52.
[18]AHMED N,SINGH I D. A technique for rapid identification of ploidy levels in tea[J]. Two and a Bud, 1993,40(2):31-33.
[19]李赟,石荫坪,束怀瑞.应用气孔性状对苹果与梨的倍性判别分析[J].果树科学,1999,16(1):9-13.
[20]张晓芬,耿三省,陈斌.辣椒染色体倍性与气孔性状的相关性[J].江苏农业学报,2009,25(2):339-342.
[21]HO I, WAN Y, WIDHOLM J M, et al. The use of stomatal chloroplast number for rapid determination of ploidy level in maize[J]. Plant Breed, 1990, 105(3): 203- 210.
【责任编辑李晓卉】
AcorrelationbetweenploidylevelsandcharactersofleafandflowerinPhalaenopsis
XIE Li1, LIU Fang1, YI Maosheng2, ZENG Ruizhen1, XIA Qing2, LI Yanghui2, ZHANG Zhisheng1
(1 Guangdong Provincial Key Laboratory of Plant Molecular Breeding, South China Agricultural University, Guangzhou 510642, China; 2 Guangzhou Flower Research Center, Guangzhou 510360, China)
【Objective】 To research the correlation between ploidy levels and main characters inPhalaenopsis. 【Method】 Ploidy levels of 85Phalaenopsisaccessions were identified by using squash method, and the length and density of stoma, the length, width and thickness of leaf, the diameter and the number of flowers were also measured. 【Result and conclusion】The results showed that in the 85 accessions there were three diploids (3.5%), nine triploids (10.6%) and 73 tetraploids (85.9%). Stomatal lengths and the length of tetraploid flower were obviously longer than those of diploid and triploid flower, while the density of stoma and the number of flowers were fewer than that of diploid and triploid(P<0.05). The differences of the stomatal cell length and diameter and the number of flower between diploid and triploid were not significant at 5% level, however, the stomatal density of triploid was less than that of diploid(P<0.05). The leaf length of tetraploid was significantly longer than that of diploid(P<0.05), while the difference of leaf length between diploid and triploid or between triploid and tetraploid was not significant(P>0.05). The difference of leaf width and thickness among diploid, triploid and teraploid were also not significant either(P>0.05).The ploidy levels were, to a large extant, positively associated to stomata length, leaf length and flower diameter, but negatively correlated with stomatal density and the number of flowers.
Phalaenopsis; ploidy level; stomatal character; flower character
2013- 08- 19优先出版时间2014- 07- 17
优先出版网址:http:∥www.cnki.net/kcms/detail/44.1110.S.20140717.0909.024.html
谢 利(1974—),女,讲师,博士,E-mail:xieli@scau.edu.cn;通信作者:张志胜(1965—),男,教授,博士,E-mail:zszhang@scau.edu.cn
广州市农业局2004专项资金项目“蝴蝶兰优良新品种的引种与选育”;广州科技局农业攻关项目“利用远缘杂交育种方法培育蝴蝶兰新品种”
谢 利,刘 芳,易懋升,等.蝴蝶兰倍性与叶部和花部性状的相关性[J].华南农业大学学报,2014,35(5):82- 87.
Q343.2; S682.31
A
1001- 411X(2014)05- 0082- 06
猜你喜欢
中国海洋大学学报(自然科学版)(2023年2期)2023-02-21
科学养鱼(2021年11期)2022-01-11
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
新疆农业科学(2020年1期)2020-02-14
亚热带植物科学(2019年2期)2019-07-22
食品与生活(2019年12期)2019-05-15
江苏农业学报(2018年5期)2018-09-10
生命科学研究(2018年1期)2018-05-29
临床与实验病理学杂志(2017年3期)2017-03-07
