
增龄与骨骼肌肌细胞的退变
2014-09-12 09:22曹玲杨光
中国老年学杂志 2014年10期
曹 玲 杨 光
(大连理工大学体育教学部,辽宁 大连 116024)
老年人随着年龄的增长,各个器官的功能逐渐衰退,从而影响其健康和日常生活。其中,骨骼肌衰老是人体在增龄过程中的必然现象,会带来更为严重的后果,比如,易导致老年人跌倒〔1〕、骨折以及各种骨骼疾病,由此造成的经济损失每年超过180亿美元〔2〕。由此可见,骨骼肌衰老不仅影响了老年人的身心健康和生活质量,也增加了家庭和社会的经济负担。增龄过程中,骨骼肌在结构组成上发生了变化,全面研究并充分认识这一变化,对预防与干预骨骼肌衰老,提高老年人的日常生活能力具有十分重要的意义。本文从细胞和分子水平上,对国内外骨骼肌肌细胞退变的相关文献进行综述,希望能为该领域的研究提供参考,并为预防与干预老年人骨骼肌功能衰退提供理论依据。
1 肌力下降
早在1989年,就有研究者提出了“肌肉减少症(sarcopenia)”这一概念〔3〕,最初是指伴随增龄的“肌肉含量的减少”。然而,近年来美国老年人研究所和欧洲肌肉减少症工作组将肌肉减少症定义为“肌肉含量的减少和肌肉力量的降低”〔4〕。在这一概念变化的基础上,用“肌肉含量的减少”就不能解释增龄带来的骨骼肌衰老的现象〔5〕。增龄过程中骨骼肌的衰老主要表现为肌肉力量(肌力)的下降。导致肌力下降的主要因素包括肌肉含量(肌肉重量和横截面积)的减少和相对肌力(肌力除以肌肉横截面积)的降低。
根据有关活动量减少〔6~10〕、老龄化〔6,9,10,11~15〕问题的动物与人体研究发现,与活动量减少的群体相比,老龄组相对肌力降低的趋势比肌肉含量减少的趋势显著。活动量减少导致的肌力降低主要与肌肉含量的变化有关;然而也有大量的研究表明,肌力的降低与相对肌力的下降有关。另外,在老龄组中,与肌肉含量的变化相比,相对肌力的降低率具有更显著的变化趋势。因此,笔者认为相对肌力的下降可能是肌力下降的主要原因。
2 增龄与骨骼肌组成的变化
老年人的骨骼肌随着年龄的增加,非收缩肌肉组织有所增加〔16〕。因此,伴随增龄,在非收缩组织含量增加的情况下,收缩蛋白质的比例降低,可以认为骨骼肌的相对肌力下降。事实上,通过计算机辅助截面摄影,根据图像中色彩的强度可以分析骨骼肌的组成成分,脂肪细胞量的增加导致相对肌力的降低,被称为“肌肉脂肪症”(myosteatosis)〔17〕。另根据核磁共振成像法对骨骼肌的测试结果发现,老年人的脂肪细胞量与年轻人相比增加了2倍以上〔18〕,也就是说,在增龄过程中,非收缩性组织的增加是骨骼肌衰老的一个主要因素,肌纤维内的脂肪沉积则是由于老化的骨骼肌细胞氧化代谢能力下降所致。这一观点也被Payne等〔19〕在大龄鼠的比目鱼肌和趾长伸肌的实验中证实。该研究还指出伴随着肌纤维间的脂肪细胞浸润,细胞间隙扩大,相对肌力逐渐下降。
3 增龄与肌细胞的变化
肌肉的收缩是基于多个复杂过程的联动而发生的(图1)。兴奋-收缩耦联是肌肉的重要功能特点之一,图1(i)~(iv)的任何一个过程出现机能障碍,无论肌肉含量是否减少,都会导致肌肉相对肌力的降低。
(i)细胞膜内产生的活动电位,引起存在于横小管的电位依赖性Ca2+通道(DHPR)的立体结构变化;(ii)DHPR和肌质网Ca2+通道(RyR)在功能上协同导致细胞内Ca2+浓度上升;(iii)Ca2+与肌钙蛋白结合;(iv)被称为横桥的肌动蛋白与肌球蛋白的相互作用。
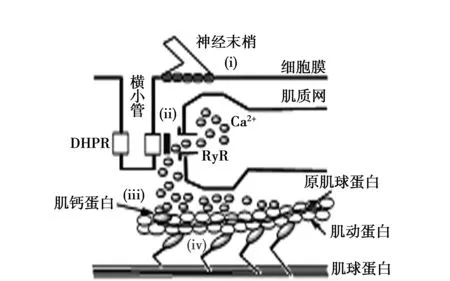
图1 兴奋-收缩耦联
3.1肌质网释放Ca2+量的降低 Delbono等〔20〕从股外肌取出肌纤维,通过去极化诱导细胞内Ca2+浓度上升,结果首次发现老年人肌细胞内的Ca2+浓度上升幅度比年轻人的要低。其他研究者在动物实验中,以小鼠作为研究对象,均得到了相同的结果〔21,22〕。由此可以证实,随着机体的老化,从细胞膜到肌质网的Ca2+通道- ryanodine 受体(RyR)之间出现了机能不健全的状况,其中 RyR则可以通过Ca2+诱导Ca2+释放机制将肌质网中的Ca2+释放于细胞质中,细胞质的浓度可以增强通道蛋白的活动,但通道的打开受到横管系统的双氢吡啶受体(dihydropyridine acceptor,DHPR)的调节,所以,DHPR与RyR在机制上是相互联系的,在增龄的过程中,这种联系在机能上可能存在联动障碍。
由于RyR是同源四聚体,1分子的RyR与 4分子的DHPR对应形成复合体,对于骨骼肌的兴奋-收缩耦联,去极化的电子信号转化为Ca2+的效率,依赖于这些可直接相互作用的复合体的量。已有研究发现,伴随老化,DHPR的1s亚单元的量在大鼠〔23〕、小鼠〔24〕以及兔子〔25〕的体内均有所减少。也有研究发现,大龄大鼠与小龄大鼠相比,联动的DHPR-RyR复合体的量,快肌约减少30%、慢肌约减40%〔23〕。因此可以认为,在增龄过程中,兴奋-伸缩耦联的信息传送效率显著降低,这就导致了骨骼肌功能的衰退。另外,形态学方面的研究结果也支持这一观点,通过电子显微镜能观察到老年人的骨骼肌横行小管和肌质网的末端组成的复合体数目减少的现象〔26〕。然而,对于大鼠〔27〕和人〔28〕的实验也有结果表明,在增龄的过程中,DHPR量的变化不受影响。之所以有不一致的观点,其原因目前尚不清楚,笔者认为与研究对象、所研究的肌肉种类、年龄等不同有关,还需要进一步的深入研究来证实。
3.2肌原纤维的收缩力降低 通过物理或化学的方法将细胞膜剥离后的单一肌纤维暴露于Ca2+溶液中得到的张力不受肌质网的Ca2+放出能力的影响,反映了肌原纤维的选择性机能。到目前为止,从人〔14,29〕和兔子〔6〕体内取出的单一肌纤维中Ca2+诱导的最大收缩力测定的数据表明,相对肌力伴随增龄而降低。也就是说,可以认为肌原纤维的机能不健全与骨骼肌衰老有关。
可以把肌原纤维称为肌肉的收缩装置,它由粗肌丝(filament)和细肌丝弯曲重叠而成。粗肌丝的主要构成体肌球蛋白头含有ATP分解酶(mATPase),在肌肉收缩时将产生能量。另一方面,细肌丝主要是由肌动蛋白分子螺旋状重叠在一起而构成。肌球蛋白头与肌动蛋白分别存在相互作用的部位,这些相互作用部位利用由ATP分解而产生的能量形成横桥,从而细肌丝向粗肌丝的中心部位滑动。在此过程中,肌球蛋白头和肌动蛋白按照被称为横桥循环的(图2)过程重复进行,形成肌肉收缩。肌原纤维的收缩力是由被调动的横桥数目以及相对于每一个横桥的张力来决定的〔30〕。
(i)在ATP分解酶的作用下,ATP分解为ADP和无机磷(P),肌球蛋白和肌动蛋白弱结合;(ii)无机磷解离后,肌球蛋白和肌动蛋白强结合;(iii)ADP解离后,肌球蛋白从肌动蛋白上脱离。
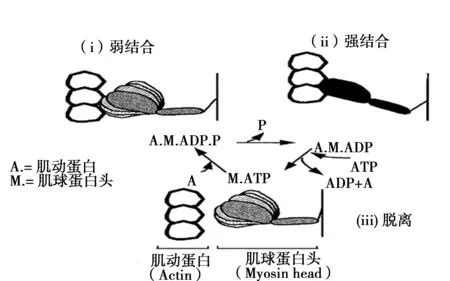
图2 横桥循环〔31〕
肌球蛋白分子是有两条肌球蛋白重链(Myosin heavy chain:MyHC)和4条肌球蛋白轻链所构成。D′Antona等〔32〕从老年人的股外肌中取出单一肌纤维,发现相对肌力与MyHC量之间的相关关系,且MyHC的减少引起被动员横桥的数目降低,从而导致骨骼肌衰老。随后的研究也支持了这一观点,如Thompson等〔33〕指出MyHC 量的降低在老龄大鼠的半膜肌中也观察得到,在增龄的过程中,快肌MyHC的合成量会有所降低〔34〕。还有报道指出,对于被统称为恶疾(cachexia)的心不全、癌症、艾滋病等伴随肌肉含量的降低,是炎症性细胞活素(Cytokine) 作为上调因子的泛激素(ubiquitin)-蛋白酶体(proteasome)复合体系与MyHC的分解相关〔35,36〕。在老年人群中也如此,血液中炎症性细胞活素的浓度与肌肉含量及其肌力间存在相关关系,这一点已得到认同〔37〕。至于在这些关系中存在怎样的因果联系,有待进一步深入研究。另一方面,对于老龄小鼠的比目鱼肌,无论MyHC的发现量是否存在变化,关于相对肌力的低下已有研究报道,仅仅以横桥量的变化很难全面说明肌原纤维机能低下的现象。
电子自旋共振(EPR)法在有关肌球蛋白分子构造变化的研究中得到了广泛应用。肌球蛋白头的催化剂部位存在高反应性的硫氢(SH)基团,通过EPR法进行分析,可以判别肌球蛋白头结合状态的强弱〔38〕。Lowe等〔31〕对大鼠半膜肌的单一肌纤维进行EPR法肌肉等长收缩研究,发现在肌球蛋白头强结合状态下老龄大鼠的肌球蛋白头的比例与小龄大鼠相比降低了约30%。而且,该研究组的研究结果还表明,肌球蛋白中游离SH基团的量随着年龄的增大而降低,活性氧物种直接影响肌球蛋白头机能性障碍。在由大鼠的半腱肌的肌球蛋白中,也发现老龄化伴随的游离SH基团的减少〔15〕,可认为因氧化而导致肌球蛋白结构的变化可能会导致肌原纤维机能的低下。因此,伴随着老化,肌球蛋白的退行性变化引起被动员横桥的数量以及每个横桥的张力降低,从而导致肌原纤维的功能低下。
综上所述,在增龄过程中,骨骼肌衰老主要表现为肌力下降,这种衰退有多种因素,本文从微观角度揭示了骨骼肌肌细胞发生退变的原因:肌质网释放Ca2+量的降低;肌动蛋白和肌球蛋白的横桥数量的减少以及肌球蛋白的结构变化导致的肌原纤维张力的降低。骨骼肌衰老是一个渐进的过程,其机制十分复杂,许多相互联系的因素都会导致骨骼肌衰老发生和发展,全面地研究增龄与骨骼肌衰老的机制,为建立准确诊断、有效预防与干预提供理论依据。
4 参考文献
1曹 玲,佟贵锋,杨 光,等.影响老年人跌倒的多维危险因素〔J〕.中国体育科技,2012;48(3):96-9.
2Rubenstein LZ.Falls in older people:epidemiology,risk factors and strategiges for prevention〔J〕.Age Aging,2006;35:3541.
3Rosenberg IH.Summary comments〔J〕.Am J Clin Nutr,1989;(50):1231-3.
4Cruz-jentoft AJ,Baeyens JP,Bauer JM,etal.Sarcopenia:European consensus on definition and diagnosis:report of the European working group on sarcopenia in older people〔J〕.Age Ageing,2010;39:412-23.
5Goodpaster BH,Park SW,Harris TB,etal.The loss of skeletal muscle strength,mass,and quality in older adults:the health,aging and body composition study〔J〕.J Gerontol A Biol Sci Med Sci,2006;(61):1059-64.
6Husom AD,Ferrington DA,Thompson LV,etal.Age-related differences in the adaptive potential of type I sketletal muscle fibers〔J〕.Exp Gerontol,2005;(40):227-35.
7Riley DA,Bain JL,Romatowski JG,etal.Skeletal muscle fiber atrophy:altered thin fiklament density changes slow fiber force and shortening velocity〔J〕.Am J Physiol,2005;(288):C360-5.
8Salazar JJ,Michele DE,Brooks SV.Inhibition of calpain prevents muscle weakness and disruption of sarcomere structure during hindhmb suspension〔J〕.J Appl Physiol,2009;(108):120-7.
9Suetta C,Hvid LG,Justesen L,etal.Effects of aging on human skeletal muscle after immobilization and retraining〔J〕.J Appl Physiol,2009;(107):1172-80.
10Zhong S,Lowe DA,Thompson LV.Effects of hindhmb unweighting and aging on rat semimembranosus muscle and myosin〔J〕.J Appl Physiol,2006;(101):873-80.
11Mcardle A,Dillmann WH,Mestril R,etal.Overexpression of HSP70 in mouse skeletal muscle protects against muscle damage and age-related muscle dysfunction〔J〕.FASEB J,2004;(18):355-7.
12Moran AL,Warren GL,Lowe DA.Soleus and EDL muscle contractility across the lifespan of female C57BL/6 mice〔J〕.Exp Gerontol,2005;(40):966975.
13Morse CI,Thom JM,Reeves ND,etal.In vivo physiological cross-sectional area and specific force are reduced in the gastrocnemius of elderly men〔J〕.J Appl Physiol,2005;(99):1050-5.
14Ochala J,Frontera WR,Dorer DJ.Single skeletal muscle fiber elastic and contractile characteristics in young and older men〔J〕.J Gerontol A Biol Sci Med Sci,2007;(62):375-81.
15Prochniewicz E,Thomas DD,Thompson LV.Age-related decline in actomyosin function〔J〕.J Gerontol A Biol Sci Med Sci,2005;(60):425-31.
16Alnaqeeb MA,Alzaid NS,Goldspink G.Connective tissue changes and physical properties of developing and ageing skeletal muscle〔J〕.J Anat,1984;(139):677-489.
17Visser M,Goodpaster BH,Kritchevsky SB,etal.Muscle mass,muscle strength,and muscle fat infiltration as predictors of incident mobility limitations in well-functioning older persons〔J〕.J Gerontol A Biol Sci Med Sci,2005;(60):324-33.
18Kent-Braun JA,NG AV,Young K.Skeletal muscle contractile and noncontractile components in young and older women and men〔J〕.J Appl Physiol,2000;(88):662-8.
19Payne AM,Dodd SL,Leeuwenburgh C.Life-long calorie restriction in Fischer 344 rats attenuates age-related loss in skeletal muscle-specific force and reduces extracellular space〔J〕.J Appl Physiol,2003;(95):255-62.
20Delbono O,O′rourke KS,Ettinger WH.Excitation-calcium release uncoupling in aged single human skeletal muscle fibers〔J〕.J Membr Biol,1995;(148):211-22.
21Gonziiez E,Messi ML,Zheng Z,etal.Insulin-like growth factor-l prevents age-related decrease in specific force and intracellular Ca2+in single intact muscle fibres from transgenic mice〔J〕.J Physiol,2003;(552):833-44.
22Jimenez-Moreno R,Wang ZM,Gerring RC,etal.Sarcoplasmic reticulum Ca2+release declines in muscle fibers from aging mice〔J〕.Biophys J,2008;(94):3178-88.
23Renganathan M,Messi ML,Delkbono O.Dihydropyridine receptor- ryanodine receptor uncoupling in aged skeletal muscle〔J〕.J Membr Biol,1997;(157):247-53.
24Renganathan M,Messi ML,Delbono O.Overexpression of IGF-I exclusively in skeletal muscle prevents age-related decline in the number of dihydropyridine receptors〔J〕.J Biol Chem,1998;(273):28845-51.
25Ryan M,Carlson BM,Ohlendieck K.Oligomeric status of the dihydropyridine receptor in aged skeletal muscle〔J〕.Mol Cell Biol Res Commun,2000;(4):224-9.
26Boncompagni S,D′amelio L,Fulle S,etal.Progressive disorganization of the excitation-contraction coupling apparatus in aging human skeletal muscle as revealed by electron microscopy:a possible role in the decline of muscle performance〔J〕.J Gerontol A Biol Sci Med Sci,2006;(61):995-1008.
27Damiani E,Larsson L,Margreth A.Age-related abnormalities in regulation of the ryanodine receptor in rat fast twitch muscle〔J〕.Cell Calcium,1996;(19):15-27.
28Ryan M,Butler-Browne G,Erzen I,etal.Persistent expression of the 1S-dihydropyridine receptor in aged human skeletal muscle:implications for the excitation-contraction uncoupling hypothesis of sarcopenia〔J〕.Int J Mol Med,2003;(11):425-34.
29YU F,Hedstrom M,Cristea A,etal.Effects of ageing and gender on contractile properties in human skeletal muscle and single fibres〔J〕.Acta Physiol (Oxf),2007;(190):229-41.
30Westerblad H,Bruton J D,Lannergren J.The effect of intracellular pH on contractile function of intact,single fibres of mouse muscle declines with increasing temperature〔J〕.J Physiol,1997;(500):193-204.
31Lowe DA,Surek JT,Thomas DD.Electron paramagnetic resonance reveals age-related myosin structural change:in rat skeletal muscle fibers〔J〕.Am J Physiol,2001;(280):C540-7.
32D′Antona G,Pellegrino MA,Adami R,etal.The effect of ageing and immobilization on structure and function of human skeletal muscle fibres〔J〕.J Physiol,2003;(552):499-511.
33Thompson LV,Durand D,Fugere NA,etal.Myosin and actin expression and oxidation in aging muscle〔J〕.J Appl Physiol,2006;(101):1581-7.
34Balagopal P,Schimke JC,Ades P,etal.Age effect on transcript levels and synthesis rate of muscle MHC and response to resistance exercise〔J〕.Am J Physiol,2001;(280):8203-8.
35Acharyya S,Guttridge DC.Cancer cachexia signaling pathways continue to emerge yet much still points to the proteasome〔J〕.Clin Cancer Res,2007;(13):1356-61.
36Van Hees HW,Li YP,Ottenheijm CA,etal.Proteasome inhibition improves diaphragm function in congestive heart failure rats〔J〕.Am J Physiol,2008;(294):L1260-8.
37Visser M,Pahor M,Taaffe DR,etal.Relationship of interleukin-6 and tumor necrosis factor-alpha with muscle mass and muscle strength in elderly men and women:the Health ABC Study〔J〕.J Gerontol A Biol Sci Med Sci,2002;(57):M326-32.
38Thomas DD,Cooke R.Orientation of spin-labeled myosin heads in glycerinated muscle fibers〔J〕.Biophys J,1980;(32):891-906.
猜你喜欢
食品工业科技(2022年17期)2022-08-27
食品科学(2022年7期)2022-05-13
现代食品科技(2022年4期)2022-04-28
青年文学家(2021年31期)2021-12-12
铁道科学与工程学报(2021年9期)2021-10-20
食品工业科技(2021年3期)2021-06-16
食品科学(2020年11期)2020-07-13
东坡赤壁诗词(2018年6期)2018-12-22
郑州大学学报(医学版)(2015年2期)2015-02-27
食品工业科技(2014年23期)2014-03-11
