
鼢鼠土丘演替对植物功能群与生产力的影响
2014-11-07 01:46张灵菲张新中张燕堃冶娜娜傅华江小雷
草业学报 2014年2期
张灵菲,张新中,张燕堃,冶娜娜,傅华,江小雷
(兰州大学草地农业科技学院 草地农业生态系统国家重点实验室, 甘肃 兰州 730020)
啮齿动物是草地生态系统中一类极为重要的食草动物,如荒漠草原的更格卢鼠(Dipodomysmerriami, kangaroo rats)[1]、北美大草原的旱獭 (prairie dogs)[2-4]和囊鼠(Geomyidae, pocket gophers)等[5-6]。许多研究表明,啮齿动物的采食和挖掘活动对植被的组成和结构 (如物种多样性)、土壤的理化性质 (如土壤通透性、土壤肥力及土壤微生物等) 产生重要影响,进而影响到草地生态系统功能。其干扰活动极大地改变了原有生境的资源分布格局[7-9], 为其他生物的生存创造了新的环境, 因而被喻为“生态系统工程师”[7]。
营地下生活的高原鼢鼠 (Myospalaxfontanerii) 作为高寒草甸草食动物的重要组成成员, 其掘土造丘活动对高寒草甸生态系统同样有重要的影响[10-11]。高原鼢鼠在青藏高原高寒草甸的分布面积可达12%以上,其种群密度可达70只/hm2左右[12]。平均而言,每只鼢鼠每年可造土丘242个,每个土丘的直径约20~70 cm[13]。在重度退化草地, 鼠丘面积可占草地面积的70%左右。鼠丘的形成造成了草地生物多样性的下降,植物群落结构发生了较大变化,毒杂草比例增加,优质牧草种类和数量减少,饲草品质和草地利用率大幅度下降,但同时也开始了土丘上植物群落的次生(恢复)演替过程。研究表明,各类挖掘类啮齿动物干扰后形成的土丘是维持草地生态系统植物群落物种多样性的重要因素[5-6]。不同演替时期的鼠丘,其演替轨迹、物种组成、群落结构及生态过程大不相同,从而形成了高寒草地生态系统中复杂多样的斑块性生境,为具备各种不同竞争能力和入侵能力植物种的定居创造了条件[11]。而不同的物种组成又改变了草地的群落结构及牧草的饲用价值,进而影响到草地生态系统的生产力。因此, 研究鼠丘植物群落演替过程中植物功能群及生产力的变化特征,对深刻认识高寒草甸生态系统的本质及草地资源的合理开发、利用有重要意义。
功能群是植物生态学研究中常借助的一个重要的功能分类单位,作为一种有效的手段常被广泛应用于植物群落响应干扰和全球变化的研究以及生物多样性与生态系统功能关系的研究中[14-16]。用功能群来解释不同演替梯度或生境中物种多样性格局的变化及生态系统功能关系将使复杂的问题变得较为简化且更具说服力[17]。因而植物功能群的方法被广泛应用于不同生境中植物群落的演替、生物多样性、干扰及环境变化对生态系统影响的研究中[18-19]。然而,高寒草甸生态系统中不同鼢鼠土丘演替对植物功能群与生产力关系影响的研究尚末见报道。本研究力求从生态学理论和生产实践两种不同的角度出发,探讨鼢鼠土丘演替过程中生活型功能群和牧草适口性功能群的变化及其对高寒草甸生态系统生产力的影响,揭示高寒草甸鼢鼠干扰后的自然恢复规律,从而为退化草地的治理与恢复提供科学依据。
1 材料与方法
1.1 实验地概况
研究设于甘肃省玛曲县万仓乡所辖区内。地理坐标为N 33°43′45″,E 101°44′50″,海拔3500 m。地貌为沟谷滩地,土层深厚,土壤为亚高山草甸土,有机质含量4.07%~5.02%。年均温1.4℃,年均降水量463.9 mm。研究样地为1998年设置的永久性鼢鼠危害研究草地,面积为15 hm2。为观察鼠丘的自然演替过程,防止放牧家畜及其他大型草食动物的干扰,样地自1998年起设置了围栏。围栏内草地类型、环境条件及植被组成相对一致。样地自建成后不断有鼢鼠入侵,因此样地内包含有新土丘到8年以上的旧土丘。不同年限的土丘根据原有实验的标记及土丘的土壤、植被状况确定[20]。样地内的植被为亚高山草甸,属冬季牧场,草地退化明显。主要植物组成为垂穗披碱草(Elymusnutans)、洽草(Koeleriacristata)、矮嵩草(Kobresiahumilis)、甘肃嵩草(K.kansuensis)、鹅绒委陵菜(Potentillaanserina)、二裂委陵菜(Potentillabifurca)、马先蒿(Pedicularissp.)、乳白香青(Anaphalislactea)、细叶亚菊(Ajaniatenuifolia)、草玉梅(Nemonerivularis)等。
1.2 实验设计与取样
于2011年7月牧草生长高峰期在试验区内以空间序列代替时间序列的方法,对不同恢复演替阶段鼢鼠鼠丘植物群落物种组成、物种多样性变化及不同植物功能群的变化进行了调查。按鼠丘形成的年限将其分为0(当年新土丘),2,4,6和8年5个演替阶段。于每个阶段内选择10个鼠丘,测定鼠丘上(30 cm×30 cm)的植物种类、各种植物的平均高度、盖度、密度和生物量。最后将样品带回实验室70℃条件下烘干48 h, 称恒重。
1.3 数据分析
根据高寒草甸植物群落物种组成的特点, 按照草地生态系统研究中常用的方法[21], 将其按生活型功能群分为: 一年生或二年生植物、多年生禾草、多年生杂类草和莎草类4个功能群。牧草适口性功能群按家畜对其的喜好程度分为: 毒草、劣等、中等、良等和优等牧草5个功能群。
分别用物种数 (S) 和Shannon 指数代表物种多样性,Shannon 指数的计算方法为:
H′=∑(ni/N) ln (ni/N)
式中,H′代表Shannon指数,ni表示种i在样方中的生物量,N表示样方中所有物种的生物量。
用地上生物量代替生产力指标。 分别以不同演替阶段内10个土丘的生物量平均值、Shannon 指数平均数代表各演替阶段的生产力和Shannon 指数(H′), 以10个土丘内的物种总数代表不同演替阶段的物种多样性(S)。
用SPSS 13.0统计分析软件对不同演替阶段功能群变化数据进行方差分析, 对演替与功能群及生产力间的关系进行相关性分析。
2 结果与分析
2.1 演替对物种多样性与生产力的影响
调查中共记载植物87种, 其中禾本科8种,莎草科7种,豆科4种,杂类草68种。单个鼠丘上的植物种数变化于2种(新土丘) 至39种(8年土丘)之间,Shannon 指数变化于0.56~2.79之间, 生产力变化于56.80~299.44 g/m2之间。物种数和Shannon 指数均随演替时间的延长而增加, 二者间呈显著正相关关系 (物种数:Y=4.55x+0.6,R2=0.8914,图1A; Shannon 指数:Y=0.277x+0.364,R2=0.8234,图1B), 生产力首先随演替时间的增加而增加, 当演替时间为6年左右时, 生产力达最高水平, 之后随鼠丘演替时间的延续呈下降趋势, 二者间呈单峰曲线关系 (Y=-5.5514x2+69.403x+56.349,R2=0.9553, 图1C)。由此可见, 鼢鼠土丘的演替对植物群落的物种组成, 物种多样性及草地生产力均有促进作用。
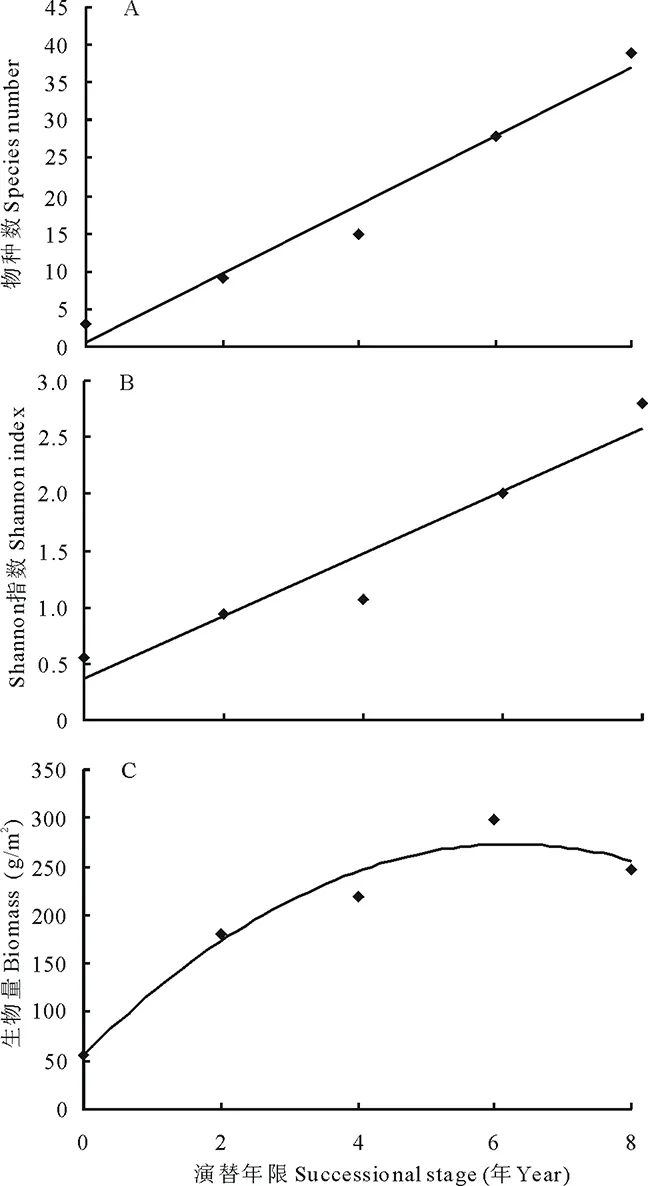
图1 演替对物种多样性和生产力的影响Fig.1 Effects of succession on species diversity and productivity
2.2 对生活型功能群的影响
鼢鼠土丘的演替对各生活型功能群物种多样性 (H′) 的影响有所不同 (图2)。一年生 (或二年生)植物的物种多样性随着演替的进程而减少, 二者间呈显著负相关关系 (Y=-0.092x+1.256,R2=0.4369, 图2A)。多年生禾草的物种多样性随演替的进程而增加, 二者间呈显著正相关关系 (Y=0.177x+0.016,R2=0.9563, 图2B)。莎草类的物种多样性先随演替的进程增加, 到演替的第6年时物种多样性达到最大值, 之后不再随演替年限的增加而增加, 而是趋于平缓, 二者间呈显著的单峰曲线格局 (Y=-0.0192x2+0.3004x+0.0291,R2=0.7096, 图2C)。同莎草功能群类似, 多年生杂类草的物种多样性与演替间也呈现单峰曲线关系 (Y=-0.0223x2+0.4501x+0.2394,R2=0.8213, 图2D)。
鼠丘的演替对生活型功能群生产力的影响与物种多样性略有不同 (图3)。一年生植物的生产力随着演替时间的增加而下降, 在演替的初期, 一年生物种在群落中占主导地位, 第1年和第2年分别占总生物量的80%和55%。但随着演替年限的增加, 其在群落中的生物量急剧下降, 到第4年其生物量仅占总生物量的 4%左右, 二者间呈显著的负相关关系 (Y=-22.968x+170.61,R2=0.8835, 图3A)。多年生禾草和莎草类的生物量则在演替后期达到最大, 二者与演替间的关系分别分单峰曲线 (Y=-0.571 4x2+13.688x-6.4294,R2=0.891, 图3B) 和正相关关系 (Y=5.8581x-0.0291,R2=0.7296, 图3C)。多年生杂类草生物量的峰值则出现在演替的中期阶段, 其与演替间的关系也为单峰曲线 (Y=-4.4311x2+47.566x+19.006,R2=0.8591, 图3D)。

图2 演替对生活型功能群物种多样性的影响Fig.2 Effects of succession on diversity of life form group
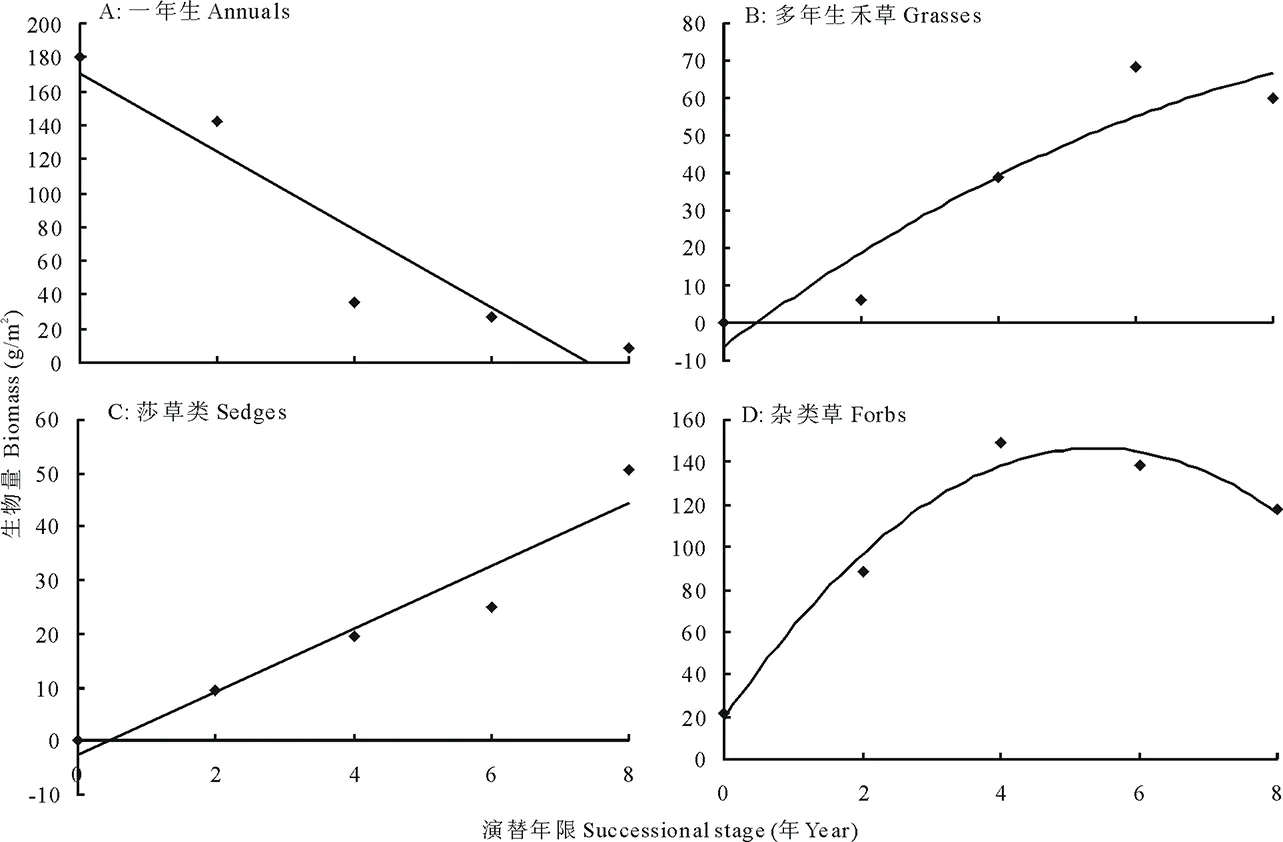
图3 演替对生活型功能群生产力的影响Fig.3 Effects of succession on productivity of life form group
2.3 对牧草适口性功能群的影响
鼢鼠土丘的恢复演替极大地改变了牧草品质和草地利用率。随着演替年限的增加, 有毒牧草和良等牧草功能群的物种多样性呈现出先增 (2~6年间) 后降 (6年以后) 的变化趋势, 二者间与演替时间的关系均为单峰曲线格局 (毒草:Y=-0.0198x2+0.3611x-0.2006,R2=0.7301, 图4A; 良等牧草:Y=-0.032x2+3.0021x-0.9327,R2=0.734, 图4D); 而劣等、中等和优等牧草功能群的物种多样性均随着演替年限的延长大幅度增加, 三者与演替年限间均呈正相关关系 (劣等牧草:Y=0.093x-0.044,R2=0.6587, 图4B; 中等牧草:Y=0.1792x+0.426,R2=0.7327, 图4C; 优等牧草:Y=0.2593x+0.1984,R2=0.7751, 图4E)。
不同适口性功能群的生产力也受到鼠丘演替的极大影响 (图5)。同物种多样性一样,有毒牧草和良等牧草的生物量亦呈现出先升后降的变化趋势,二者间与演替年限间均呈单峰曲线关系(有毒牧草:Y=-1.4761x2+17.682x-5.8806,R2=0.542,图5A;良等牧草:Y=-2.8119x2+31.2801x+14.076,R2=0.6018,图5D)。劣等牧草的生物量随演替年限的增加而减少, 二者间表现为负相关关系(Y=-6.225x+61.578,R2=0.3826, 图5B)。而中等牧草和优等牧草的生物量变化则与之不同, 均随演替年限的延长而急剧增加, 二者与演替间呈正相关关系 (中等牧草:Y=7.198x+1.558,R2=0.8035, 图5C; 优等牧草:Y=22.546x-25.208,R2=0.7903, 图5E)。
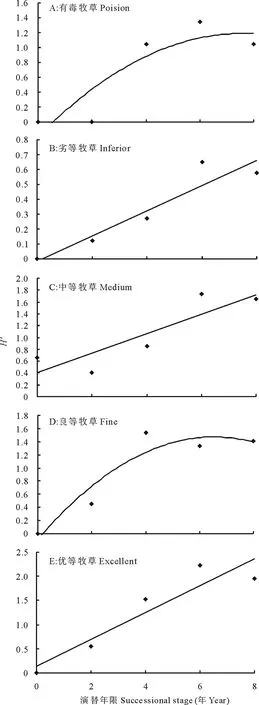
图4 演替对牧草品质功能群物种多样性的影响Fig.4 Effects of succession on diversity of forage quality group
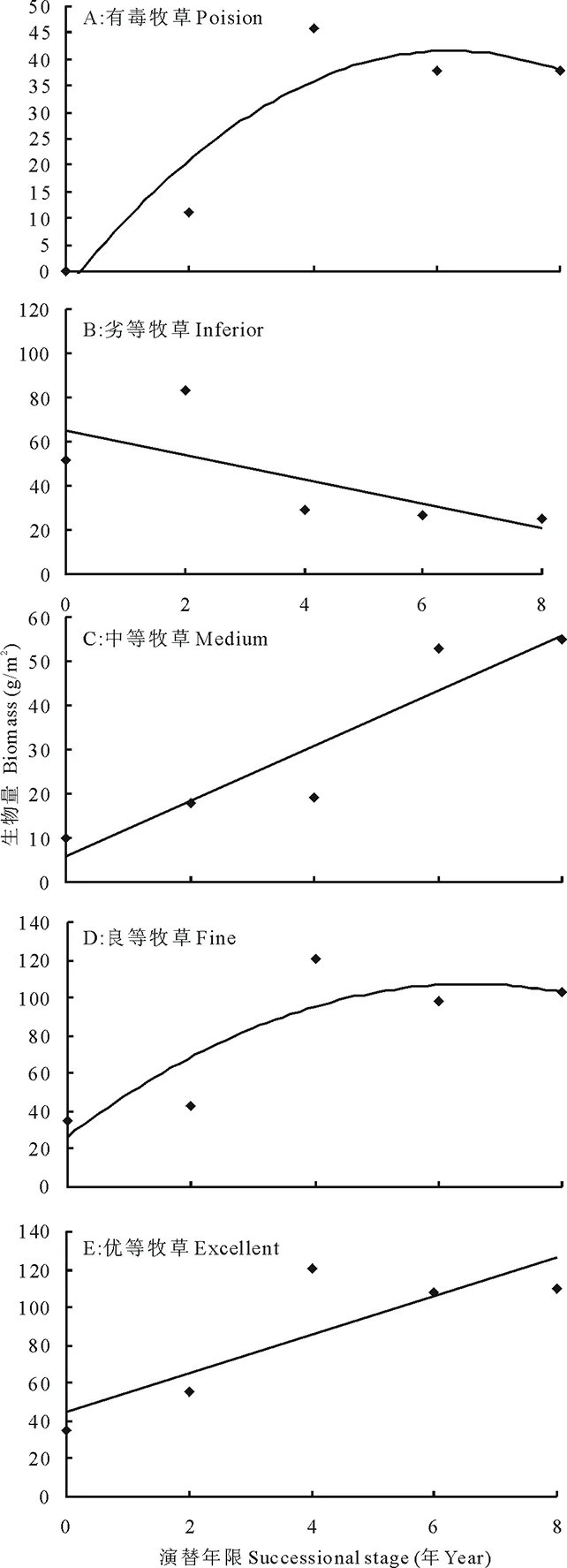
图5 演替对牧草品质功能群生产力的影响Fig.5 Effects of succession on productivity of forage quality group
3 讨论
高原鼢鼠对草地的影响主要是通过掘土造丘活动而产生的。其挖掘活动将地下土壤堆至地表, 形成了理化性质完全不同于周围生境的土丘斑块[22], 为其他物种的生存创造了条件[11]。随着土丘的形成, 开始了土丘上植物群落的演替过程, 从而导致草地植物群落的物种组成和多样性发生极大的变化[6]。关于草地演替过程中物种多样性的变化格局, 有两种不同的理论解释。一种理论认为, 在演替的过程中, 植物种类随时间的延长而逐渐增加, 至演替后期达到最大值, 即物种多样性随演替时间的延长而增加[23]。另一种理论认为, 由于短命植物和长寿植物的共存, 物种多样性在演替的中期阶段达到峰值, 即演替的中度干扰理论[24]。本研究结果中植物总体多样性 (物种数和H′) 变化格局符合第1种演替理论, 即物种多样性随演替时间的延长而增加, 二者间呈正相关关系 (图1A, B)。该结果与其他地下啮齿动物土丘演替的研究结果类似, 证明鼠丘改变了原有草地植物组成的格局, 其演替为不同竞争能力物种的替换和共存创造了条件, 因而导致物种多样性的增加[25-26]。而总体生产力变化模式则符合第2种理论: 即生产力水平在鼠丘演替的中期阶段达到最大 (图1C)。 同时, 生产力水平与物种多样性关系也符合草地生态系统中物种多样性-生产力关系研究模式: 即物种多样性中等水平时, 生产力达到最大值[27-28]。
鼠丘的演替对不同生活型功能群的影响则有很大区别 (图2,图3)。鼠丘的形成改变了原有的土壤特性和物种竞争关系, 为 “杂草对策型” 物种的侵入创造了有利条件, 因而一年生物种的多样性和生产力水平在演替初期都很高[29]。但随着演替的进展, 先期入侵种使环境条件发生改变, 增加了鼠丘土壤的异质性[6], 使其更加适宜其他物种的入侵, 进而发生竞争替代作用, 其结果是先期入侵的一年生植物多样性和生产力急剧下降 (图2A,图3A), 而竞争耐受型物种多年生禾草、莎草类和多年杂类草的物种多样性和生产力则不断上升 (图2B, C, D, 图3B, C, D), 直至演替的中、后期达到峰值。该结果显示, 鼢鼠鼠丘植被的演替轨迹与其他小型哺乳动物干扰地的植被演替动态类似[6,26], 随着演替时间的延长, 不同生活对策的物种交替轮换, 相互共存, 直至达到平衡状态[25-26,30]。但也有研究表明, 局部干扰地演替过程中物种多样性的变化模式取决于斑块内的有效资源水平[9], 因而, 小型哺乳动物干扰后土丘上植物多样性变化表现为不同的格局[25]。
鼢鼠土丘演替对牧草适口性功能群也有较大的影响 (图4,图5)。适口性是体现植物竞争能力的重要功能特征之一, 与其生存环境有着密切的联系[31]。一些利用植食性昆虫的研究表明, 植物的食口性与植物群落的演替密切相关[32-33]: 演替初期, 由于 “杂草型” 植物所具备的防御对策主要是逃避, 而不是分泌较多的次生化合物, 因而其适口性一般较好[34]。随着演替阶段的延长, 占据优势地位且具有较强竞争能力的物种多依赖于次生化合物分泌作为防御手段, 因而其适口性较差[35]。与上述结果不同, 本研究结果表明, 在演替初期, 各适口性功能群的物种多样性和生产力水平均较低, 即可食性牧草总量较少。随着演替时间的延长, 各功能群的多样性虽然都有程度不同的增加, 但总体而言, 中等, 良等, 优等牧草多样性(中, 良, 优H′分别为1.41~1.83, 0~1.18, 0~2.09, 图4C, D, B) 和生产力 (中, 良,优分别为5.72~60.27 g/m2, 21.16~118.04 g/m2, 0~180.76 g/m2, 图5C, D, B) 的增加幅度更大, 而毒草和劣等牧草的多样性(毒, 劣H′分别为0~1.25, 0~0.69, 图4A, B)和生产力水平(0~50.28 g/m2)增幅较小或急剧下降(101.28~8.26 g/m2), 且毒杂草在植物群落中所占比例较低。因此, 牧草的适口性是随着演替过程的延长而增加的, 即草地的牧草品质是随着鼠丘演替时间的延长而提高的。这可能是由于研究中所使用的草食动物种类不同, 各自具备独特的进化史和生理代谢特征, 对不同植物种的喜好程度不同所致。其他研究中以喜食双子叶植物的无脊椎草食动物的适口性为评判标准[33], 而本研究中是以喜食禾草的有蹄类家畜(绵羊)的适口性为评判标准。此外, 研究所涉及的草地生态系统类型和干扰类型亦不相同, 因而植物群落的演替轨迹及群落结构的变化格局也存在较大差别[6,18]。
综上所述, 本研究从生态学理论和生产实践两种不同的角度出发,探讨鼢鼠土丘恢复演替对生活型功能群和牧草品质功能群的变化及其对高寒草甸生态系统生产力的影响。结果表明,鼢鼠土丘的演替对植物群落的物种组成,功能群结构及草地生产性能等均产生了重要影响。无论是植物功能群的物种多样性, 还是草地生产力水平, 均随鼠丘的演替有程度不同的提高。研究证明, 利用功能群理论研究鼠丘演替过程中植物群落及生态系统功能(生产力)的变化,能够较好地揭示高寒草甸生态系统植被响应生物干扰的对策。
猜你喜欢
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
今日农业(2021年10期)2021-07-28
兽医导刊(2020年3期)2020-12-30
东坡赤壁诗词(2020年4期)2020-09-02
防护林科技(2019年7期)2019-03-25
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20
草业科学(2018年11期)2018-11-28
中国诗歌(2018年3期)2018-11-14
美文(2018年6期)2018-03-28
