
云南横断山区蚤类科、属、种多样性空间分布特征及其关系*
2014-11-10 01:27龚正达高子厚琚俊科连宏宇段兴德张丽云
寄生虫与医学昆虫学报 2014年1期
李 栋 龚正达 高子厚 琚俊科 连宏宇 段兴德 张丽云
(1.大理学院公共卫生学院,云南大理 671000; 2.云南省地方病防治所,云南大理 671000; 3.潞安矿业集团总医院,山西 046204)
物种多样性的大空间尺度格局研究主要是生物类群间物种数的协同变化、物种和高级分类阶元的关系、局域物种多样性与区域物种多样性关系以及全球气候变化影响等(Meserveetal.,1999; Gastonetal.,2000)。许多研究表明,一地区许多类群中物种数量与其高级分类阶元数量间具有很高的相关关系,如植物、兽类、鸟类、两栖类类等。这种关系具有普遍意义,无论是不同地理尺度和区域,还是类群都存在(Gaston,2000a),这表明物种在科、属水平上的频次分布有一定的规律性和物种和高级阶元的数量相关应是规律性的。因此,有人探讨利用高级分类阶元代替物种作为生物多样性的度量单位来进行热点地区的选择和评价,发现两者效果一致,认为高级分类阶元的应用可大大节约了时间和费用,具有应用价值(Balmfordetal.1996a,1996b;Balmfordetal.,2000;Gaston,2000c)。在国内,也有学者对鸟类、哺乳类等动物高等分类阶元丰富度,如属、科、目等丰富度与物种丰富度之间的关系进行了系列研究,研究和探讨它们能否成为物种丰富度的替代物等(冯永军等,2006;郑孜文 等,2008)。 这种做法在古生物学中已被广泛应用(Sepkoski,1991,1998;Gaston 2000a),但在现代生物学中的应用却受到争议(Williamsetal.,1994;Gaston,2000b)。因此,不同区域或尺度物种与高级分类阶元之间的数量关系、局域物种多样性与区域物种多样性关系、高级分类阶元代替物种作为生物多样性的度量单位进行热点地区的选择和评价等问题的研究和探讨具有重要意义。
国内关于病媒生物的相关研究,见有蚤类物种与高级分类阶元在数量关系研究(连宏宇等,2009)和陆生哺乳动物与寄生蚤类丰富度的关系及协同进化趋势的研究和探讨(连宏宇等,2011)。我国地域辽阔,东西和南北跨度很大,其中山地、高原和丘陵约占总积的65%,从南到北或从低到高,涵盖了从热带到寒温带的各种气候环境。上述不同自然地理环境形成的地域差异、气候作用与干扰等,可能对不同地区或局域宿主与寄生虫多样性及其关系之间产生重要影响。因此,宿主动物、寄生虫多样性及它们之间的关系沿着不同气候或地理环境梯度的变化趋势、规律与格局,以及不同动物地理区系过渡带或交错区的边缘效应、重要环境因素等对它们的影响都是值得研究和探讨的问题(连宏宇等,2011)。
本研究以云南西部横断山区(约北纬21°00~29°00、东经97°30~102°00;海拔500~5 000 m)为研究的空间范围及尺度,在既往蚤类区系和多样性调查研究的基础上,就云南横断山区(局部地区)蚤类科、属、种丰富度之间的关系及其空间分布特征,不同山峰(局域)蚤类科、属、种丰富度之间相关度及其与物种多样性之间的关系,局域与区域物种多样性的关系以及地理环境因子对它们的影响等问题进行分析和探讨。
1 材料与方法
1.1 采集地点
云南横断山区位于云南西部(元江以西),是青藏高原的南延部分,北部有高黎贡山、怒山、云岭等巨大山脉和怒江、澜沧江及金沙江等大河,由北向南相间排列,形成了著名的三江并流的纵谷地区。这里不仅是反映地球演化事件的关键地区,也是亚洲大陆物种多样性最丰富的地区和多物种保存、分布、分化、形成及起源的中心。该区域海拔最低约210 m,最高达6 740 m,由于地理位置不同和高差大,气候有明显的变化。从南向北,或者由低到高依次可出现低山、中山、亚高山、高山、极高山地带和热带、亚热带、温带、寒带不同的气候带和森林植被类型。迄今,这里的蚤类已知有9科45属约153种(亚种)。其中,科的数量占我国已知蚤类科数的90%;属的数量占我国已知蚤属的60%,占青藏高原已知蚤属的90%;物种数占我国已知蚤种25%(龚正达和张丽云,2003;龚正达等,2007;连宏宇和龚正达,2009)。
1.2 资料与数据来源
云南西部横断山区蚤类区系和物种多样性的地理分布数据及资料,主要源于我们近年来的系列调查结果及论文,它们包括:中国“三江并流”纵谷地蚤类丰富度与区系沿纬度梯度的水平分布格局(龚正达等,2007);云南横断山区蚤类物种丰富度与区系的垂直分布格局(龚正达等,2005);云南横断山区蚤类多样性的地理分布趋势与重要环境因素的关系(龚正达等,2001a);以及能代表云南蚤类区系概貌的蚤类名录及《云南蚤类志》的区系分布总表(解宝琦和曾静凡,2000)等。此外,横断山区不同山峰蚤类区系和物种多样性数据及资料源于我们对永德大雪山(龚正达等,2000)、无量山(龚正达等、2004)、大理苍山(龚正达等,1999)、高黎贡山北部和南部(龚正达等,1996)、老君山(杨光荣和陶开会,1990)、白草岭(冯锡光等,2002)和玉龙雪山(段兴德等,2002)共8个山地蚤类物种多样性的系列调查结果。
1.3 数据的统计与分析
研究区海拔或纬度梯度的划分参照和沿用既往的方法与数据(龚正达等,2005;2007)。对不同空间发布类型中的蚤类物种与科、属的数量及其与物种多样性关系应用SPSS 16.0软件进行相关分析,首先用Kolmogorov-Smirnov检验方法对数据进行正态分布检验。符合正态分布的数据,采用Pearson线性相关分析。
2 结果
2.1 云南横断山区蚤类(科、属、种)丰富度的空间分布格局
2.1.1云南横断山区蚤类(科、属、种)丰富度沿纬度梯度的分布特征:横断山区蚤类已知有9科45属约153种(亚种)。但随着纬度梯度(北纬21°00′~29°00′)和气候环境的变化,蚤类的组成和分布明显不同(表1)。蚤类总体丰富度沿纬度梯度分布的主要情况见表1。 比较而言,蚤类的科、属、种丰富度在北纬25°00′~27°00′N(Ⅴ-Ⅶ带)之间均较高,而纬度较高和较低的区域的丰富度较低。
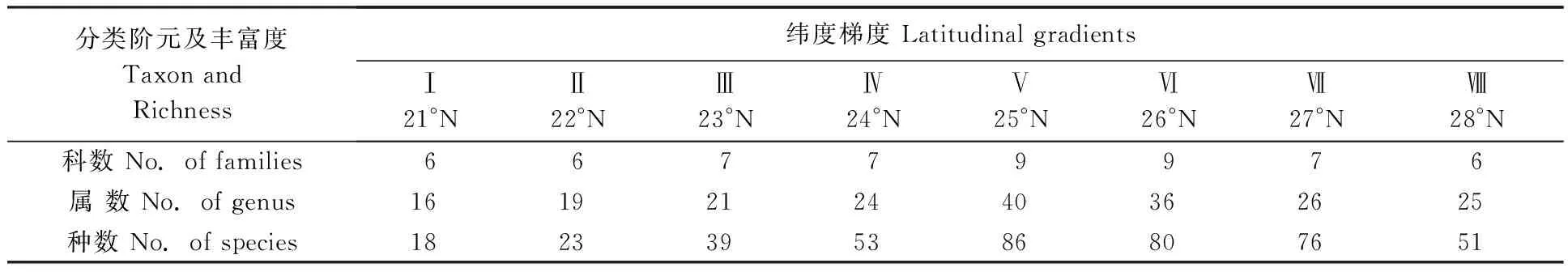
表1 云南横断山区蚤类科、属、种丰富度沿纬度梯度的变化Tab.1 The changes of family,genus and species of fleas richness along latitudinal gradients in Hengduan Mountains of Yunnan
2.1.2云南横断山区蚤类(科、属、种)丰富度沿海拔梯度的分布特征:随着海拔(500~5 000 m)梯度和气候环境的变化,蚤类的组成和分布明显不同。蚤类丰富度沿海拔梯度分布的主要情况见(表2)。相对而言,蚤类科和属丰富度在海拔2 500~2 900 m之间(E带)最高,而种丰富度在海拔2500~3 800 m(E~G带)之间较高,但科的丰富度在此区间2 900~3 800 m之间显得相对较低。
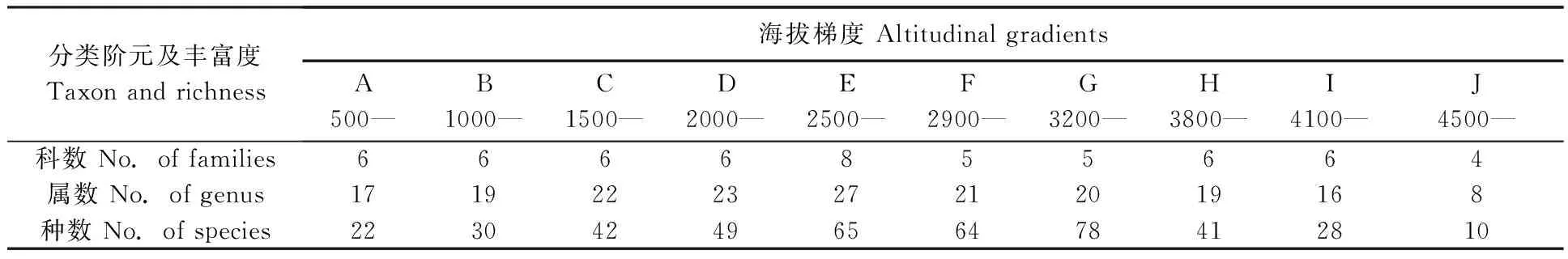
表2 云南横断山区蚤类科、属、种丰富度沿海拔梯度的变化Tab.2 The changes of family,genus and species of fleas richness along altitudinal gradients in Hengduan Mountains of Yunnan
2.2 云南横断山区蚤类种、属、科丰富度之间的关系
应用Kolmogorov-Smirnov方法,分别对云南蚤类8个纬度梯度和10个海拔梯度物种-属-科的数据进行分布型检验表明,均属正态分布,因此,应用Pearson线性相关分析。
2.2.1云南横断山区蚤类种、属、科丰富度之间沿纬度梯度分布(水平分布)的关系(图1~3;表3):相关分析显示,在横断山区北纬21°00′~29°00′之间8个纬度梯度带中蚤类种-属-科丰富度之间均呈显著正相关。其中种-属、属-科丰富度之间呈极显著相关,分别为(r>0.916;P<0.01和r>0.920;P<0.01),而种-科丰富度呈显著相关(r>0.829;P<0.05),前者大于后者,详见表3。
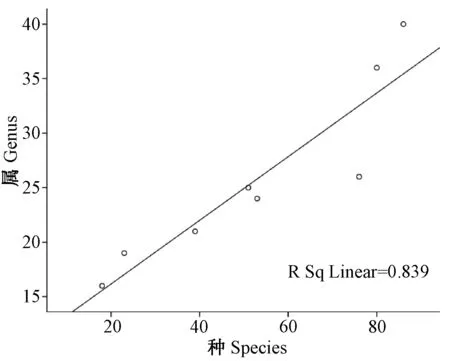
图1 蚤类种与属丰富度之间沿纬度梯度分布的关系图Fig.1 Relationship between species and genus in number of fleas along latitudinal gradients distribution
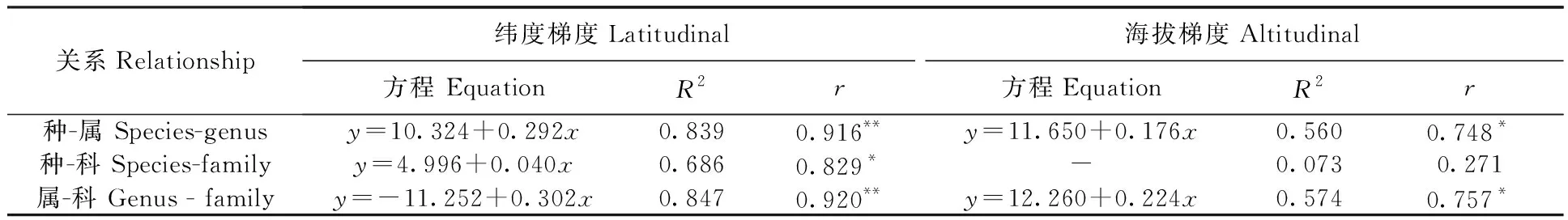
表3 云南横断山区蚤类种、属、科丰富度之间沿纬度和海拔梯度分布的相关关系Tab.3 Correlation relationship of family,genus and species of fleas richness along latitudinal and altitudinal gradients distribution in Hengduan Mountains of Yunnan
*表示相关显著(P<0.05); ** 表示相关极显著(P<0.01)。* show correlation level atP< 0.05; ** show correlation level atP< 0.01.
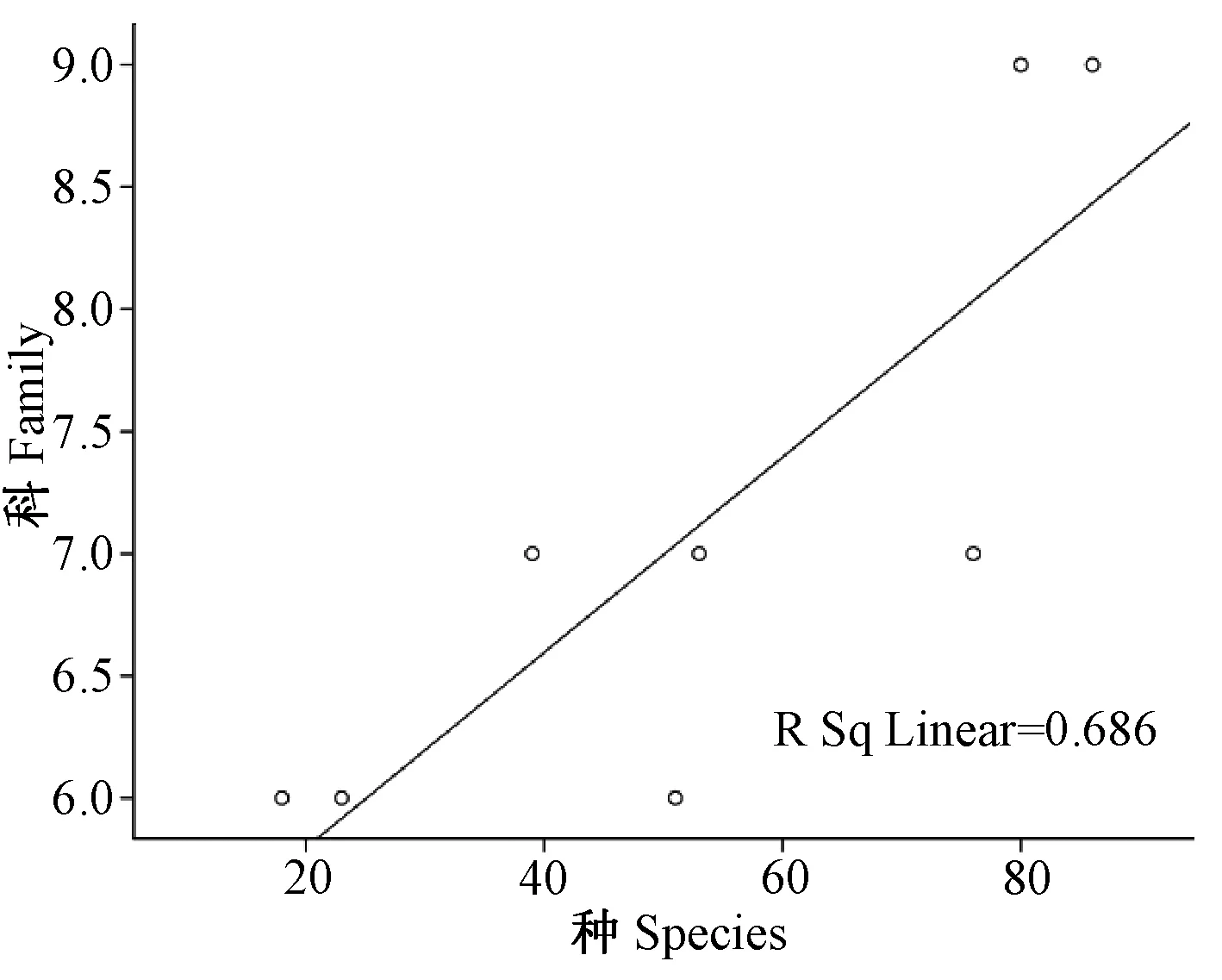
图2 蚤类种与科丰富度之间沿纬度梯度分布的关系图Fig.2 Relationship between species and family in number of fleas along latitudinal gradients distribution
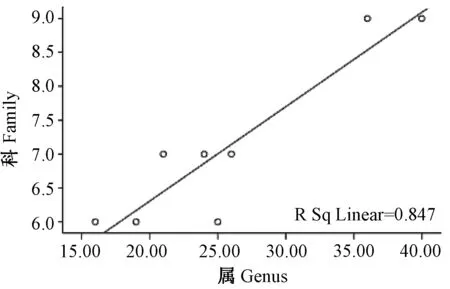
图3 蚤类属与科丰富度之间沿纬度梯度分布的关系图Fig.3 Relationship between genus and family in number of fleas along latitudinal gradients distribution
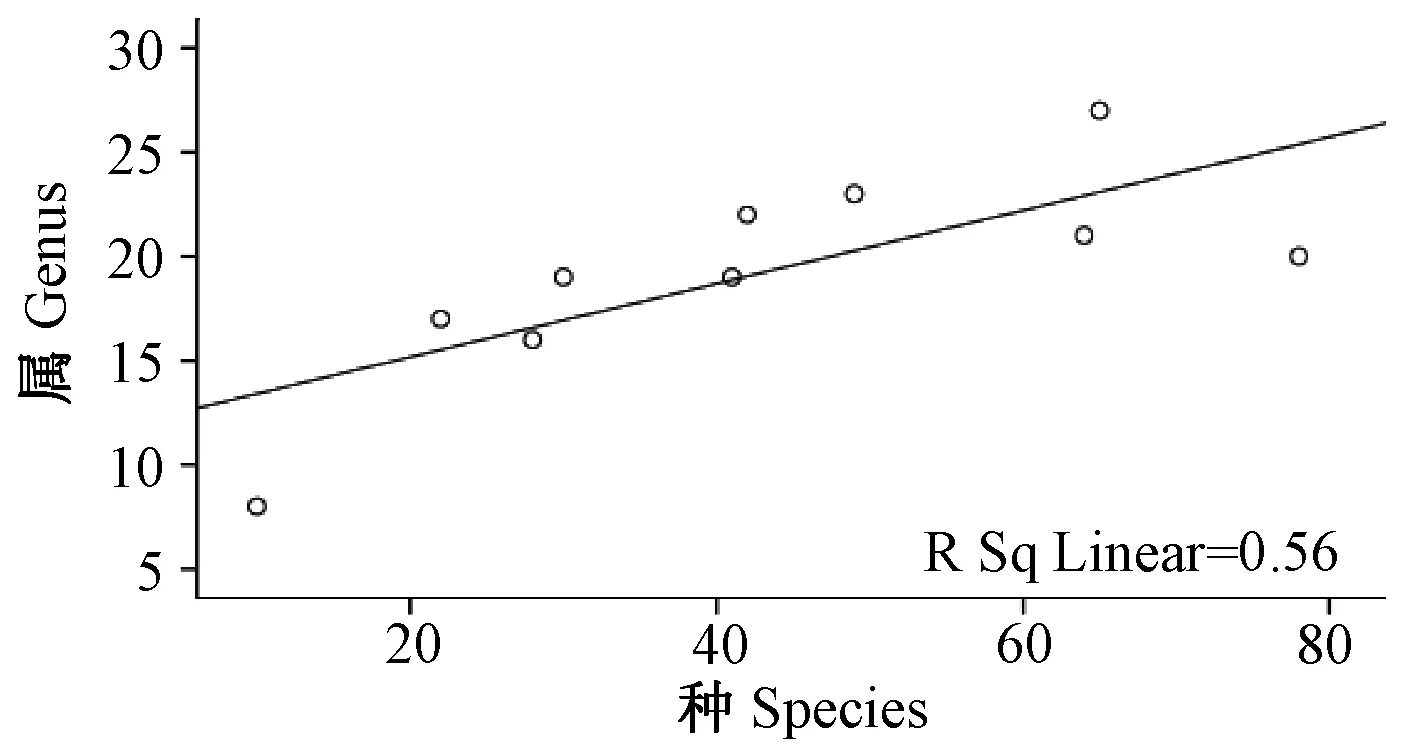
图4 蚤类种与属丰富度之间沿海拔梯度分布的关系图Fig.4 Relationship between species and genus in number of fleas along altitudinal gradients distribution
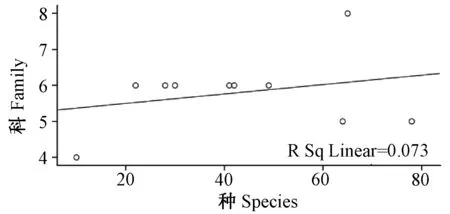
图5 蚤类种与科丰富度之间沿海拔梯度分布的关系图Fig.5 Relationship between species and family in number of fleas along altitudinal gradients distribution
上述结果反映了云南横断山区蚤类科、属、种之间沿纬度梯度分布中呈现的协同变化关系。
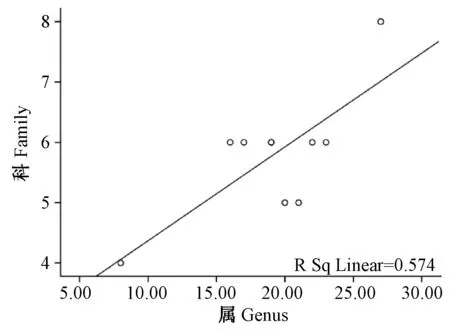
图6 横断山区蚤类科与属丰富度之间沿海拔分布的关系图Fig.6 Relationship between genus and family in number of fleas along altitudinal gradients distribution
2.2.2云南横断山区蚤类种、属、科丰富度之间沿海拔梯度分布(垂直分布)的关系(图4~6;表3):从图4、6中看出,蚤类在海拔500~5 000 m之间的10个梯度带中种-属和属-科的数量变化呈增长趋势。分析结果显示:种-属和属-科之间均呈现了显著正相关(r>0.748;P<0.05和r>0.757;P<0.05);但是,在种-科丰富度之间没规律(图5),并显示出种-科丰富度的之间相关性不显著(F= 0.271);P>0.05)。详见表3。
上述情况反映了云南横断山区蚤类科、属、种之间沿海拔梯度分布中呈现的变化、关系及其与纬度梯度分布上存在一定的差异。
2.3 横断山区8个山峰蚤类的丰富度、相关度及与物种多样性之间关系
2.3.1横断山区8个山峰不同森林植被带蚤类科、属、种丰富度及其分布(表4):由于横断山区8个山峰(永德雪山、无量山、大理苍山、高黎贡山北部(贡山)、高黎贡山(泸水)、老君山、白草岭、玉龙雪山)的地理位置、海拔高度、气候环境、森林植被带的差异,其蚤类科、属、种的组成和空间分布特征各不相同(表4)。各山峰在局域(或群落)水平上,蚤类丰富度随海拔梯度的变化及情况复杂得多。
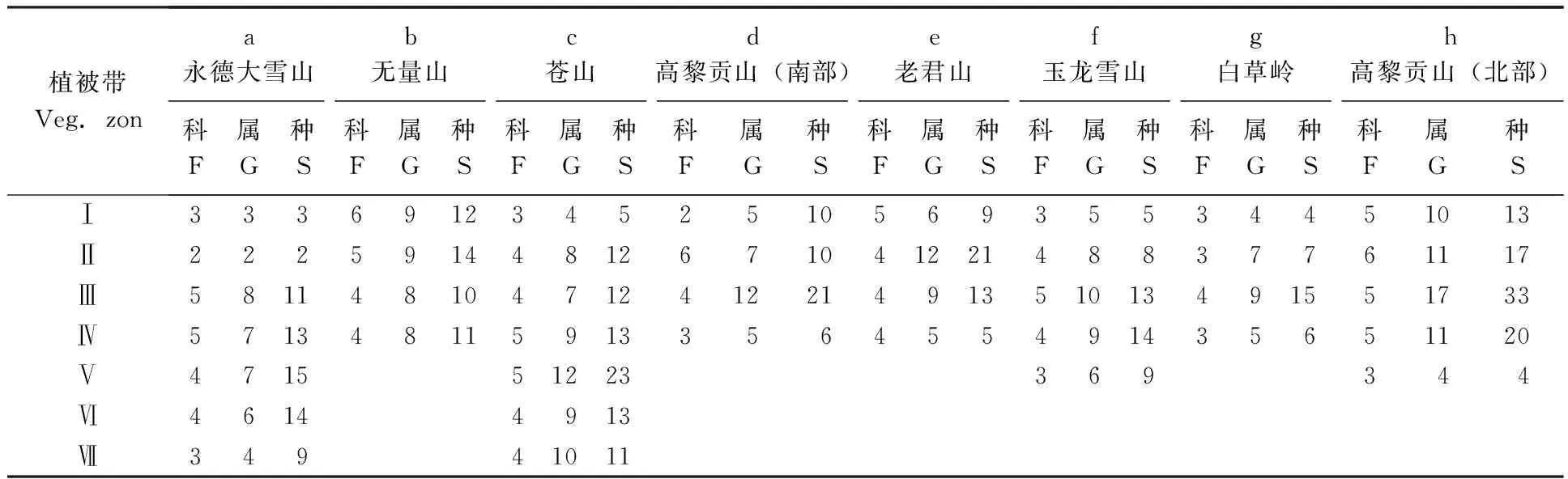
表4 云南横断山区不同山峰森林植被带蚤类的组成及分布情况Tab.4 Composition and distribution of fleas in different mountains in Hengduan Mountains of Yunnan
Veg.zon:植被带Vegetational zon;a:永德大雪山Yongde snow Mountains;b:无量山 Wuliang Mountains;c:苍山 Cangshan Mountains;d:高黎贡山(南部) South of Gaoligong Mountains;e:老君山 Laojun Mountains;f:玉龙雪山 Yulong snow Mountains;g:白草岭 Baicao Mountains;h:高黎贡山(北部) North of Gaoligong Mountains.表5同 The same with Tab.5.
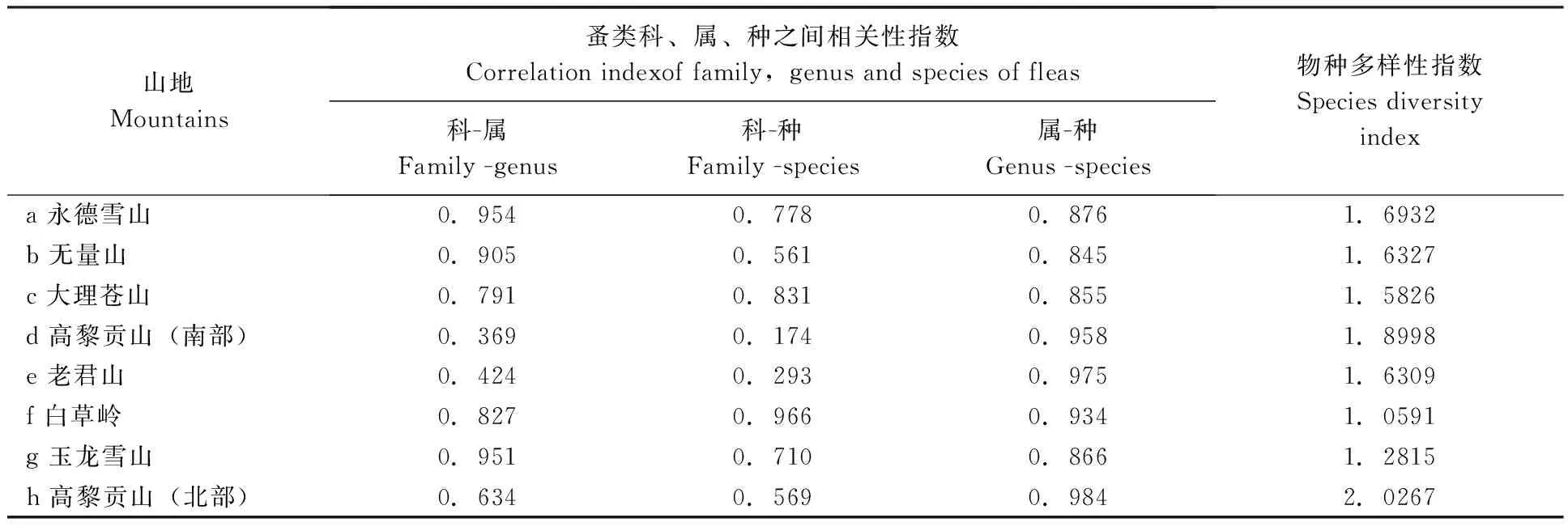
表5 云南横断山区不同山峰蚤类丰富度的相关度与物种多样性之间的关系Tab.5 Relationship between relevancy and species diversity of fleas richness in different mountains in Hengduan Mountains of Yunnan
2.3.2云南横断山区8个山峰蚤类科、属、种之间相关度与其物种多样性的关系:通过蚤类科、属、种之间的相关性(表5)测定后看出:由于地理位置和气候环境的差异,各山地之间蚤类科、属、种之间相关性和物种多样性指数明显不同和非一致性。比较而言,在蚤类科、属、种之间的相关性中,蚤类属-种和科-属的相关性多数大于科-种之间的相关性。但在物种多样性较低的白草岭例外。
通过横断山区8个山地蚤类科、属、种丰富度之间相关性与物种多样性指数之间相关分析,结果显示:蚤类科-属相关性与物种多样性指数之间呈负相关(r=-0.507,P>0.05);蚤类科-种相关性与物种多样性指数之间呈负相关(r=-0.634,P>0.05);蚤类属-种相关性与物种多样性指数之间呈正相关(r=0.359,P>0.05)。
除蚤类属-种之间相关性与物种多样性之间关系呈正相关外,蚤类科-属、科-种相关性与物种多样性指数之间关系呈负相关。 但它们的相关性与物种多样性指数之间的关系均不显著。
综上所述,不同山峰(局域)蚤类科、属、种丰富度之间相关性并非一致及其与物种多样性指数关系并不密切。同时,也反映了在横断山区不同空间尺度上,局域(山峰)蚤类科、属、种多样性空间分布的特征、关系及其与该区域(横断山区)分布特征之间的差异。
3 讨论
物种和高级分类阶元的关系主要包括两个方面的内容: 一是数量上的关系,即物种丰富度和属以上阶元的丰富度关系; 二是频次分布关系,即属以上阶元所含物种数的频次分布。
3.1 物种与属、科丰富度之间关系及其空间分布特征
物种与高级分类单元间数量上呈显著的相关关系具有普遍意义。这种结果在许多地区不同尺度上的分类类群中得到验证,有人并对此作了全面的总结(Gaston,2000b)。 云南由于地理位置不同和高差大,从南向北,或者由低到高依次可出现低山、中山、亚高山、高山、极高山地带和热带、亚热带、温带、寒带不同的气候带和森林植被类型。横断山区(局部地区)蚤类沿纬度梯度的水平分布中,物种-属、种-科、属-科丰富度之间均呈显著正相关,反映了蚤类科、属、种之间水平分布中呈现的协同变化关系。这与其他地区的研究结论是相一致的(Balmfordetal.,2000;Gaston,2000b;连宏宇等,2009)。蚤类沿海拔梯度的垂直分布中,虽然种-属和属-科之间也呈现了类似水平分布的显著正相关,但种-科丰富度之间却没规律和相关性不显著。说明横断山区地理环境(海拔或纬度)的差异对蚤类丰富度之间的数量关系存在一定影响,此影响在海拔梯度分布中尤为明显。
蚤类种-属-科丰富度之间关系不论在纬度梯度还是在海拔梯度中,其种-属的数量相关要强于种-科的数量相关。这与我国动物地理亚区蚤类(连宏宇等,2009)或其他的研究(McAllisteretal.,1994;Balmfordetal.,2000;Olsgardetal.,2003;冯永军等,2006;郑孜文等,2008),种与属的数量相关明显大于物种与科的数量相关的结果基本一致。这种关系在鸟、宿主动物及其寄生虫的物种预测或生物多样性评价与保护方面有着重要意义。
3.2 物种、属、科多样性空间分布特征与重要环境因素的关系
种、属和科是人为划分的单元,但是属、科划分的标准及地位的确定是来自于客观的物种特征,而属和科较为真实了反映了物种间亲缘关系远近、性状和生态特征相似性大小(Mayr,1953;Gaston,2000b)。种和属、科之间的数量关系应有着内在的机制在起作用,而非仅仅是简单的数量关系,同时还反映了生物与生物之间、生物与地理和气候环境之间综合作用的结果。
云南横断山区,蚤类科、属、种丰富度,特有种丰富度和特有度,以及东洋界和古北界区系成分物种丰富度都呈现了随纬度或海拔的增加先增高后降低的单峰分布格局,最大峰值出现在北纬25°~27°N和海拔2 500~3 800 m之间,形成物种密度高峰的主要因素是两大动植物区系的分界线和交错区边缘效应的影响(龚正达等,2005,2007)。在横断山区所知蚤类9科45属约153种(亚种)中,有102种为横断山所特有,特有度高达66.67%(龚正达等,2007)。其中,在中纬度梯度(25°~27°N)和中海拔梯度(海拔2 500~3 800 m),特有度分别高达约65%(63%~68%)和 75%(72.58%~75.32%),特有种的组成及分布在当地蚤类物种水平和垂直分布格局的形成中占有举足轻重的地位(龚正达等,2005,2007)。由此认为,两大动植物区系的分界线和交错区边缘效应的影响首对蚤类物种的分化和形成关系最大、其次为属丰富度、而对科丰富度形成不明显。此现象尤其在蚤类垂直分布中尤为明显,并形成了蚤类种-科丰富度之间相关性低于种-属或属-科丰富度之间相关性的特征。
3.3 区域物种多样性与局域物种多样性
横断山区(局部地区)蚤类丰富度(γ多样性)之间相关性基本呈现为显著正相关,与我国(区域)蚤类物种与属以上阶元在数量上具强相关关系(连宏宇等,2009)结果一致。但各山峰(局域)蚤类丰富度之间相关性、物种多样性指数(α多样性)均不同,而且蚤类丰富度相关性与物种多样性指数之间关系均不显著(表4,5)。由于空间范围和尺度上的差异,前者反映了横断山区(区域)物种多样性(γ多样性:物种的分布状况和多样化)的空间分布规律与特征;而后者则反映了横断山区内不同山峰局域(α多样性:局域、群落的组织水平)物种多样性特征。既往研究证实,形成和影响横断山区两者蚤类物种多样性的重要环境因素显然不同:前者主要是受两大动、植物区系分界线和交错区的边缘效应(龚正达等,2005; 2001b; 2007);而后者主要取决于该区域雨量空间分布的特征(龚正达等,2001a)。
由于空间尺度、物种多样性测度和地理环境等因素之间存在差异,在应用高级分类阶元代替物种作为生物多样性的度量单位来进行中、小尺度的热点地区、局域(或自然保护区)的选择、评价及其可行性等可能会存在一定的问题。因此,应引起研究或应用者的注意。
3.4 蚤类属以上阶元所含物种数的频次分布
我国蚤类物种与属以上阶元在数量上具强相关关系(连宏宇等,2009),这种关系在横断山区研究中得到了进一步证实和具有一定的意义。但是,横断山区与我国蚤类属以上阶元所含物种数之间存在明显的差异。横断山区已知蚤类9科45属约153种和亚种,与我国已知蚤类10科74属650余种和亚种(吴厚永等,2007)的构成分别相比,科占90%(9 /10),属约占60%(2/3),而种仅约占24%(约1/4),即横断山区蚤类科数与我国几乎相等的情况下,其属和种丰富度构成比明显较低。此外,前者科、属均含物种数分别为1∶17、1∶3.4,而后者高达1∶65、1∶8.8。它反映出横断山区(局部地区)与我国(区域)空间尺度不同,形成的蚤类属以上阶元所含物种数或频次分布之间存在明显差异。
云南由于地理和气候环境的差异, 5个不同自然地理小区蚤类在科的数量(6或7个)变化不大的情况下,横断山中部小区蚤类达41属和119种(属、种比例为1∶2.9),而滇南山地小区仅有19属24种(属、种比例为1∶1.3)(解宝琦和曾静凡,2000),小区之间属均包含的物种数差异也很大。在不同海拔垂直梯度带中,属以上阶元所含物种数差异更大,其中尤以中海拔梯度的情况较为突出(龚正达等,2005)。显示出云南蚤类区系中,各小区蚤类在科数量变化不大的情况下,而属和种数量,尤其是种的数量差异很大。这也反映了当地各小区蚤类科、属、种之间并非呈简单正比关系的区域特征,并提示了地理环境因素对蚤类多样性空间分布特征具有重要影响。
因此认为,利用高级分类阶元代替物种作为生物多样性的度量单位,进行不同区域的评价和运用中也可能存在一定的问题。
目前,物种和高级分类阶元在数量上具强相关关系,已在许多生物类群(如植物、兽类、鸟类、两栖类、鱼类、蚤类等)上被报道。然而,对物种和高级分类阶元的数量相关机理的探讨较少和在实际研究中却很少频次关系的相关报道(胡军华等,2007)。因此,在横断山区特定区域和复杂的地理环境条件下,蚤类与主要宿主动物不同类群间物种-属-科丰富度之间的关系、他们属以上阶元所含物种数的频次分布规律及其空间分布特征、不同地理和气候环境因素,尤其是两大动、植物区系交错区的边缘效应对它们的影响及其变化趋势以及与疾病的关系等,都是值得进一步研究和探讨的问题。
猜你喜欢
云南社会科学(2021年4期)2021-08-14
经济研究导刊(2019年22期)2019-09-17
齐鲁周刊(2017年29期)2017-08-08
中国学术期刊文摘(2016年1期)2016-02-13
中国医学装备(2015年10期)2015-12-29
太阳能(2015年12期)2015-04-12
电测与仪表(2015年7期)2015-04-09
四川师范大学学报(自然科学版)(2015年4期)2015-02-28
西安建筑科技大学学报(自然科学版)(2014年1期)2014-11-12
测绘学报(2014年2期)2014-01-11
