
阿坝牧区草地不同利用方式与程度对植被碳含量的影响
2014-11-12 02:08杨树晶李涛干友民王永纪磊宋中齐刘焘
草业学报 2014年3期
杨树晶,李涛,干友民*,王永,纪磊,宋中齐,刘焘
(1.四川农业大学草业科学系,四川 雅安625014;2.西南民族大学生命科学与技术学院,四川 成都610065)
草地生态系统是全球生态系统中必不可少的构成成分,对研究全球碳循环过程具有重要作用[1-3]。草地生态系统的碳含量主要由植被碳含量和土壤碳含量组成,而植被碳含量包括地上生物和地下生物碳含量[4]。有研究表明,不同的利用方式对草地植被的碳含量影响极大,放牧草地和人工草地的植被碳含量显著小于原生草地[5]。放牧是人类干扰草地生态系统的主要方式。放牧不仅会降低草地地上生物量,改变草地群落组成,而且影响草地植物碳含量[6],由于草地补偿生长现象的存在,草地初级生产力并非随着放牧强度的增加而线性下降[7],但是过度放牧减少了碳素由植被向土壤中的输入,降低草地的固碳能力[8]。
阿坝州位于青藏高原东南缘,草地类型以亚高山草甸为主,其独有的气候环境,使得冻土对热力极其敏感,具有很大的碳排放潜力,从而直接影响草地生态系统的碳循环[4]。国内外学者自20世纪相继对高寒草甸植被碳含量进行研究[9],曾永年等[10]研究高寒草地碳含量发现,在自然扰动和人为因素的影响下,降低了草地植被生产力,并加快了土壤有机质分解速率。草地植被碳含量受到气候、土壤特征、植被群落结构和人为干扰等综合因素的影响[8]。目前的研究多集中于地上植被和土壤,缺乏对草地枯落物和地下根系碳含量的深入研究[9-10],尤其是凋落物碳含量的研究更是少见。因此,系统地研究草地生态系统碳含量,解决草地生态系统是源还是汇这一难题,有助于增加对全球碳平衡与草地生态系统碳循环的了解,对研究青藏高原区域草地碳含量具有重要的理论价值与现实的生态意义。本研究旨在通过对草地地上植被、枯落物和地下根系三方面的比较和分析,揭示人为干扰对高寒草甸植物碳含量变化的影响,探讨草地植被碳含量对不同利用方式和生长季不同利用程度的响应,从而评估草地植被碳含量在草地生态系统中的价值和贡献,对高寒草地的生态保护、合理利用和植被恢复具有重要的指导意义,并为全球草地生态系统的碳循环研究提供科学依据。
1 材料与方法
1.1 研究区域概况
试验区域位于阿坝州 (30°35′~34°19′N,100°30′~104°27′E)红原、若尔盖、阿坝牧区县,海拔3442~3789 m,地势高平,年降水量500~800 mm,年均温-2~5℃,属于高寒半湿润和湿润地区[11]。土壤主要为亚高山草甸土,有机质含量丰富,无盐碱化。草地植被主要是亚高山草甸。
1.2 试验方法
按照利用方式将阿坝牧区的草地分为封育天然割草地、人工栽培草地、冬春放牧草地和夏秋放牧草地;根据草地植被地上生物量、牦牛的理论采食量、草地面积及在植物生长季利用程度大小等将草地分为冬春放牧草地(夏秋利用率为0)、夏秋轻度放牧(利用率为30%)、夏秋中度放牧(利用率为50%)、夏秋重度放牧(利用率为70%)4个强度[12-13]。2011年7-8月,依据草地植被类型和土壤类型相似等特征选取18个试验样地(表1),进行野外取样及室内测定。
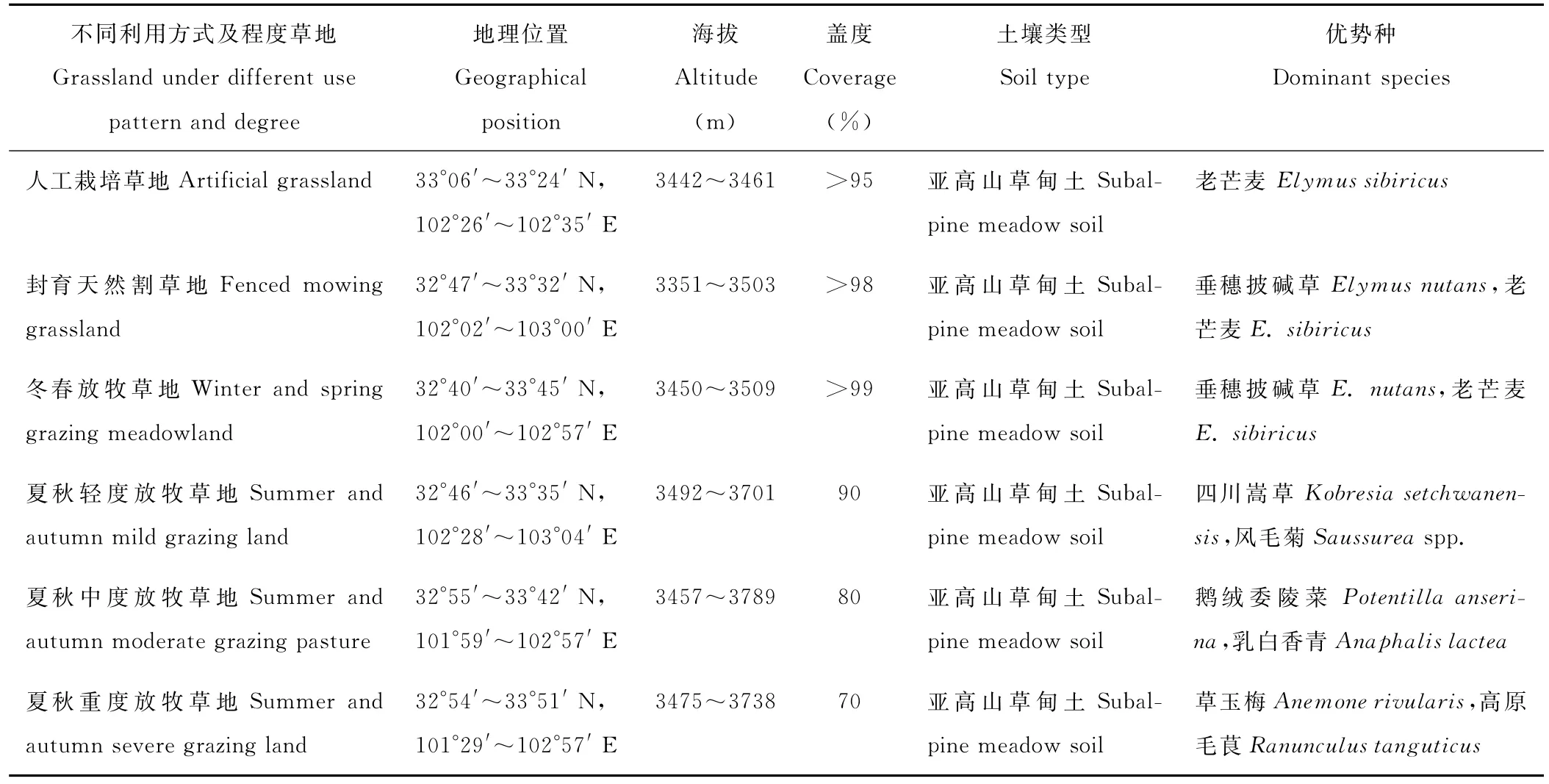
表1 不同利用方式与程度草地样地概况Table 1 The basic situations of every sampling site of grassland under different use pattern and degree
在每个样地选取100 m×100 m区域进行取样调查,在其对角线上按照10 m等距设置1 m×1 m的样方10个。将地面以上的所有绿色部分用剪刀齐地面剪下后装袋标记为地上生物量,65℃烘干待测;用手将地表的凋落物捡起并去掉上面附着的细土粒,装入信封内标记;将收集过地上生物量的样方土壤表面的残留物和杂质清理干净,采取根钻法用7 cm直径根钻分别取3钻混合,分成0~5,5~10,10~20,20~30,30~50,50~70,70~100 cm七层,依次取植被地下生物量,带回实验室冲洗至无杂质,65℃烘干至恒重,具体试验操作参照中国科学院战略先导科技专项(XDA05050404-1-2)调查规范。用K2Cr2O7-H2SO4氧化-外加热容量法测定地上及地下生物量的有机碳含量[14]。
1.3 数据分析
使用Excel 2003和SPSS 17.0进行统计分析,选用最小显著差异法(LSD)进行多重比较,显著水平为P<0.05。采用国际通用的转换率0.45将生物量统一以碳(g/m2)的形式表示[15]。
2 结果与分析
2.1 草地地上植被碳含量特征
从表2可以看出,地上植被碳含量在不同利用方式上表现为封育天然割草地(331.781 g/m2)、冬春放牧草地(278.305 g/m2)和人工栽培草地(143.746 g/m2)显著高于夏秋放牧草地(112.871 g/m2);生长季不同利用程度上冬春放牧草地(278.305 g/m2)和夏秋轻度放牧草地(191.983 g/m2)显著高于重度放牧草地(90.712 g/m2)和中度放牧草地(55.916 g/m2)。
2.2 草地枯落物碳含量特征
草地枯落物碳含量在不同利用方式上表现为人工栽培草地(5.525 g/m2)>冬春放牧草地(3.228 g/m2)>封育天然割草地(3.091 g/m2)>夏秋放牧草地(2.772 g/m2);生长季不同利用程度上冬春放牧草地(3.228 g/m2)>夏秋中度放牧草地(2.953 g/m2)>夏秋轻度放牧草地(2.787 g/m2)>夏秋重度放牧草地(2.576 g/m2)。差异均不显著(P>0.05)(表2)。

表2 不同利用方式与程度草地植被碳含量Table 2 Vegetation carbon storage of grassland under different use pattern and degree
2.3 草地各层根系碳含量特征
从根系总碳含量来看(表2),不同利用方式上夏秋放牧草地(21.935 g/m2)和封育天然割草地(21.637 g/m2)显著高于冬春放牧草地(16.247 g/m2)和人工栽培草地(15.682 g/m2);生长季不同利用程度上夏秋中度放牧草地(23.012 g/m2)和夏秋轻度放牧草地(22.232 g/m2)显著高于夏秋重度放牧草地(20.561 g/m2)和冬春放牧草地(16.247 g/m2)(P<0.05)。
阿坝牧区不同利用方式与程度草地根系碳含量较一致地表现为从表层向下依次显著的递减。0~10 cm的根系碳含量占总量的65%以上,其中封育天然割草地和夏秋放牧草地的根系碳含量占总量的70%以上(表3)。
30 cm以下土层地下根系生物量很少,尤其是50 cm以下,几乎没有。30~100 cm总碳含量在0.43~0.95 g/m2之间。
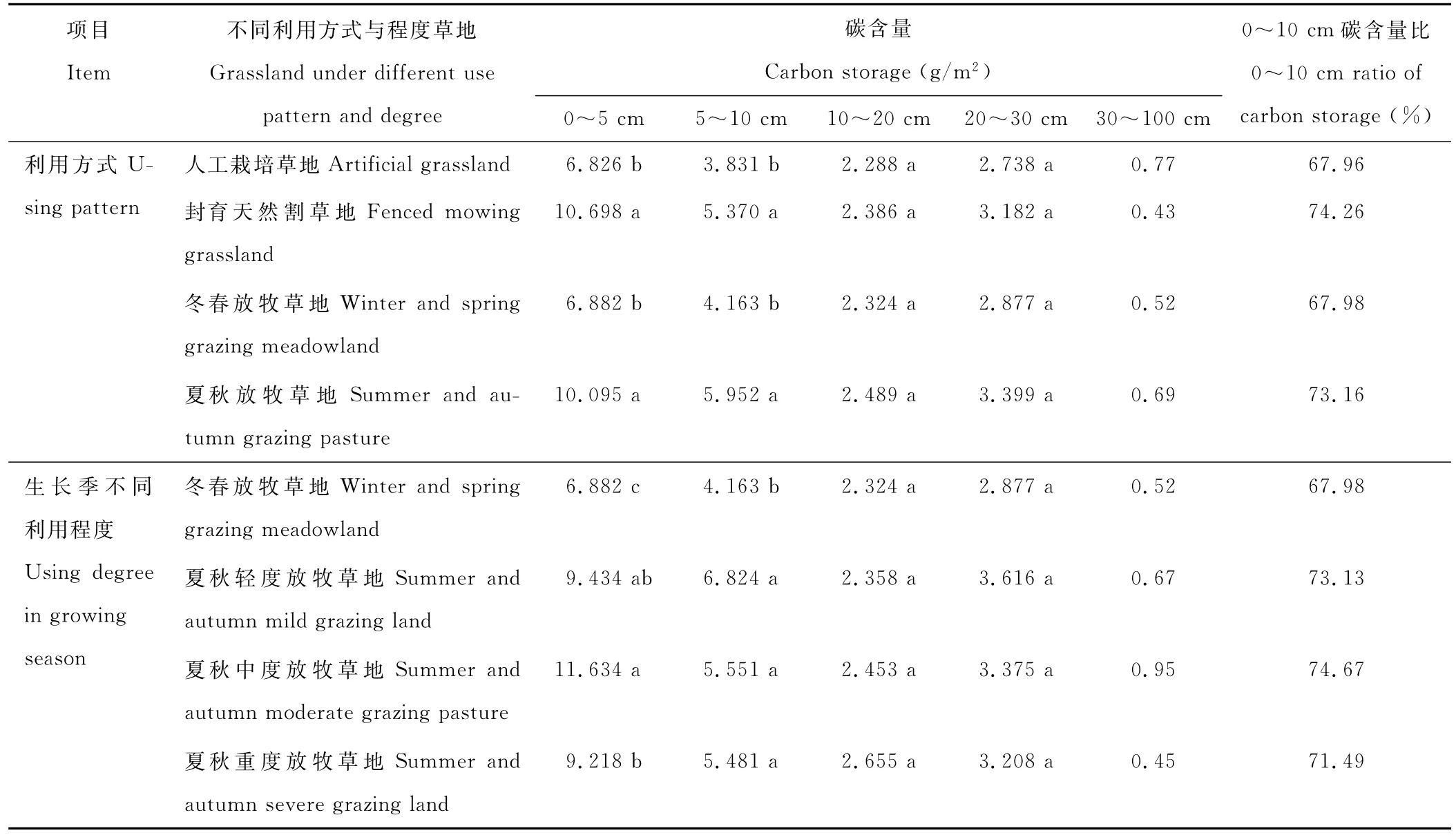
表3 不同利用方式与程度草地各层根系碳含量Table 3 Root carbon storage at various layers of grassland under different use pattern and degree
2.4 草地同层根系碳含量特征
2.4.1 草地0~5 cm根系碳含量特征 0~5 cm地下根系碳含量在不同利用方式上表现为封育天然割草地(10.698 g/m2)和夏秋放牧草地(10.095 g/m2)显著高于冬春放牧草地(6.882 g/m2)和人工栽培草地(6.836 g/m2)(表3);在生长季不同利用程度上夏秋中度放牧草地(11.634 g/m2)和夏秋轻度放牧草地(9.434 g/m2)显著高于夏秋重度放牧草地(9.218 g/m2)和冬春放牧草地(6.882 g/m2)。
2.4.2 草地5~10 cm根系碳含量特征 如表3所示,5~10 cm根系碳含量在不同利用方式上表现为夏秋放牧草地(5.952 g/m2)和封育天然割草地(5.370 g/m2)显著高于冬春放牧草地(4.163 g/m2)和人工栽培草地(3.831 g/m2);在生长季不同利用程度上冬春放牧草地(4.163 g/m2)显著低于夏秋轻度放牧草地(6.824 g/m2)、夏秋中度放牧草地(5.551 g/m2)和夏秋重度放牧草地(5.481 g/m2)。
2.4.3 草地10~20 cm根系碳含量特征 在10~20 cm地下根系碳含量中,不同利用方式表现为夏秋放牧草地(2.489 g/m2)>封育天然割草地(2.386 g/m2)>冬春放牧草地(2.324 g/m2)>人工栽培草地(2.288 g/m2);生长季不同利用程度上夏秋重度放牧草地(2.655 g/m2)>夏秋中度放牧草地(2.453 g/m2)>夏秋轻度放牧草地(2.358 g/m2)>冬春放牧草地(2.324 g/m2),差异均不显著(P<0.05)(表3)。
2.4.4 草地20~30 cm根系碳含量特征 在20~30 cm地下根系碳含量中,不同利用方式表现为夏秋放牧草地(3.399 g/m2)>封育天然割草地(3.182 g/m2)>冬春放牧草地(2.877 g/m2)>人工栽培草地(2.738 g/m2);生长季不同利用程度上夏秋轻度放牧草地(3.616 g/m2)>夏秋中度放牧草地(3.375 g/m2)>夏秋重度放牧草地(3.208 g/m2)>冬春放牧草地(2.877 g/m2),差异均不显著(P<0.05)(表3)。
2.5 草地植被碳含量数量关系
草地植被碳含量在利用方式上表现为封育天然割草地(356.509 g/m2)>冬春放牧草地(297.780 g/m2)>人工栽培草地(164.953 g/m2)>夏秋放牧草地(137.577 g/m2),地上植被碳含量分别为地下根系碳含量的15.334,17.130,9.167和5.146倍(表2);在生长季不同利用程度上表现为冬春放牧草地(297.780 g/m2)>夏秋轻度放牧草地(217.002 g/m2)>夏秋重度放牧草地(113.849 g/m2)>夏秋中度放牧草地(81.882 g/m2),地上植被碳含量分别为地下根系碳含量的17.130,8.636,4.412和2.430倍。
3 讨论
曹广民等[16]发现高寒草地植物生长季(5-9月)CO2的排放量大约为非生长季(10-翌年4月)的2~3倍,因此,在植物生长季研究草地植被碳含量意义重大。植被作为草地生态系统碳含量的重要组成部分,包括地上植被、枯落物和地下根系碳含量3个部分[17]。草地地上植被的生物量与碳含量息息相关;植物茎叶脱落于地表形成枯落物,其中一部分碳素经过微生物分解作用释放到大气中,另一部分碳素以微生物的形式被固定在土壤中[18],是连接土壤碳库和植被碳库的重要部分[19];地下根系作为吸收和贮藏营养的重要器官,其生物量的多少及其分布直接影响地上植被的生长状况,同时,也是土壤有机碳的重要来源[16,20]。
草地利用方式的不同,不仅会引起地上植被的变化,而且影响地下根系的分布[21]。由于长期的封育不用,草地原生植被得以恢复和生长,植物群落丰富度和多样性大[21-23],优良禾本科牧草的优势地位提高,地上生物量和地下根系都增加,草地枯草层厚,枯落物多,因此,封育天然割草地的地上植被、枯落物和地下根系碳含量都相对较高。冬春放牧草地在整个植物生长季未被利用,冬春季家畜的采食,降低了地上植被和枯落物的生物量,因此,冬春放牧草地的植被碳含量要小于封育草地。通过开垦而建立的人工栽培草地,由于原生植被被取代后,碳素向地下根系和土壤中的分配比例降低[24-25],以收获和高产为目的收割又减少了地上生物量中碳素的累积,且其植被组成单一,植株相对较高,但是植物根系所占比例减小,导致了人工草地的植被碳含量小于封育天然割草地和冬春放牧草地。夏秋放牧地由于在生长季进行放牧,地上植被和枯落物被家畜采食大幅度减小,而植被为保证物种延续,将资源先向根系转移[26],虽然地上植被和枯落物碳含量小于其他3种利用方式的草地,但是根系碳含量最高。在本次试验样地中,封育天然草地和冬春放牧草地以原生垂穗披碱草(和老芒麦)等为优势种,地上植被和地下根系生物量都相对较高;人工栽培草地则以收获种子的老芒麦为主,虽然具有较高的地上生物量,但是植物种类单一,地下根系很小;夏秋放牧地则以原生的莎草科和阔叶类植物为优势种,虽然地上植被和枯落物生物量相对较小,但是地下根系相对发达。由于不同的优势种对草地植被碳含量的贡献率的差异[8],草地植被碳含量表现为封育天然割草地>冬春放牧草地>人工栽培草地>夏秋放牧草地,这一结论与乔友明等[21]及韩道瑞等[5]的研究结果一致。0~10 cm为草根层,植被地下生物量大量聚集于此,微生物活动活跃,加大了碳循环作用,将大气碳素固定转化为植被中含碳化合物,因此碳含量大;而随着深度增加,10~30 cm地下生物量越来越少,植被固定和转化外界碳能力减弱,所以其碳含量相应较小[20],该深度草地植被根系碳含量在不同土层深度上的差别很小,说明草地从地上部分枯落物向根系转移碳含量的功能减弱,其原因可能是草地的开垦和放牧破坏了地上植被,使得草地根系生物量以及枯落物的积累量都明显减少,最终导致根系碳含量的下降[5]。
过度放牧是我国草地退化的主要原因[27]。放牧会降低群落优势种的优势度,使草地生态系统的群落组成发生变化。在轻度和适度的放牧下,其植被盖度、物种丰富度和生物量都相对较大,因而将碳素固定和转化到植被的效率就较高,固定的碳量也就相对较多[28-29],草地重度放牧草地则正好相反,长期的过度放牧还会导致草地初级生产力固定碳素的能力降低,家畜采食减少了碳素由植物凋落物向地下的输入[8]。冬春放牧草地在整个生长季植被得以恢复,只有冬春季进行放牧,在植物生长季受到的干扰最小,是植物生长季利用程度最轻的草地,由于地上生物量和枯落物生物量较高,根系相对于夏秋放牧地却很小,说明草地不同季节的利用对植被碳含量的影响较大[16]。不同放牧强度明显改变了植物有机碳的地上/地下分配比,随着放牧压力的增大,草地植物群落植物先将有机碳向地下转移[26],但是本试验结果发现,地下根系生物量随着放牧强度增强而降低,夏秋重度放牧草地地下根系碳含量小于夏秋中度放牧草地,与这一理论结果不一致[26],王启基等[30]的研究成果也表明重度放牧草地植物根系碳含量明显小于轻度和中度放牧草地,这可能与植被群落组成不同及植被中C4植被根系较浅等因素有关[26]。本研究中,重度放牧草地的植被碳含量要高于中度放牧,可能是重牧对草地土壤理化性质和微生物的活动产生大的影响,导致地上植被向地下根系转移的碳素减少,也可能由于重牧改变了植被群落结构,毒杂草的优势地位增大或成为优势种,继而引起植被碳含量贡献率发生差异[7]。
植物凋落物和根系分泌物是形成土壤碳的重要来源,王启基等[30]在研究高山嵩草(Kobresia pygmaea)草甸碳含量时发现土壤碳含量与植物根系呈显著的正相关,而草地地下根系碳含量远远大于植被碳含量[31]。在本试验的结果中,不同的利用方式及程度草地地上植被碳含量是地下根系碳含量的数倍,究其原因有以下两点:试验样地植被物种丰富,优势种有禾本科、莎草科和阔叶类植物,且草地植被盖度在70%以上或大于90%,导致地上植被碳含量要远高于高山嵩草草甸和干旱草甸;本试验根系生物量采用根钻法,可能导致所测得的地下根系碳含量有异于挖掘法[20]。
4 结论
不同的利用方式对草地植被碳含量影响显著,封育天然割草地植被碳含量高于冬春放牧草地和人工栽培草地,这表明高寒草地通过围栏封育、植被恢复及提高植物多样性可增加草地植被的碳含量。人为干扰程度是影响草地植被碳含量的重要因素,草地季节轮换放牧、合理的放牧强度均增加草地碳含量,重度放牧则有可能促进草地从碳汇向碳源的转变,且草地地上植被碳含量高于地下根系碳含量,虽然草地生态系统中绝大部分的碳贮藏在土壤中,但是草地植被碳含量也是草地生态系统碳循环的重要组成部分,制定合理的草地放牧策略更有利于高寒草地植被的保护和利用,如何确定人类活动对草地植被合理的干扰强度指标,还需进一步研究。
猜你喜欢
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
绿色科技(2021年10期)2021-06-23
今日农业(2020年19期)2020-12-14
电脑报(2020年12期)2020-06-30
草业学报(2019年2期)2019-02-25
安徽医科大学学报(2016年12期)2017-01-15
癌症进展(2016年9期)2016-08-22
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
