
不同地理区域桔小实蝇解毒酶系活性及其与抗药性水平关系
2014-12-09 09:14梁广文陆永跃
环境昆虫学报 2014年1期
金 涛,梁广文,曾 玲*,陆永跃*
(1.华南农业大学昆虫生态研究室,广州 510642;2.中国热带农业科学院环境与植物保护研究所,海口 571101)
桔小实蝇Bactrocera dorsalis (Hendel),属双翅目Diptera,实蝇科Tephritidae,寡毛实蝇亚科Dacinae,是我国和许多国家重要害虫。桔小实蝇生活周期短、繁殖力强,寄主范围广,可为害番石榴、杨桃、芒果等46 个科250 多种水果、蔬菜和花卉,是一种危害性大的害虫(Bateman,1972;Allwood et al.,1999)。近年来,在我国发现该虫有不断向长江流域地区扩散的趋势 (范京安,1998;蒋小龙等,2001;侯柏华和张润杰,2005;熊焰等,2006)。目前防治桔小实蝇还是以化学农药为主,辅以引诱剂和水果套袋技术。田间持续的化学防控已使得桔小实蝇抗药性不断增强(潘志萍等,2005;章玉苹等,2007;章玉苹等,2008;Jin et al.,2011)。室内培育不同抗药性品系的研究发现,桔小实蝇对敌百虫、高效氯氰菊酯和阿维菌素抗性发展较快,汰选至14 代其抗性分别增长至84.16 倍、27.1 倍和7.7 倍(潘志萍等,2008)。对实验室内已形成的对敌百虫产生桔小实蝇高抗品系衰退再增长的趋势研究表明,桔小实蝇通过6至9 个世代的选育,抗性会迅速上升至原来水平(章玉苹等,2008)。抗性增强导致农药使用效率下降,加大了防治难度。对桔小实蝇抗药性机制的研究有助于抗药性治理和制定有效的防治策略,延缓抗性水平的上升。
昆虫的抗药性机制大致可分为行为躲避、代谢解毒能力增强和靶标不敏感等类型。相关研究表明,与杀虫剂代谢相关的解毒酶活性的增强是害虫对多种杀虫剂产生抗性的主要生理标志(刘泽文等,2002;张红英等,2002;Liu,2003;张咏梅等,2006;刘永杰等,2007;van Pottelberge et al.,2008)。昆虫解毒酶是一类异质酶系,能够结合代谢大量的内源或外源底物,主要包括多功能氧化酶、羧酸酯酶和谷胱甘肽S-转移酶等,不同的解毒酶对不同杀虫剂的抗性发展作用不同(Oppenoorth,1965;Terriere,1984;刘泽文等,2002;张红英等,2002)。其中,MFO 的底物谱极广,几乎能氧化代谢所有的杀虫剂,与很多的害虫抗药性形成有关(周成理等,1993;陈之浩等,1993)。CarE 是昆虫体内重要的几种酶之一,其高亲和性-低能解毒作用的活力增强是昆虫对有机磷和氨基甲酸酯类杀虫剂产生抗性的重要机制(何林等,2003)。GST 是昆虫体内的一类与抗性有关的初级代谢及次级代谢酶系。该酶系能使有害的亲电物质与内源的还原型谷胱甘肽结合,参与转运体内重要的脂类化合物,能为谷胱甘肽的S原子提供电子催化亲核反应。该酶在一些昆虫对有机磷类、氨基甲酸酯类和有机氯类等传统杀虫剂的抗性中所起的作用很早以前就得到证实(Oppenoorth,1976)。作为与昆虫抗性水平相关的这几种解毒酶,其活性大小也会与对不同药剂的抗性水平存在联系。本文以具有不同抗性水平的桔小实蝇地理种群品系为材料,在测定了解毒酶的活性和抗性水平的基础之上,比较了这几种酶的活性和抗性水平关系,为寻找抗性监测的生化指标,明确桔小实蝇解毒酶活性与其抗性水平关系,进行有效的桔小实蝇抗药性治理提供科学依据。
1 材料与方法
1.1 材料
1.1.1 供试虫源
相对敏感品系:桔小实蝇采集于广东省广州市杨桃公园。在华南农业大学昆虫生态研究室昆虫饲养室饲养,在室内不接触任何药剂条件下,用人工饲料、香蕉等连续饲养繁殖33 代。饲养环境:温度26-30℃,光照周期L∶D=16∶8 h,相对湿度:75%-90%。药膜法测定该品系对敌百虫、高效氯氰菊酯和阿维菌素抗药性程度为低抗性水平(致死中浓度均小于3 mg/L)。
地理种群品系:选定全国桔小实蝇主要发生为害区域。于2007年至2008年各地区水果成熟季,一次性采集各地区杨桃、番石榴、柑桔等落果,约挑出老熟的3 龄幼虫约300 头,使用人工饲料饲养至下一代,在室内建立地理种群。
1.1.2 主要试剂及仪器
主要试剂:α-乙酸萘酯,化学纯,中国医药集团上海化学试剂公司;α-萘酚,分析纯,上海化学试剂公司;毒扁豆碱,生化试剂,Fluka 公司;对硝基苯酚和对硝基苯甲醚,上海三爱思试剂有限公司;十二烷基硫酸钠(SDS)、固蓝B 盐和考马斯亮蓝G-250,为Sigma 公司产品;还原性辅酶Ⅱ和牛血清蛋白(组分五),Roche 试剂公司。丙酮、无水乙醇、Na2HPO3和NaH2PO3均为国产分析纯。谷胱甘肽-S 转移酶测试试剂盒购于南京建成生物工程研究所。
主要仪器:751 型分光光度计;Centrifuge 5810 冷冻高速离心机;恒温水浴锅。
1.2 抗性水平的测定
从每个测定酶活的桔小实蝇相对敏感种群和地理种群中,选羽化后3-5 d 附肢齐全、行为活泼的成虫供试。采用药膜法测定不同地理种群桔小实蝇对敌百虫(Trichlorphon)、高效氯氰菊酯(β-cypermethrin)和阿维菌素(Avermectin)抗性水平(潘志萍等,2005;Lin et al.,2013)。
应用SPSS 数据处理系统(贾春生,2006)和EXCEL 表格分析软件(张志祥等,2002),计算出的致死中浓度(Lethal Concentration 50,LC50)及其95%置信度,抗性倍数(Resistance Multiple,RM)为供试种群的LC50与相对敏感种群的LC50的比值。
1.3 酶活测定方法
1.3.1 多功能氧化酶(MFO)-O-脱甲基活性测定
越秀说:“我们成亲后,乔瞧姐就很少出门,也不替人看病了,看到她,跟她说话,她就当没听见。她都二十五六了,还没嫁人。”
在室内饲养1代后,从各地理桔小实蝇种群中取5 头羽化后3 至5 天后成虫,置于冰浴的研钵中,加入少量的石英砂研磨后。加入0.1 m mol/L PH7.8 磷酸缓冲液1.2 mL,充分匀浆。在4℃,3600 rpm 离心15 min。取上清液作为酶源备用。每处理重复3 次。测定多功能氧化酶-O-脱甲基活性,参照Hung 等(Hung and Sun,1989)的方法并改进。取0.5 mL 的酶液、1 mL 的1 m mol/L NADPH、0.1 mL 的0.1 m mol/L 对硝基苯甲醚。置于30℃水浴锅中反应30 min,然后加1 mL 浓度为1 mmol/L HCl 终止反应。再向试管中加入5 mL氯仿萃取,分层后在氯仿层移取3 mL 到另一试管内,再加3 mL 的0.5 mol/L NaOH 萃取。取NaOH溶液层2 mL 于比色皿中,于400 nm 处测OD 值,根据对硝基苯酚标准曲线和酶源的蛋白质含量,将OD 值换算成比活力μmol· (mg·30 min)-1。
1.3.2 羧酸酯酶(CarE)活性测定
酶源制备方法基本同上,只是使用0.04 mmol/L PH7.0 磷酸缓冲液代替原来的PH7.8 的磷酸缓冲液作为提取液。参照van Asperen (Vail et al.,1980)的方法。取0.1 mL 的酶液和5 mL 的反应底物(1 mL 的0.03 mol/L α-醋酸萘酯、1 mL 的3×10-4mmol/L 毒扁豆碱以及98 mL 的0.04 m mol/L PH 7.0 的磷酸缓冲液的混合物)混合后,再加入1 mL 显色剂(5%十二烷基硫酸钠溶液和1%的固蓝B 盐以5∶2 比例配制)。混匀后室温静置30 min,600 nm 下测OD 值。
1.3.3 谷胱甘肽-S 转移酶(GST)活性测定
酶源制备方法基本同上,但使用ddH2O 代替磷酸缓冲液。使用南京建成工程研究所研制的谷胱甘肽-S 转移酶测定试剂盒。其原理为GST 具有催化还原谷胱甘肽(GSH)与1-氯-2,4-二硝基苯结合的能力,在一定反应时间内,其活性的高低与反应前后底物浓度的变化呈线性关系。因而通过检测GSH 浓度的高低来反映GST 活力的大小,其GSH 浓度降低的越多则GST 的活力越大。相关操作过程参照试剂盒说明书。
1.3.4 蛋白质含量测定
参照 Bradford 考马斯亮蓝 G-250 方法(Bradford,1976)。以牛血清蛋白为标准。
1.4 相关分析及通径分析方法
以3种测定酶活性测定值与相对敏感品系酶活性平均值的比值和地理品系对3种杀虫剂的抗性水平作相关性分析,并进行通经分析。试验数据使用SAS 9.0 数据处理软件和EXCEL 表格软件分析、处理。
2 结果与分析
2.1 桔小实蝇各地理品系对三种杀虫剂的抗性水平
桔小实蝇各地理品系对敌百虫、高效氯氰菊酯和阿维菌素的抗性情况如表1 所示。广东茂名桔小实蝇种群对敌百虫的抗性水平最高,是相对敏感品系的100.2271 倍,处于高抗水平。广东惠州地区桔小实蝇种群对敌百虫的抗性水平最低,只有相对敏感品系的2.1552 倍。各地区桔小实蝇品系对高效氯氰菊酯的抗性水平在1.0065 到26.0026 倍之间。抗性水平比较发现,部分地区抗性倍数之间存在显著差异,如广州明显高于福州。云南元江桔小实蝇对高效氯氰菊酯抗性水平最低,只有相对敏感品系的1.0065 倍,明显低于其他地区。各地区桔小实蝇对阿维菌素的抗性倍数在2.3353 至29.0688 倍之间,以茂名的最高,南宁的最低。
2.2 桔小实蝇不同品系的几种酶活性
各地区桔小实蝇成虫的MFO-O-脱甲基活性见表2。广州和无锡桔小实蝇体内的酶活性最高,分别为相对敏感品系的1.4782 和1.4641 倍;而茂名品系的酶比活性最低,只有相对敏感品系的0.8649 倍;其它各地区桔小实蝇品系酶活性比值在0.9603 和1.2950 之间。
2.2.2 桔小实蝇不同品系的羧酸酯酶 (CarE)活性
桔小实蝇不同地理品系的成虫CarE 活性测定结果表明广东惠州桔小实蝇种群最高,为相对敏感品系的1.8147 倍;广西南宁的桔小实蝇酶活性最低,只有相对敏感品系的0.9636 倍;其它各地区桔小实蝇品系酶活性与相对敏感品系酶活性比值在1.1018 和1.3995 之间。9 个桔小实蝇的地理种群CarE 活性变化幅度较小,表明了CarE 活性水平较为稳定(表3)。

表1 各地区桔小实蝇品系和相对敏感品系的抗性倍数Table 1 Resistance folds in different populations and sensitive strains of Bactrocera dorsalis
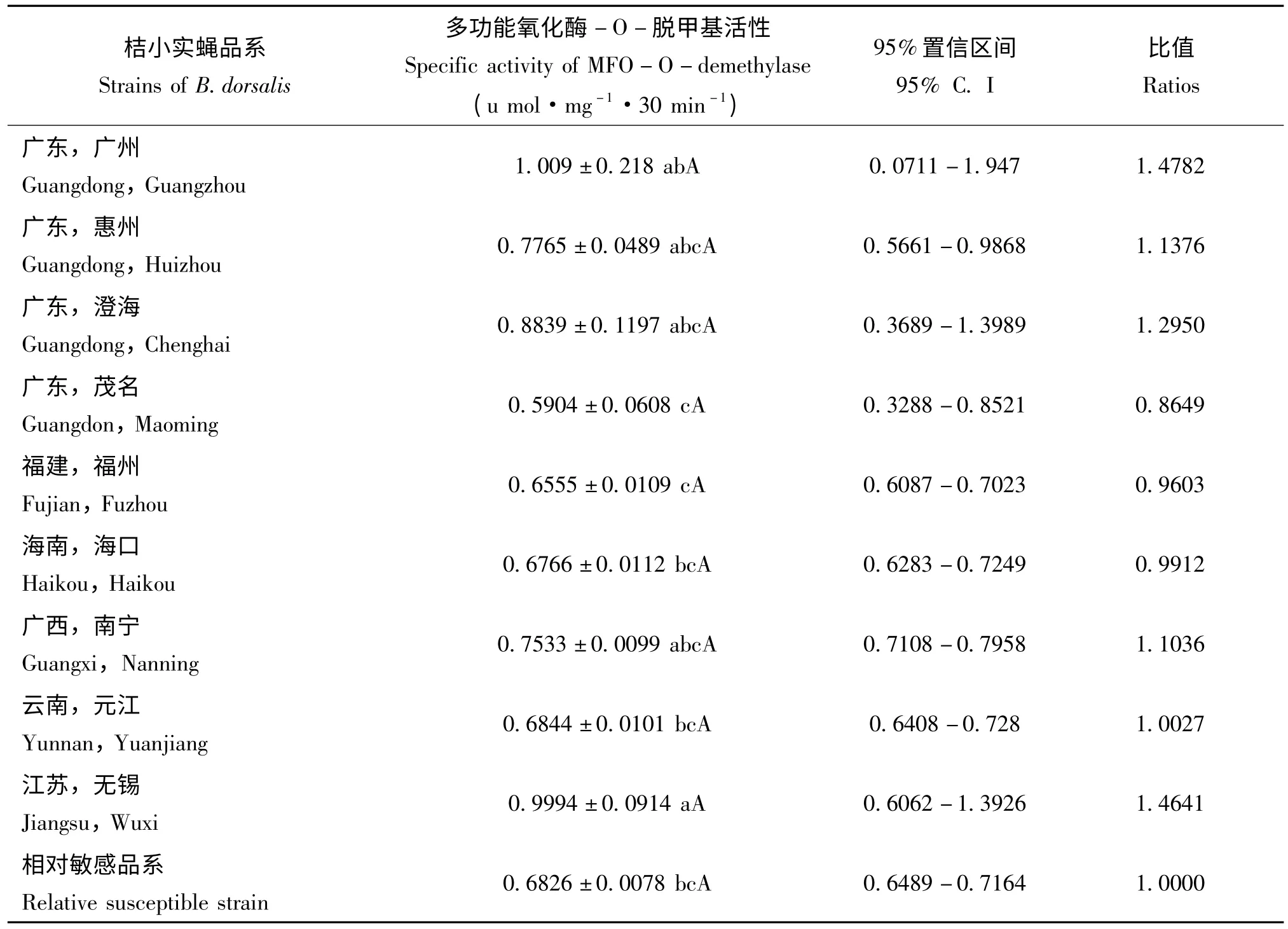
表2 桔小实蝇地理种群的多功能氧化酶-O-脱甲基活性Table 2 Activity of MFO-O-demethylase in different strains of Bactrocera dorsalis
2.2.3 桔小实蝇不同品系的谷胱甘肽-S 转移酶(GST)活性
各地区桔小实蝇成虫的GST 活性测定结果表明广东茂名种群的GST 活性最高,是相对敏感品系的2.2557 倍;江苏无锡和广东澄海种群的GST活性次之,分别为相对敏感品系的2.0283 和2.0102 倍;而广东广州种群活性只有相对敏感品系的1.1622 倍,最低;其它各地区桔小实蝇品系的GST 活性与相对敏感品系的比值在1.2145 至1.9030 之间(表4)。
2.3 桔小实蝇地理品系几种酶活性和抗性水平相关性分析
表5 中相关性分析结果显示,桔小实蝇体内谷胱甘肽-S-转移酶活性和对敌百虫抗性水平相关系数为0.41,达显著性水平;羧酸酯酶活性与桔小实蝇对高效氯氰菊酯抗性水平存在极显著正相关,相关系数为0.50。桔小实蝇对敌百虫和阿维菌素的抗性水平之间也存在极显著关系,其相关系数为0.76。研究结果显示了桔小实蝇几种解毒酶与抗药性水平之间以及对不同的杀虫剂的抗性水平之间存在显著联系,表明了桔小实蝇解毒酶活性对杀虫剂的响应作用是复杂的,不同解毒酶活性对不同杀虫剂的抗性水平相关性程度不同。
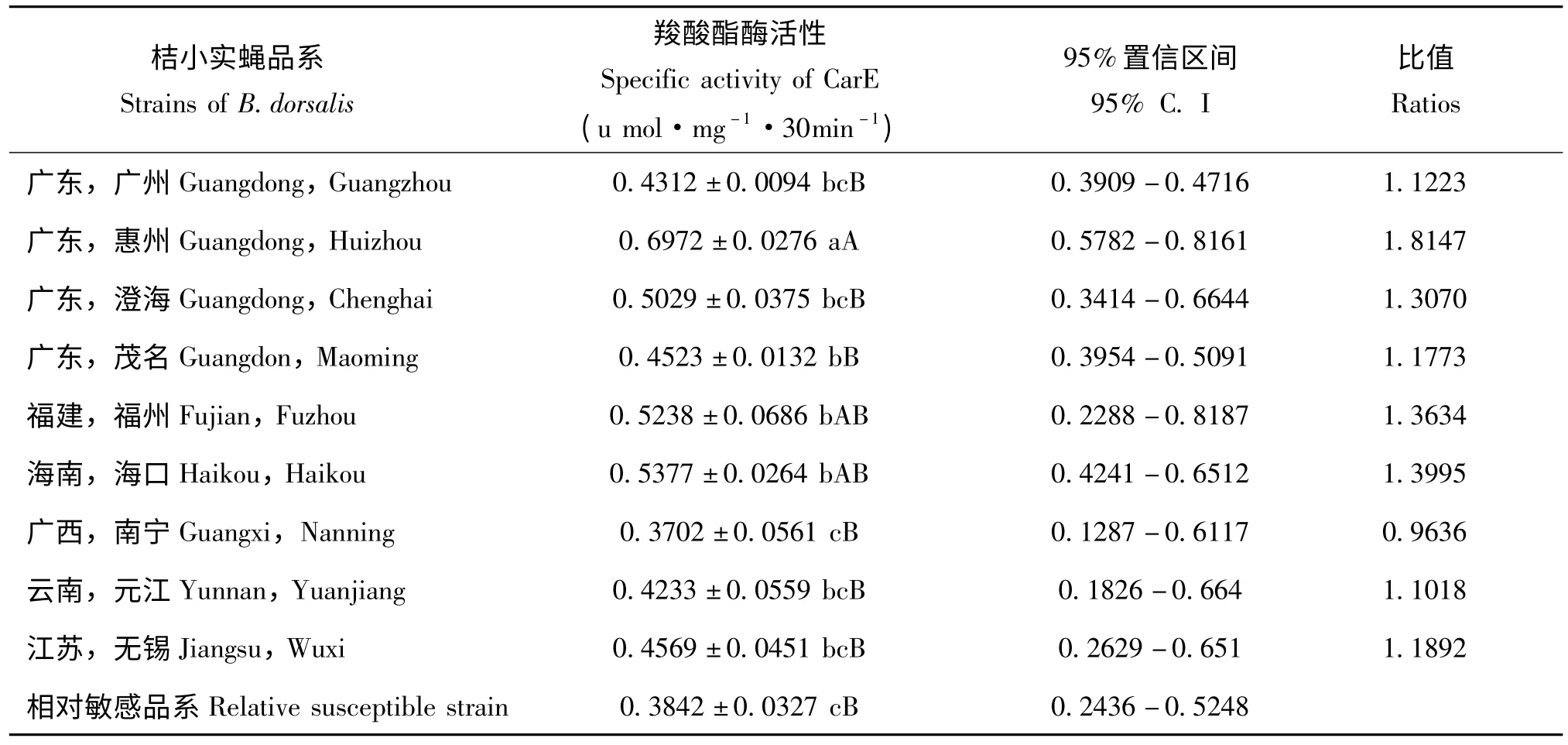
表3 桔小实蝇地理种群的羧酸酯酶活性Table 3 The CarE activity in different strains of Bactrocera dorsalis
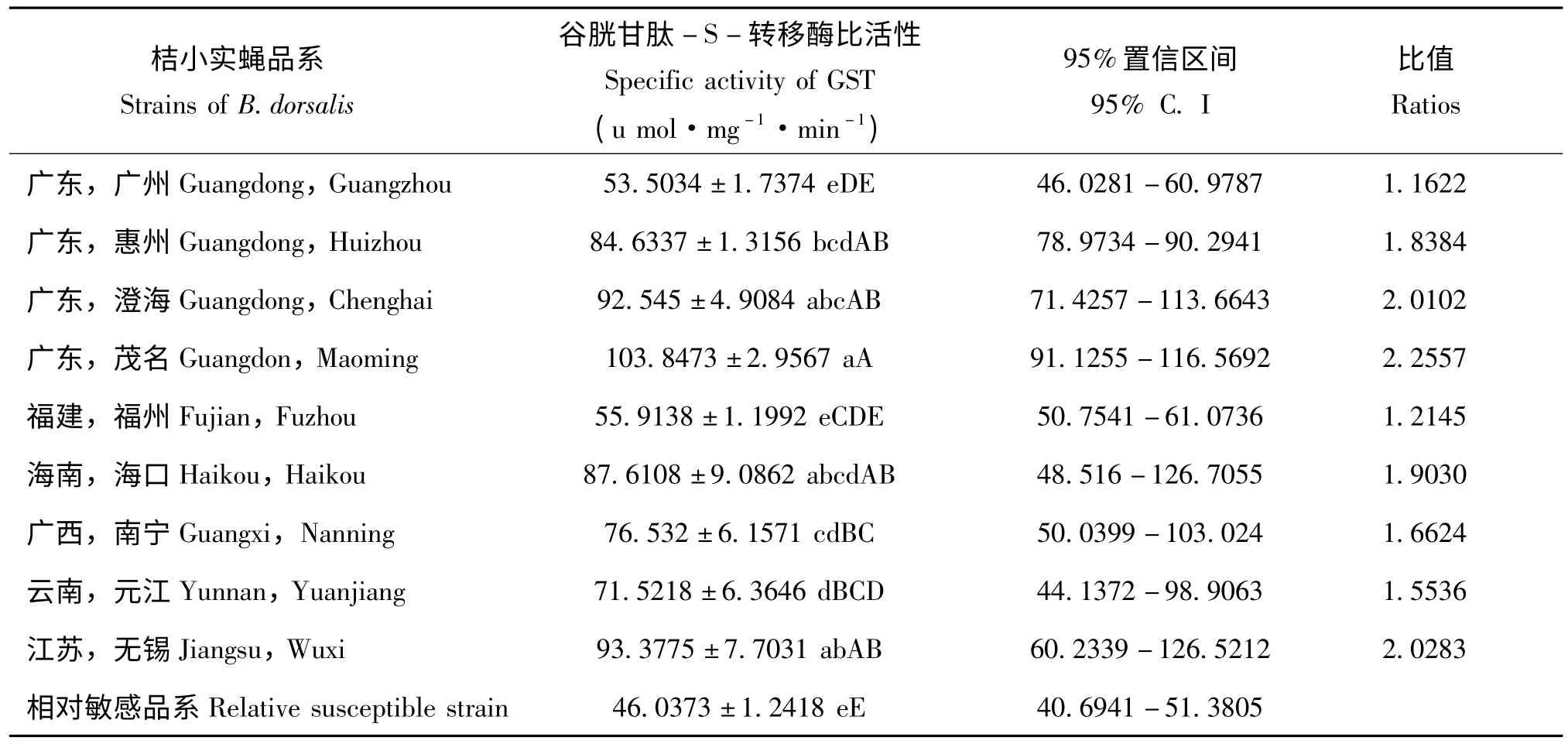
表4 桔小实蝇地理种群的谷胱甘肽-S-转移酶活性Table 4 The GST activity in different strains of Bactrocera dorsalis

表5 几种酶活性比值与各抗药性水平的相关性分析Table 5 The correlative analysis of detoxification enzyme activity ratios and resistance levels
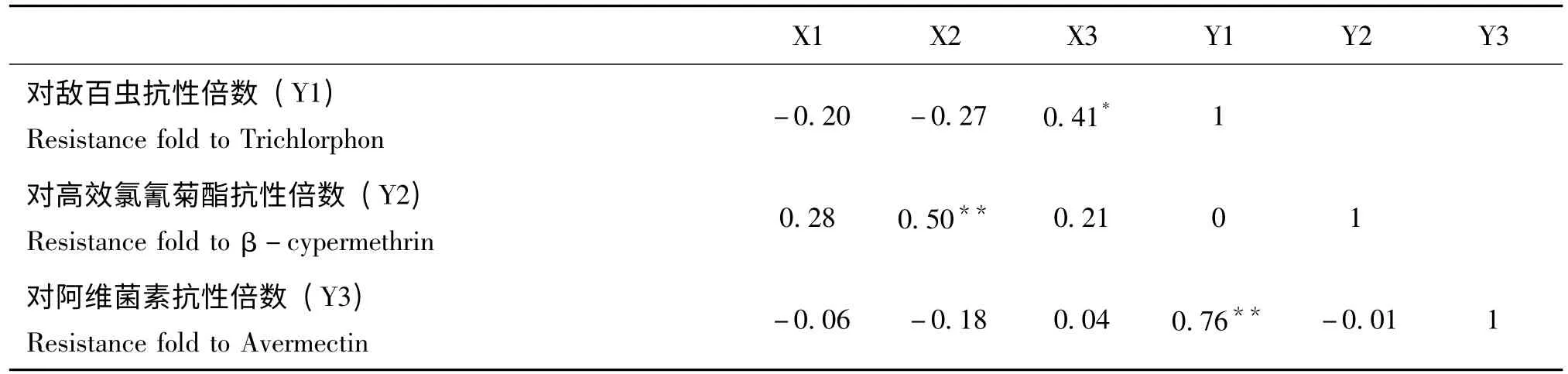
(续上表)
2.4 各几种酶活性与杀虫剂抗性水平的通径分析
通径分析分析结果见表6 和图1。图1 中,除对角线所示的直接通径系数外的数值均为间接通径系数,它表明某一酶活对抗性水平的评分的影响力是通过对其他酶的活性影响而间接影响其抗性水平。其结果表明,GST 活性对敌百虫的抗药性水平的直接通径系数分别为0.4414,起到正向作用;而MFO-O-脱甲基活性和CarE 活性对敌百虫的抗性水平的直接通径系数分别为-0.1768和-0.3531,对抗性水平的均起到负向作用。MFO-O-脱甲基活性、CarE 活性和GST 活性对高效氯氰菊酯抗性水平的直接通径系数为0.3311,0.4946 和0.1775,均起到正向作用。三种解毒酶中,只有GST 活性与阿维菌素的抗性水平的直接通径系数很小,为0.0668;其余2种酶,MFO-O-脱甲基活性和CarE 活性对阿维菌素的直接通径系数均为负值,分别为-0.0608 和-0.1913,对阿维菌素抗性程度起到负向作用。
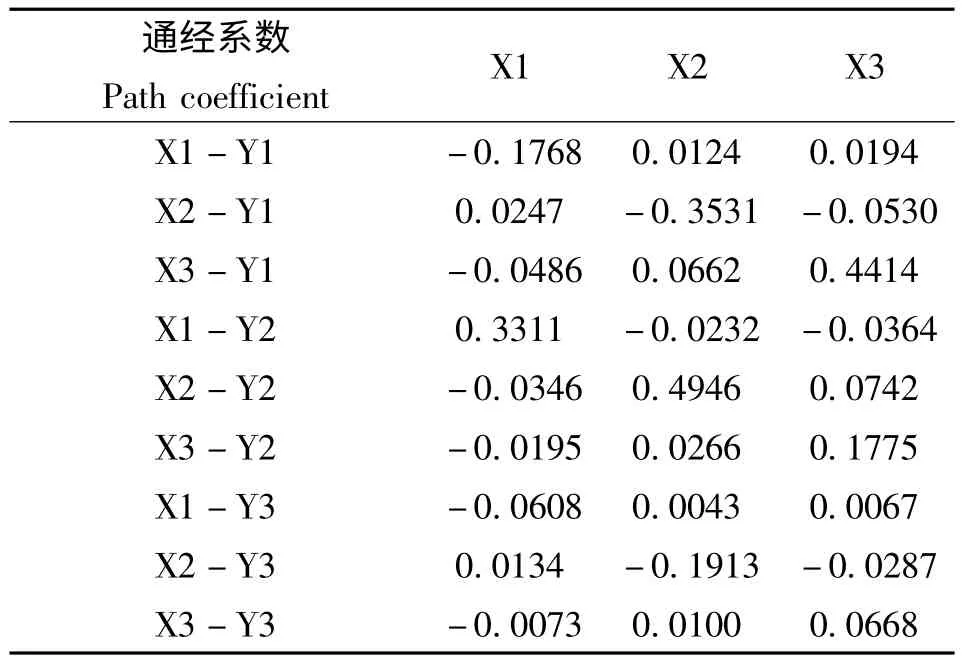
表6 解毒酶活性比值和不同抗药性指标的通径分析Table 6 Path coefficient analysis of each detoxification enzymes activity on resistance level traits
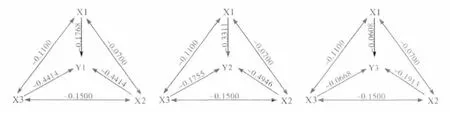
图1 解毒酶活性与抗药性水平的评分通径图Fig.1 The path analysis of detoxification enzymes activity on resistance level
3 结论与讨论
对杀虫剂产生抗药性是害虫的自然适应性特征,是害虫不断的应对杀虫剂而形成的防御性策略。抗性产生的代谢特征主要是解毒作用的增强,生理水平上显然有多种酶系参与,将进入虫体内的药剂经过氧化、还原、水解或结合后,降低其毒性,或增强其水溶性,将其排出体外,达到解毒目的。而各种酶在不同类型的抗药性发展过程中的作用是不同的,本研究的结果表明,桔小实蝇体内解毒酶系的活性变化与抗性水平不存在单一增长关系,而是各种酶在虫体内维持一个活性优化的平衡状态,对不同类型的杀虫剂多种解毒酶的代谢维持着一种均衡,即获得代谢解毒能力并保持其它代谢正常有序,来达到代谢杀虫剂的目的。虫体的抗药代谢过程是相当复杂的,有多种解毒酶、水解酶、合成酶和靶标酶以及特异蛋白质的参与(Araújo et al.,2008;Huang et al.,2004;Oppenoorth,1965)。本文只选用了3种报道较多的解毒酶,其通径分析结果也只是显示出在3种酶活性相互影响下的抗性水平关系,如MFO-O-脱甲基活性和CarE 活性对敌百虫的抗药性水平起负相关的结果,即表明MFO-O-脱甲基活性和CarE 活性是相对于GST 活性的比较下所呈现负向作用,并不是说明MFO-O-脱甲基活性和CarE活性对敌百虫代谢变化中表现负向关系。
昆虫的抗性分子遗传和进化的关键就是点突变和调节突变,而代谢抗性很可能是调节突变,引起昆虫个体水平对杀虫剂适应性进化的主要原因是抗性相关酶量或活性的变化 (刘泽文等,2002)。当前,昆虫抗药性的生理学和分子生物学研究已经取得了一定的进展,但害虫抗药性发展规律和趋势预测应与此相对应的生理生化等指标相结合,才能深入了解这几种解毒酶的活性与抗性水平的关系,为克服和延缓桔小实蝇的田间抗性水平提供重要的科学理论依据。
References)
Allwood AJ,Chinajariyawong A,Drew RAI,et al.Host plant records of fruit flies (Diptera:Tephritidae)in Southeast Asia [J].Raffles Bulletin of Zoology Supplement,199,7:7-92.
Araújo RA,Guedes RNC,Oliveira MGA,et al.Enhanced proteolytic and cellulolytic activity in insecticide-resistant strains of the maize weevil,Sitophilus zeamais [J].Journal of Stored Products Research,2008,44 (4):354-359.
Bateman MA.The ecology of fruit flies [J].Annual Review of Entomology,1992,17 (1):493-518.
Bradford MM.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding [J].Analytical Biochemistry,1976,72 (1-2):248-254.
Chen ZH,Liu CX,Li FL,et al.Development of Diamond-back moth strains resistant to dimehypo and cartap with reference to the mechanism of resistance [J].Acta Entomologica Sinica,1993,36(4):410-418.[陈之浩,刘传秀,李凤良,等.杀虫双和杀螟丹选育对小菜蛾抗药性的形成及其抗性机制[J].昆虫学报,1993,36 (4):410-418]
Fan JA.Studies on the assessment of potential establishment of oriental fruit (Bactrocera dorsalis)in China with synthetic judgement on fuzzy mathematics [J].Plant Quarantine,1998,12 (2):76-80.[范京安.用模糊综合评判法探讨桔小实蝇在中国的适生分布[J].植物检疫,1998,12 (2):76-80]
He L,Tan SL,Cao XF,et al.Study on resistance selection and activity of detoxifieation enzyme in Tetranychus cinnabarinus (Boiduval)[J].Chinese Journal of Pesticide Science,2003,5 (4):23-29.[何林,谭仕禄,曹小芳,等.朱砂叶螨的抗药性选育及其解毒酶活性研究[J].农药学学报,2003,5 (4):23-29]
Hou BH,Zhang RJ.Potential distributions of the fruit fly Bactrocera dorsalis (Diptera:Tephritidae)in China as predicted by CLIMEX[J].Acta Ecologica Sinica,2005,25 (7):1569-1574.[侯柏华,张润杰.基于CLIMEX 的桔小实蝇在中国适生区的预测[J].生态学报,2005,25 (7):1569-1574]
Huang ZW,Shi P,Dai JQ,et al.Protein metabolism in Spodoptera litura (F.)is influenced by the botanical insecticide azadirachtin[J].Pesticide Biochemistry and Physiology,2004,80 (2):85-93.
Hung,CF,Sun CN.Microsomal monooxygenases in diamondback moth larvae resistant to fenvalerate and piperonyl butoxide [J].Pesticide Biochemistry and Physiology,1989,33 (2):168-175.
Jia CS.Calculatiang the LC50of insecticides with software SPSS [J].Entomological Knowledge,2006,43 (3):414-417.[贾春生.利用SPSS 软件计算杀虫剂的LC50[J].昆虫知识,2006,43(3):414-417]
Jiang XL,He WZ,Xiao S,et al.Study on the biology and survival of Bactrocera dorsalis in the border region of Yunnan [J].Journal of Southwest Agricultural University,2001,23 (6):510-517.[蒋小龙,和万忠,肖枢,任丽卿,等.桔小实蝇在云南边境生物学研究及适生性分析[J].西南农业大学学报,2001,23 (6):510-517]
Jin Tao,Zeng Ling,Lin Yuying,et al.Insecticide resistance of the oriental fruit fly,Bactrocera dorsalis (Hendel) (Diptera:Tephritidae),in mainland China [J].Pest Management Science,2011,67 (3):370-376.
Lin Yuying,Jin Tao,Zeng Ling,et al.Insecticide toxicity to oriental fruit fly Bactrocera dorsalis (Diptera:Tephritidae)is influenced by environmental factors [J].Journal of Economic Entomology,2013,106 (1):353-359.
Liu YJ,Shen JL,Jia BT,et al.Resistance reduction to lambda-cyhalothrin and activity change of multi-function oxidases in the beet armyworm,Spodoptera exigua,under non-selection pressure in the laboratory [J].Acta Entomologica Sinica,2007,50 (4):349-354.[刘永杰,沈晋良,贾变桃,等.甜菜夜蛾不同世代对氯氟氰菊酯抗性减退及多功能氧化酶系活性变化[J].昆虫学报,2007,50 (4):349-354]
Liu ZW,Han ZJ,Wang YC.The Roles of Carboxylesterase and acetylcholinesterase insensitivityin development of methamidophos resistance in Nilaparvata lugens [J].Chinese Journal of Pesticide Science,2002,4 (4):51-55.[刘泽文,韩召军,王荫长.羧酸酯酶和乙酰胆碱酯酶在褐飞虱对甲胺磷抗性发展中的作用[J].农药学学报,2002,4 (4):51-55]
Liu ZW,Han ZJ,Zhang LC.The Roles of detoxifying enzymes and AChE insensitivity in methamidophos resistance development and decline in Nilaparvata lugens [J].Insect Science,2003,10 (3):179-184.
Oppenoorth FJ.Biochemistry and physiology of resistance [J].New york,Plenum Press,1976.
Oppenoorth FJ.Biochemical genetics of insecticide resistance [J].Annual Review of Entomology,1965,10 (1):185-206.
Pan ZP,Lu YY,Zeng L,et al.Development of resistance to trichlorophon,alphamethrin,and avermectin in laboratory populations of the oriental fruit fly,Bactrocera dorsalis (Hendel)(Diptera:Tephritidae)[J].Acta Entomologica Sinica,2008,51(6):609-617.[潘志萍,陆永跃,曾玲,等.桔小实蝇实验种群对敌百虫、高效氯氰菊酯和阿维菌素的抗性增长规律[J].昆虫学报,2008,51 (6):609-617]
Pan ZP,Zeng L,Lu YY.Monitoring of resistance of oriental fruit fly adults to insecticides in South China [J].Journal of South China Agricultural University,2005,26 (4):23-26.[潘志萍,曾玲,陆永跃.华南地区桔小实蝇对几种农药的抗药性研究[J].华南农业大学学报,2005,26 (4):23-26]
Terriere LC.Induction of detoxication enzymes in insects [J].Annual Review of Entomology,1984,29 (1):71-88.
vail PV,Seay RE,DeBolt J.Microbial and chemical control of the cabbage looper on fall lettuce [J].Journal of Economic Entomology,1980,73 (1):72-75.
Van Pottelberge S,Van Leeuwen T,Van Amermaet K,et al.Induction of cytochrome P450 monooxygenase activity in the two-spotted spider mite Tetranychus urticae and its influence on acaricide toxicity[J].Pesticide Biochemistry and Physiology,2008,91 (2):128-133.
Xiong Y,Xiang Y,Bao YX,et al.Study on the suitable establishment of Bactrocera dorsalis Hendel in Jiangsu Province,China [J].Journal of Nanjing Agricultural University,2006,29 (4):48-52.[熊焰,项勇,包云轩,等.桔小实蝇在江苏省的适生性分布研究[J].南京农业大学学报,2006,29 (4):48-52]
Zhang HY,Chi GT,Zhang JL.The progress of detoxification enzyme systems and resistance of insecticide [J].Journal of Agricultural University of Hebei,2002.25 (S1):93-95.[张红英,赤国彤,张金林,2002.昆虫解毒酶系与抗药性研究进展[J].河北农业大学学报,2002,25 (S1):93-95]
Zhang YM,Sun CX,Li JY.Detection of insecticide resistance and its relationship with two detoxification enzymes of Musca domestica in different areas of Tianjin [J].Acta Entomologica Sinica,2006,49(3):538-542.[张咏梅,孙晨熹,李今越.天津地区家蝇抗药性水平及与两种解毒酶的关系[J].昆虫学报,2006,49(3):538-542]
Zhang YP,Zeng L,Lu YY,et al.Monitoring of insecticide resistance of Bactrocera dorsalis adults in South China [J].Journal of South China Agricultural University,2007,28 (3):20-23.[章玉苹,曾玲,陆永跃,等.华南地区桔小实蝇抗药性动态监测[J].华南农业大学学报,2007,28 (3):20-23]
Zhang YP,Zeng L,Lu YY.Resistance stability and re-growth in adults of the oriental fruit fly,Bactrocera dorsalis (Diptera:Tephritidae)to trichlorphon [J].Acta Entomologica Sinica,2008,51 (10):1044-1049.[章玉苹,曾玲,陆永跃,等.桔小实蝇对敌百虫抗性稳定性及再增长趋势[J].昆虫学报,2008,51 (10):1044-1049]
Zhang YP,Zeng L,Lu YY,et al.Monitoring of Insecticides Resistance of Oriental Fruit Fly Field Populations in South China [J].Journal of Huazhong Agricultural University (Natural Science Edition),2008,27 (4):456-459.[章玉苹,曾玲,陆永跃,等.华南地区桔小实蝇田间种群抗药性的监测[J].华中农业大学学报(自然科学版),2008,27 (4):456-459]
Zhang ZX,Xu HH,Cheng DM.Calculating toxicity regression with EXCEL [J].Entomological Knowledge,2002,39 (1):67-70.[张志祥,徐汉虹,程东美.EXCEL 在毒力回归计算中的应用[J].昆虫知识,2002,39 (1):67-70]
Zhou CL,Tang ZH,Zhang LM.Resistance of diamondback moth to synthetic pyrethroids and its relationship with microsomal mixed-founction oxidase [J].Acta Phytophylacica Sinica,1993,20(1):91-95.[周成理,唐振华,张丽妹.小菜蛾幼虫对拟除虫菊酯类杀虫剂的抗药性与多功能氧化酶的关系[J].植物保护学报,1993,20 (1):91-95]
猜你喜欢
江苏工程职业技术学院学报(2022年1期)2022-05-05
黑龙江气象(2021年2期)2021-11-05
数理化解题研究(2021年13期)2021-08-19
机械工程与自动化(2020年6期)2020-12-28
农业知识(2020年3期)2020-12-18
食品与机械(2018年3期)2018-05-31
蔬菜(2016年8期)2016-10-10
湖南农业(2016年3期)2016-06-05
湖南农业(2016年3期)2016-06-05
湖南农业(2015年5期)2015-02-26
