
晚稻田褐飞虱和白背飞虱种群结构时序动态研究
2014-12-14 02:50李小珍李少山王建国徐昭焕
江西农业大学学报 2014年5期
李小珍,李少山,王建国,刘 伟,徐昭焕
(江西农业大学 农学院,江西 南昌 330045)
昆虫种群结构主要包括年龄结构、多型现象和性别比例,是昆虫生物生态学的重要研究内容。昆虫种群结构既取决于种的遗传特性,同时也取决于具体的环境条件[1]。因此,同一昆虫种类在不同年份、同一年份不同季节或同一季节不同生境均可能表现出不同的种群结构特点。害虫种群结构时序动态的研究,可以阐明和预测有害生物在发生过程中数量变动的规律性,对保证农业的稳定生产有重要的经济意义。
褐飞虱Nilaparvata lugens(Stål)和白背飞虱Sogatella furcifera(Horváth)是中国、日本和韩国等亚洲国家广大稻区的重要迁飞性害虫[2-4]。近年来,这两种飞虱呈逐年加重危害的趋势,这除了与迁入的虫源基数、迁入地的气候条件以及水稻的生育期等种群外部环境条件有关外,也与种群迁入定殖后,种群内部年龄结构、翅型与性别的分化等密切相关。目前,国内外对褐飞虱和白背飞虱种群年龄结构和雌雄性别时序变化规律的研究较少,而翅型分化方面的研究则较多,并已经证实在不同温度、光周期、若虫密度、寄主植物、水稻生育期和外源激素类物质处理下,长、短翅型分化明显不同[5-10]。然而,这些研究多数在室内设定的条件下进行,而就大田环境条件下褐飞虱和白背飞虱翅型分化的动态研究仍然不多。本文通过对2010—2011 年江西南昌地区晚稻田内稻飞虱的系统调研,分析褐飞虱和白背飞虱的成虫和若虫动态、翅型和性别分化时序动态规律,对稻飞虱的短期预测预报与可持续治理具有积极作用。
1 材料与方法
1.1 材料
试验研究地点:江西南昌江西农业大学教学科研基地,东经115.72°,北纬28.76°。水稻品种:中优赣晚籼19 号,为江西主要栽培的水稻品种之一,在该水稻品种上研究稻飞虱的时序结构特点。2010—2011 年南昌温度等资料来源于江西省气候资料处理部门逐月上报的《地面气象记录月报表》的信息化资料,图中温度为调查日前7 天的温度均值。
1.2 方法
1.2.1 试验设计 试验调研于2010 年6 月1 日—10 月4 日和2011 年6 月4 日—10 月6 日,连续2 年在江西农业大学教学科研基地进行。水稻种植分为3 个小区,每个小区约100 m2,每个小区之间间隔约1 m。水稻于5 月底种植,返青后开始调查稻飞虱;10 上旬水稻黄熟收割后,停止调查。水稻种植期间,不施用任何化学杀虫剂防治水稻上的病虫害。
1.2.2 调研方法 每个小区对角线方法抽取3 个点,3 个小区共取9 个调查点,每个点取水稻1 纵,每7 d调查1 次。在水稻整个生育期间,2010 年和2011 年分别调查了19 次。以一个点的稻飞虱调查为例,取长×宽×高=40 cm×30 cm×8 cm 的长方形塑料托盘装适量肥皂水(约0.2 cm,浸满托盘底部即可),滴入几滴油、混均,以防止落盘飞虱从托盘跳出。将托盘轻放在选好的水稻基部,用手拍打水稻叶鞘部位4~5 次,拍完后取回托盘,上面迅速蒙上带孔纱网,计数收集的各种飞虱的成虫和若虫、长翅和短翅、雌性和雄性数量。部分难以鉴定的个体,用吸虫管转移到试管,带回室内解剖镜下鉴定。
1.3 数据统计与分析
以Berger-Parker(1974)提出的优势度指数(D)计算各种类的优势度,D=Ni/N。其中,N=∑Ni为所有物种个体数量之和,Ni为i 物种的个体数量。采用SPSS 12.0 内的二元一次线性回归方程分析褐飞虱和白背飞虱田间成虫种群密度(Y)与短翅率(X1)和性比(X2)之间的关系。
2 结果与分析
2.1 种类构成
江西南昌地区,水稻上共调查到4 种飞虱即褐飞虱、白背飞虱、灰飞虱和稗飞虱。2010 年稻飞虱发生重,收集到的各种飞虱的数量均高于2011 年数量。2010 年,褐飞虱的优势度指数最高,达到0.635 5;白背飞虱次之,为0.346 5。2011 年,白背飞虱的优势度指数最高,为0.497 7;褐飞虱次之,为0.423 9,这两种飞虱为水稻上的优势种类。灰飞虱和稗飞虱的数量少,优势度指数低,未对这两个种群的时序结构动态进行分析。

表1 2010—2011 年南昌地区稻飞虱种类的优势度分析Tab.1 Dominance indexes of rice planthopper in Nanchang area
2.2 种群动态
2010—2011 年6 月中旬之前,晚稻上仅出现褐飞虱成虫,且密度低。6 月中旬之后,若虫陆续出现,为刚羽化的第1 代若虫。8 月中旬开始,褐飞虱成虫和若虫密度急剧上升,2010 年于9 月6 日达到高峰,成虫密度达到(116.0±12.1)头/丛,若虫密度达到(137.3±21.0)头/丛;2011 年于9 月15 日达到高峰,成虫密度达到(13.3±5.0)头/丛,若虫密度达到(27.3±7.1)头/丛。到水稻收割,成虫和若虫密度平缓下降(图1)。

图1 2010 年(左)—2011 年(右)褐飞虱成虫和若虫的田间动态(Mean±SE)Fig.1 Dynamics of N.lugens adults and nymph from 2010(left)to 2011(right)in late rice fields(Mean±SE)
白背飞虱在6 月上旬即水稻刚刚栽培时就开始出现,6 月下旬到7 月中旬分别出现若虫和成虫的第1个密度小高峰。2010 年6 月27 日若虫出现第1 个密度峰值为(5.0±1.5)头/丛,成虫于7 月11 日出现第一个密度峰值为(27.3±5.8)头/丛;成虫和若虫均于8 月24 日出现第2 个密度峰值,成虫密度达到(99.3±12.8)头/丛,若虫密度达到(104.7±19.8)头/丛。2011 年成虫的第一个密度峰值不明显,若虫于7 月21 日出现,密度达到(12.7±4.7)头/丛;第2 个峰值,若虫于8 月25 日出现,密度达到(56.3±10.7)头/丛,成虫于9 月1 日出现,密度达到(28.3±13.1)头/丛。随后直到水稻收割,白背飞虱成虫和若虫密度迅速下降(图2)。

图2 2010 年(左)—2011 年(右)白背飞虱成虫和若虫的田间动态(Mean±SE)Fig.2 Dynamics of S.furcifera adults and nymph from 2010(left)to 2011(right)in late rice fields(Mean±SE)
由此可见,2010 年在水稻赣晚籼19 号上不管是褐飞虱还是白背飞虱均发生较重,2011 年明显较轻。白背飞虱出现两个高峰,且整体发生的时间较早;褐飞虱发生时间较晚,仅一个明显的密度峰值。从个体数量上看,调查到的褐飞虱和白背飞虱的若虫密度比成虫密度高。
2.3 翅型分化
2010 年6 月1 日调查开始田间就有长翅型褐飞虱,应该是迁入的成虫。短翅型成虫于6 月下旬开始出现。随后长翅型和短翅型个体数均逐渐增加,并于9 月6 日同时达到峰值,长翅型密度为(34.7±8.1)头/丛,短翅型密度为(81.3±16.3)头/丛。8 月17 日之前,长翅型虫口密度较高;之后短翅型虫口密度较高。2011 年,长翅型和短翅型成虫分别于6 月1 日和6 月29 日开始出现,并于9 月15 日达到虫口密度高峰,分别为(7.00±2.2)头/丛和(6.33±2.3)头/丛,长、短翅型虫口密度变化较小(图3)。

图3 2010 年(左)—2011 年(右)褐飞虱田间种群翅型分化规律(Mean±SE)Fig.3 Wing differentiation of N.lugens adults from 2010(left)to 2011(right)in late rice fields(Mean±SE)
白背飞虱长翅型成虫自2010 年6 月1 日开始出现后,并于8 月24 日达到最高密度(93.3±17.7)头/丛,短翅型最高虫口密度仅为(6.0±2.1)头/丛;2011 年,长翅型最高虫口密度为(27.3±12.1)头/丛,短翅型个体仅零星出现(图4)。综合来看,白背飞虱在水稻各生育期间均以长翅型为主,短翅型个体少量出现。

图4 2010 年(左)—2011 年(右)白背飞虱田间种群翅型分化规律(Mean±SE)Fig.4 Wing differentiation of S.furcifera adults from 2010(left)to 2011(right)in late rice fields(Mean±SE)
2.4 性别分化
2010—2011 年8 月中旬之前,褐飞虱田间虫口密度较低,雌性密度略高于雄性虫口密度,雌雄性比≤3;8 月中旬之后,田间雌、雄虫口密度急剧升高,但雌性虫口密度增加速度较快,导致雌雄性比进一步扩大,于2010 年9 月28 日达到最大值为6.7;2011 年9 月22 日达到最大值为4.3(图5)。
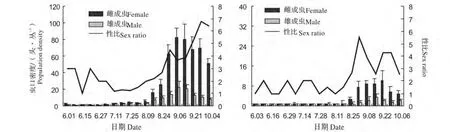
图5 2010 年(左)—2011 年(右)褐飞虱田间种群的性别分化动态(Mean±SE)Fig.5 Sex differentiation of N.lugens adults from 2010(left)to 2011(right)in late rice fields(Mean±SE)
白背飞虱雌性虫口密度高于雄性虫口密度。8 月中旬之前,白背飞虱虫口密度不高,雌性密度略大于雄性虫口密度;8 月下旬之后,雌、雄虫口密度均快速增加,但雌雄性比变化不大,2010 年和2011 年雌雄性比均维持在1~4(图6)。
综合来看,在褐飞虱和白背飞虱田间种群的性别分化中,通常雌雄性比大于1;褐飞虱随着虫口密度的增加,性比扩大;白背飞虱的性比则相对稳定。

图6 2010 年(左)—2011 年(右)白背飞虱田间种群的性别分化动态(Mean±SE)Fig.6 Sex differentiation of S.furcifera adults from 2010(left)to 2011(right)in late rice fields(Mean±SE)
2.5 种群密度与短翅率和性比的关系
根据褐飞虱和白背飞虱田间成虫种群密度(Y)与短翅率(X1)和性比(X2)之间的二元一次线性回归方程的结果,表明褐飞虱种群密度与短翅率和性比存在显著的线性相关性(2010 年,r=0.87,P<0.01;2011 年,r=0.80,0.01<P<0.05),表明田间种群密度升高,可能导致短翅率和性比升高。而白背飞虱种群密度与短翅率和性比的线性相关性不显著(2010 年,r=0.31,P>0.05;2011 年,r=0.76,P>0.05)(表2)。

表2 种群密度(Y)与短翅率(X1)和性比(X2)的相关回归分析Tab.2 Correlation and regression analyses of population density with percentage of brachypterous individual and sex ratio
3 讨论
两年的系统调查表明,白背飞虱在水稻上出现的密度高峰较褐飞虱早,主要是由于白背飞虱先于褐飞虱迁飞进行江西境内。徐善忠[11]和赵险峰等[12]灯下诱集表明,白背飞虱在正常年份于5 月上中旬开始迁飞进入江西吉安地区,褐飞虱则在5 月下旬至6 月上旬开始迁入,因此6 月上旬在江西南昌稻田收集到的白背飞虱和褐飞虱可能是刚迁入的个体。水稻生育前期白背飞虱的密度较高,后期则褐飞虱的密度较高,一方面这与迁入时间有关,另一方面与它们的栖息习性有关,通常白背飞虱在水稻上的栖息部位较褐飞虱高,群集拥挤的习性较褐飞虱差,密度较高时,即转移至其他植株[13]。
不少学者认为,褐飞虱的翅型分化具有密度依赖性[14-15]。Kisimoto[16]室内研究表明,在若虫低密度下,极少出现短翅型,高密度下也极少,而在中间的某一密度范围内比例最高。张增全[17]室内外试验表明,稻株虫口密度与短翅型出现比率之间,不存在随虫口密度提高而短翅型出现比率下降的可能性。笔者的田间调查数据表明,褐飞虱在高密度下,短翅型的密度明显高于长翅型的密度;在低密度下,长、短翅型密度则无明显变化,结果同样可以说明褐飞虱的翅型分化与种群密度有关。
白背飞虱在水稻各生育期间均以长翅型为主,短翅型个体出现较少,这一结论与胡国文和朱明华[18]提出的白背飞虱短翅型雄虫极为少见的结论相似。刘佳妮等[19]曾比较了不同温度、光周期、若虫密度、寄主植物、水稻生育期和植株营养下白背飞虱的翅型分化状况,发现无论在哪一条件组配下白背飞虱长翅型的分化率始终高于短翅型,这同样证实白背飞虱短翅型个体分化较低。与褐飞虱相似,白背飞虱的翅型分化同样存在密度依赖性[20],由于笔者在稻田自然环境下未能控制其它因子如温度、光照和植株营养等的影响,未能对其密度依赖性作进一步分析。
昆虫性别决定与分化是个体正常发育和生存不可缺少的一环,也是种族得以繁衍延续的物质基础[21]。褐飞虱和白背飞虱在水稻各个生育期,雌性个体占优势,即性比≥1,而就其性比发生的原因本文未作探讨。在褐飞虱和白背飞虱田间种群的性比分化规律方面,还未见报道,仅陈福寿[22]就温度对半闭弯尾姬蜂发育和性比的影响作了研究。
本文仅调研了2010—2011 年白背飞虱和褐飞虱在江西南昌地区晚稻田内的消长动态、翅型和性别分化规律。由于不同年份白背飞虱和褐飞虱生长、发育和繁殖的环境条件并不一样,同时迁入的日期和基数也不相同,白背飞虱和褐飞虱的种群时序结构规律在不同年份同样存在差异。因此,要进一步从宏观上明确白背飞虱和褐飞虱田间种群结构的时序分化规律,需要进一步累积多年不同水稻品种上的田间调研数据。
[1]戈峰.现代生态学[M].北京:科学出版社,2002:109-110.
[2]汤金仪,胡泊海,王建强.我国水稻迁飞性害虫猖獗成因及其治理对策建议[J].生态学报,1996,16(2):167-173.
[3]黄凤宽,韦素美,黄所生.稻褐飞虱翅型分化研究进展[J].西南农业学报,2003,16(1):82-85.
[4]周强,张润杰,古德祥.白背飞虱在稻田内空间结构的分析[J].昆虫学报,2003,46(2):171-177.
[5]Iwanaga K,Tojo S,Nagata T.Immigration of the brown planthopper,Nilaparvata lugens,exhibiting various responses to density in relation to wing morphism[J].Entomologia Experimentalis et Applicata,1985,38:101-108.
[6]Iwanaga k,Tojo S.Comparative studies on the sensitivities to nymphal density,photoperiod and rice plant stage in two strains of the brown planthopper Nilaparvata lugens Stål(Homoptera:Delphacidea)[J].Japanese Journal of Applied Entomology and Zoology,1988,32:68-74.
[7]Shobu S,Mikuriya H,Yamaguchi J,et al.Fluctuations and factors affecting the wing-form ratio of the brown planthopper,Nilapamata lugem(stål)in rice fields[J].Japanese Journal of Applied Entomology and Zoology,2002,46(3):135-143.
[8]祝增荣,程家安,刘永军.白背飞虱长、短翅型成虫实验种群生物学比较[J].中国水稻科学,2001,15(3):229-232.
[9]张琼秀,孙梓暄,李广宏,等.三种外源激素类物质对白背飞虱(Sogatella furcifera)翅型分化的影响[J].生态学报,2008,28(12):5994-5998.
[10]吕再萍,杨长举,姚英娟,等.外界环境对褐飞虱翅型分化的影响研究[J].湖南农业科学,2009,48(8):1866-1868.
[11]徐善忠,曾宜杰.2002 年泰和县稻飞虱监测动态[J].江西植保,2003,26(1):28.
[12]赵险峰,袁雪梅,邹金苟,等.2003 年上高县早稻稻飞虱大发生[J].江西植保,2003,26(4):159-160
[13]朱春福,杨慧中,陈超,等.不同光周期条件下白背飞虱和褐飞虱翅型分化[J].江西植保,2010.33(3):109-110.
[14]Denno R F,Roderick G K,Dobel H G.Density-related migration in planthoppers(Homoptera:Delphacidae):the role of habitat persistence[J].Am Nature,1991,38:1513-1541.
[15]王希仁,张灿东.稻虱翅型分化因子的探讨[J].昆虫知识,1981,18(4):145-148.
[16]Kisimoto R.Studies on the polymorphism and its role playing in the population growth of the brown planthopper,Nilaparvata lugens Stål[J].Bulletin of Shikoku Agricultural Experiment Station,1965,13:1-106.
[17]张增全.褐稻虱翅型分化的研究[J].昆虫学报,l983.26(3):260-265.
[18]胡国文,朱明华.白背飞虱短翅型雄虫的观察[J].昆虫知识,1981,18(4):154-156.
[19]刘佳妮,桂富荣,李正跃.影响白背飞虱翅型分化的相关因子[J].植物保护学报,2010,37(6):511-516.
[20]Matsumura M.Genetic analysis of a threshold trait:density-dependent wing dimorphism in Sogatella furcifera(Horváth)(Hemiptera:Delphacidae),the whitebacked planthopper[J].Heredity,1996,76:229-237.
[21]查幸福,夏庆友,向仲怀.昆虫性别决定的分子机制研究进展[J].蚕业科学,2006,32(2):151-156.
[22]陈福寿,王燕,李志敏,等.温度对半闭弯尾姬蜂发育和性比的影响[J].环境昆虫学报,2011,33(2):257-260.
猜你喜欢
作物学报(2022年11期)2022-08-31
农业工程学报(2022年10期)2022-08-22
上海农业科技(2022年4期)2022-08-15
天津农业科学(2022年5期)2022-05-31
昆虫学报(2021年8期)2021-09-06
海峡科学(2021年6期)2021-08-04
园艺与种苗(2021年6期)2021-07-19
昆虫学报(2020年8期)2020-10-15
环境昆虫学报(2016年6期)2016-12-27
长江大学学报(自科版)(2015年21期)2015-12-04
