
台湾桤木引种的生长节律与生物量研究
2014-12-28 06:37彭玉华黄志玲申文辉朱积余郝海坤
中南林业科技大学学报 2014年6期
彭玉华,黄志玲,申文辉,朱积余,郝海坤
(1. 广西壮族自治区林业科学研究院; 2. 国家林业局中南速生材繁育实验室;3. 广西优良用材林资源培育重点实验室,广西 南宁 530002)
台湾桤木引种的生长节律与生物量研究
彭玉华1,2,3,黄志玲1,2,3,申文辉1,2,3,朱积余1,2,3,郝海坤1,2,3
(1. 广西壮族自治区林业科学研究院; 2. 国家林业局中南速生材繁育实验室;3. 广西优良用材林资源培育重点实验室,广西 南宁 530002)
研究台湾桤木年生长动态、植株生物量积累与分配模式以及植株生长指标与生物量组分的相关性,结果表明:台湾桤木年生长规律符合Logistic曲线特征,据此方程可以建立株高、地径与生长时间的拟合模型, 决定系数(R2)均大于0.94,达到显著相关,具有非常高的可靠性,因此利用Logistic曲线方程对台湾桤木株高和地径年生长节律进行拟合是可行的;根据Logistic曲线方程上两个曲率最大的点准确地将台湾桤木年生长进程划分为:渐增期、速生期和缓增期, 其中处于速生期植株的株高和地径生长量均超过全年总量的70%;苗木生物量积累与苗高和地径的相关性极为密切,其中地上部分生物量占全株的78.06%,枝和叶的干质量占地上干质量的53.95%,全株含水量是全株干质量的1.39倍,大于5 mm的侧根平均每株达9条;为台湾桤木在广西成功引种栽培提供科学理论依据。
台湾桤木;生长节律;Logistic曲线;生物量;含水量
台湾桤木Alnus formosana为桦木科桤木属落叶大乔木,在原产地高达20 m以上,胸径30 cm。其干形通直、高大、木材细软、纹理直,韧性好,易于加工,成型,气干密度0.52 g/cm3; 耐腐性、耐蛀性能好,属二级木,可用于建筑、器具、箱板、家具和造纸等。其适应性强,喜光,喜湿且耐湿,耐瘠薄,并具一定的耐干旱能力,生长快,分布范围广,已逐渐成为当地水土保持和涵养水源的重要树种[1-3],以及重要的药用植物[4]。台湾桤木为非豆科菌根树种,能与固氮菌共生、固定空气中游离的氮素,为重要的培肥树种[5-6]。目前,广西人工林种植以松、杉、桉为主要树种,树种单一[7],且连续针叶林经营中,林分层次结构简单,伴生树种、下木和草本等植物种类稀少,造成生态系统多样性下降,导致地力衰退、林分稳定性差,抵御自然灾害能力减弱,找到能够保持地力并且能够满足工业需求的树种成为大家的理想。台湾桤木适应性广,生长迅速,并能进行生物固氮,增强林地肥力,台湾恺木的原产地台湾,其水热条件和地理环境与广西地区相似,具有较高的引种和应用推广价值。台湾恺木自身具有的固氮特性、生长发育特性和较高的商用价值可用于当前的天然林保护工程,退耕还林工程和生态环境治理工程,这不仅可丰富我国南方地区阔叶造林树种种类,而且对改良土壤具有重大意义。
台湾桤木在我国大陆引种已有近30年的历史,已成为我国大陆引种栽培成功的树种之一,台湾桤木已成为我国长江以南部分地区极有潜力的商用阔叶造林树种之一,无论是从生态保护,还是致力于经济发展,皆具有广阔的应用前景。早期营造的台湾桤木人工林已经成材,成为家具、建筑和制板业等的优质原材料[7]。我国福建1987 年最早引种台湾桤木,8 年生台湾桤木平均树高为 11.1 m,胸径为 11.8 cm,表现出了明显的优势[9]。目前在我国有关台湾桤木的研究已做了大量的工作,有台湾桤木引种栽培、扦插繁殖技术、采穗圃建立技术、光合生理特性、花粉生活力的测定方法、生长性状等等研究[8-17],但对台湾桤木生长前期的生长节律研究较少,仅见有熊大国等[18]对台湾桤木在元坝区生长动态的研究。本文报道台湾桤木在广西南宁引种的年生长节律和生物量,为台湾恺木在广西的引种和发展提供相应的理论依据和技术支撑。
1 材料与方法
1.1 引种试验区概况
试验设置于广西林业科学研究院科研试验地,108°56′E,22°56′N,海拔约 80 m,≥ 10℃积温为7 200℃,年平均气温21.7℃,极端最低温-1.5℃,极端最高温39.4℃,全年无霜期高达360 d,年均降雨量为1 300 mm,年均相对湿度为80%左右。土壤类型为砖红壤性红壤,pH值为5~6,肥力中等。
1.2 研究方法
1.2.1 台湾桤木生长节律的测定方法
2012年3月从四川调入台湾桤木1年生播种苗,平均苗高为85.45 cm、地径为7.12 mm。2012年3月30日种植在广西林业科学研究院试验林区,种植1个月后结合铲草追施第1次肥(复合肥150 g/株),3个月后铲草施肥第2次(复合肥250 g/株),期间注意防治病虫害。2012年4月15日开始测定,每隔15 d固定测量其苗高和地径(如遇特殊情况不能测量,延后或提前1 d),至2013年4月15日停止测定。苗高生长量用钢卷尺量,精度到0.1 cm,地径用电子游标卡尺量,精度到0.01 mm。
1.2.2 试验设计
采取3次重复,每重复随机选取30株作为固定标准株,四周均预留保护行。
1.2.3 台湾桤木生物量测定方法
生物量指的是生物现存量,平均标准木法测定。在每重复固定测定的30标准株中,从中选取5株接近于平均木进行生物量测定,生物量分地上部分(分主茎、侧枝、叶)、地下部分,用电子天平称分别称鲜重,之后自然干燥, 4个月后分别测定其气干重。
1.3 分析方法
1.3.1 Logistic曲线拟合
以生长时间(t)为自变量,累积生长量(Y)为应变量,根据拟合方程[19-21]建立台湾桤木种植1年的Logistic生长曲线、连日生长量变化速率曲线。

式中:Y为苗木累积生长量;t为苗木生长天数;a,b是待定系数,用最小二乘法求得;k表示苗木生长极限值,用苗高或地径的四点法[22]求得:
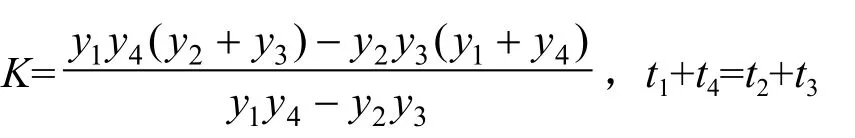
式中:(t1,y1)、(t4,y4)分别为实测数据序列的始点、终点,(t2,y2)、(t3,y3)则为实测数据序列中间两点。
1.3.2 连日生长量变化速率曲线
对Logistic曲线方程求一阶导数得连日生长量随时间变化的方程,求二阶导数为连日生长量的变化函数曲线,其公式为:
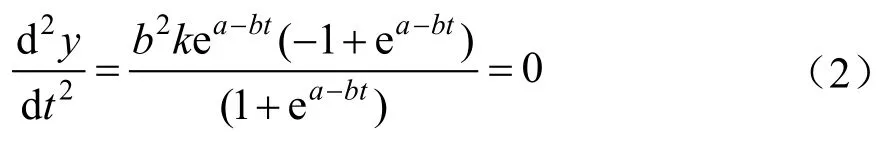
令(2)式为0求得理论上连日生长量的速生点


对(1)式求三阶导数并令其等于0求得理论上连日生长量变化速率最快的2个点:
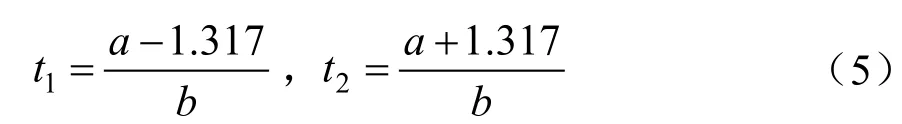
根据Logistic方程的基本特征,利用(5)求得Logistic曲线上的两个生长拐点(t1,y1)、(t2,y2),(t1,y1)是植株由渐增到速生的分界点,(t2,y2)是植株由速生到缓慢增长的分界点。因此,这两个拐点能准确地将植株的生长进程划分为渐增期(0,t1)、速生期(t1,t2)和缓增期(t2,+∞)。其中,t1、t2间苗木平均生长速度为线性生长速度(VLGR),t1到t2苗木生长量为线性生长量(ATLG)。经计算可得:
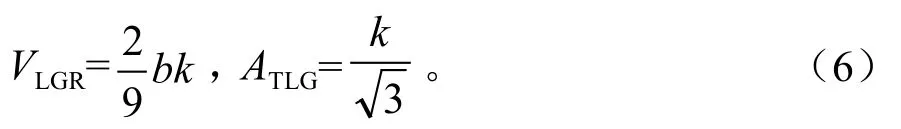
根据(3)、(4)、(5)、(6)式求得台湾桤木株高、地径的速生点、两个拐点和苗木最大线性生长速率(RMGR)、线性生长速度(VLGR)和线性生长量(ATLG)。
数据分析采用SPSS17.0统计软件进行。
2 结果与分析
2.1 台湾桤木种植年生长规律
试验结果表明,台湾桤木在广西南宁地区种植年生长无封顶期,株高、地径生长明显呈现S型生长,其最终生长量平均值株高达249.67 cm、地径为36.52 mm。从图1中可以看出:株高生长第1个高峰期出现在植后90天(即6月1日至6月15日),15 d的株高净增达12.29 cm,第2个高峰期呈速增速减,出现在植后225天(即11月1日至11月15日),15 d的株高净增达12.93 cm,虽然植后1年的株高15 d净增有14.35 cm,但因测定时间与第1次高峰的生长时间基本在一个时间段,故把它归为第1次生长高峰期;地径生长全年只有1次生长高峰,出现在植后90后(即6月1日至6月15日),15天的地径净增生长量达3.2 mm。
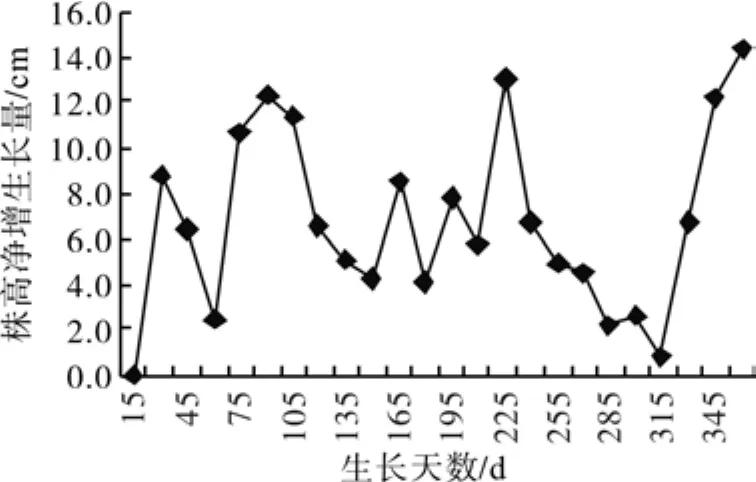
图1 株高净增生长量Fig. 1 Net increment of height
2.2 台湾桤木年生长模型的建立与拟合
台湾桤木生长拟合曲线见图3、图4。方差分析结果、拟合方程见表1,由表1可知:台湾桤木的株高、地径Logistic方程拟合模型回归关系达到极显著水平,决定系数分别达0.986和0.948,表明方程具有非常高的可靠性,这点可从图3、图4中株高、地径生长的实际观测值与理论拟合值的对比关系得到证实,因此利用Logistic方程曲线方程对台湾桤木株高和地径年生长节律进行拟合是可行的,可用于台湾桤木年生长的分析和预测。
2.3 台湾桤木株高、地径连日生长量拟合与生长阶段划分
2.3.1 株高、地径连日生长量的拟合

图2 地径净增生长量Fig. 2 Net increment of basal diameter
从表2可见:理论上株高生长最大时间为108.25 d(7月18日),与观测值90 d(6月30日)相差18 d,其日平均生长速度0.56 cm,与观测值(0.81 cm)相差0.25 cm,其速生阶段为[56.37,272.88],即5月26日至笠年1月2日,在这一时期平均每天生长量约0.46 cm,其生长量占整个生长量的70.73%。同理,理论上地径生长最大时间为111.27 d(7月21日),与观测值90 d(6月30日)相差21 d,其日平均生长速度0.12 mm,与观测值(0.21) mm相差0.09 mm,地径的速生期起点较株高的速生期起点提前47.91 d,其速生阶段为[8.45,231],即4月9日至11月21日,在这一时期平均每天生长量约0.12 mm,其生长量占整个生长量的74.18%,地径的速生期终止点较株高的速生期终止点提前24.88 d。
2.3.2 株高、地径各生长阶段的划分
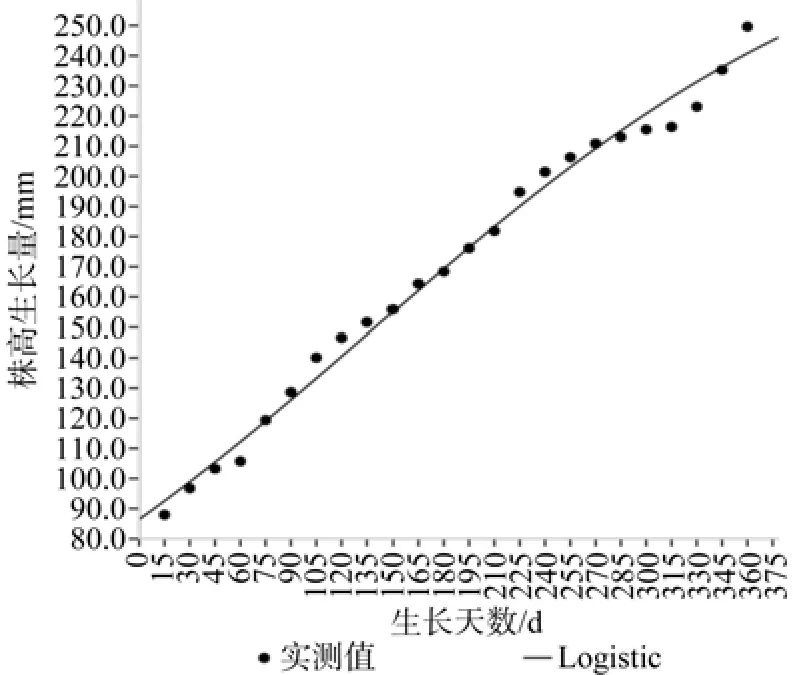
图3 株高生长动态与Logistic 曲线Fig. 3 Growth dynamics of height and its Logistic fi tting curve
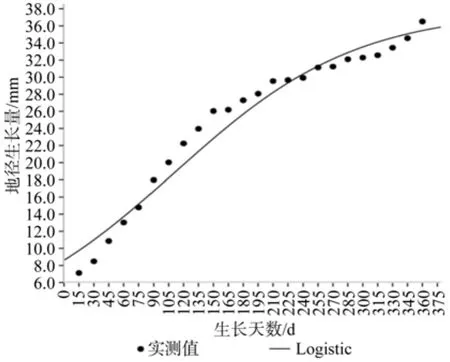
图4 地径生长动态与Logistic 曲线Fig 4 Growth dynamics of basal diameter and its Logistic fi tting curve

表1 株高、地径的Logistic方程及回归假设检验Table 1 Logistic equations of height and basal diameter and the regression test
根据以上分析并结合实际观测值,可以把台湾桤木株高年生长过程划分为3个阶段:第1阶段为渐增期,即从种植开始至5月25日,这期间很短,生长量仅占全年总生长量的10.56%;第2阶段为速生期,从5月26日至翌年1月2日,生长量占全年总生长量的70.73%;第3阶段为缓增期,从1月3日至3月30日,生长量仅占全年总生长量的18.71%。同理,可以把台湾桤木地径年生长过程划分为3个阶段:第1阶段为渐增期,即从种植开始至4月8日,这期间很短,几乎植后恢复即进入速生期,生长量仅占全年总生长量的2.79%;第2阶段为速生期,从4月9日至11月21日,生长量占全年总生长量的74.18%;第3阶段为缓增期,从11月22日至翌年3月30日,生长量仅占全年总生长量的23.03%。

表2 台湾桤木年生长阶段分析及特征值Table 2 Annual growth phases and the characteristics
2.3.3 台湾桤木生物量的积累与分配及其与生长观测指标的相关分析
从表3可知:台湾桤木在广西南宁的年株高平均可达到249.67 cm,地径可达3.65 cm,平均每株含水量可达1 761.3 g,为全株干质量的1.39倍,全株干质量为1 265.37 g,其中地上部分为987.74 g,占全株的78.06%,地下部分为277.63 g,占全株的21.94%,叶和枝的干质量占整个地上干质量的53.95%,大于5 cm的侧根达9条。
对台湾桤木的株高、地径与生物量、含水量进行多元回归分析,从表4、5可以看出:全株干质量、地上干质量、地下干质量、全株含水量、地上部分含水量和地下部分含水量与株高和地径所建立的逐步回归方程均达到显著差异,因此,利用台湾桤木的生长指标株高和地径对其生物量与含水量进行分析与预测是可行的;其中以地上部分含水量和地下部分含水量与株高、地径的相关关系高度相关,其复相关系数达1.000和0.999;其各测定组分与生长指标的偏相关系数看,地径与全株干质量和地上干质量的相关关系比株高更密切,而株高与地下干质量、全株含水量、地上部分含水量和地下部分含水量的相关关系明显比地径更密切,这说明株高对其地下干质量、全株含水量、地上部分含水量和地下部分含水量的贡献率要大于地径,而地径对其全株干质量和地上干质量的贡献率要大于株高。

表3 年生长性状指标及其生物量分配模式Table 3 Annual growth indexes and distribution of biomass
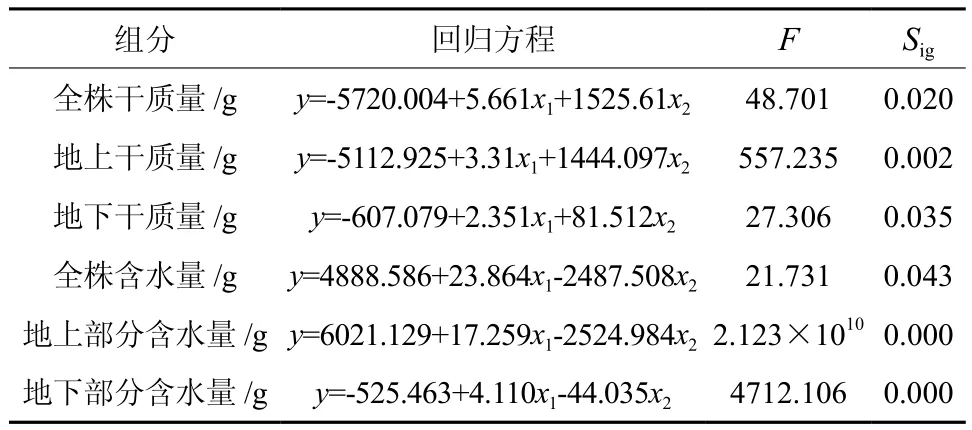
表4 各测定组分与生长指标(株高/x1、地径/x2)的多元回归方程Table 4 Multi-regression of height and basal diameter

表5 各测定组分与生长指标(株高/x1、地径/x2)偏相关分析Table 5 Partial correlation analysis between measurement character and increment of height and basal diameter
3 结论与讨论
台湾桤木的年生长节律遵循Logistic曲线,并且用此方程可以模拟株高和地径的年生长过程,拟合效果理想,据此方程可以预测植株生长期间株高和地径的生长量,并且由此方程导出的最大日平均生长量的出现时间及连日生长量变化速率的2个拐点与实际观测值基本相符,这有助于对台湾桤木生长动态的跟踪和预测。
树木生长规律是树木遗传特性与环境条件相适应的反映,掌握了树木生长规律,可以根据树木生长过程中不同时期的生物学特性来科学管理树木,避免管理上的盲目性,不仅可降低树木成本,同时依据树木的生长规律,人为地施加促生条件,加快树木生长[23]。研究发现,台湾桤木一年生植株的高生长和地径生长均呈“S”形曲线,整个生长经历了一个“慢—快—慢”的过程,生长呈波折状,阶梯性生长明显,具有明显的阶段性,属于全期生长类型。依据台湾桤木的年生长特点,可将其生长过程划分为渐增期、速生期、缓增期三个生长时期。渐增期维持时间较短,株高植后50多天即开始进入(5 月25日)速生期,直到翌年1月2日止,速生期维持时间较长,达217天,地径植后即开始(4月9日)进入速生期,直到11月21日,达223天,这一时期是台湾桤木快速增长期,株高和地径的生长量占全年总生长量的70.73%和72. 67%,由于植株营养分配重心的转移等原因,株高、地径生长均出现了生长暂缓期,这过程也很短,从实际测定中可知,株高、地径至翌年3月1日开始进入渐增至快速增长生长。针对不同的生长阶段,制定不同的栽培技术措施,速生期是植株生长的关键期,速生期的长短决定了其当年生长量,所以在速生期应加强水肥管理,满足植株旺盛生长所需的营养条件,促进树木的快速生长。
台湾桤木的速生期内株高出现明显的速生双峰值,峰期出现在植后90天(即6月1日至6月15日)和植后225天(即11月1日至11月15日),而地径年生长只有 1 次生长高峰期,出现在植后90后(即6月1日至6月15日),这与熊大国等[17]在四川省广元市元坝区研究的 1 年生长节律的结论有点不同,台湾桤木在四川省广元市元坝区生长株高、地径均有2次生长高峰,峰期出现在7月和9月,其至11月中旬树木开始停止生长,12月份树木全部处于休眠状态,而在广西没有休眠期,全年均可生长,只是增长快与慢的区别,是否受到试验地点的自然环境影响,有待进一步研究。
植株生物量积累与株高和地径的相关性极为密切,所建立的苗高、地径与全株生物量、全株含水量回归模型均达到显著水平,其中个别函数模型其复相关系数甚至达到或接近1.0,可利用台湾桤木的生长指标株高和地径对其生物量与含水量进行分析与预测;植株含水量丰富,株含水量为全株干质量的1.39倍,叶和枝的干质量占整个地上干质量的53.95%,大于5 cm的侧根达9条。通过对台湾桤木年生长特性的研究,并对其生长期进行划分,为台湾桤木栽培管理提供了理论依据,但适宜台湾桤木栽培的不同地区其气候条件有所差异,本研究中的台湾桤木年生长特性和生长阶段划分是依据广西南宁地区的气候特征而指出的,并不能完全代表其它气候区的划分标准。
[1] 王金锡, 朱万泽. 台湾桤木生态生物学特性及引种推广前景[J]. 四川林业科技, 2000, 21(4): 16-19.
[2] Huang Yaw-fuh, Haw-Farn. Studies on the manufacture of hardboard from fast-growing wood species (II). Malacca albizzia,Formosan sweet gum, Formosan alder and India-charcoal trema as raw materials for hardboard manufacturing [J]. Quarterly Journal of Chinese Forestry, 1994, 27 (3):119-139.
[3] 林信辉, 张俊兵. 中横崩塌地植被特性及其优势植物主要生理反应之研究[J]. 中华水土保持学报, 1995, 26 (1): 1-6.
[4] Lai Y C, Chen C K, Lin W W, et al. A comprehensive investigation of anti-inflammatory diarylheptanoids from the leaves of Alnus formosana[J]. Phytochemistry, 2012,73(1):84-94
[5] 潘 燕. 不同施肥对台湾桤木根系生长和衰老生理的影响研究[D]. 成都:四川农业大学, 2008.
[6] 朱万泽, 王金锡, 薛建辉. 台湾桤木引种气候生态适生区分析[J]. 热带亚热带植物学报, 2005, 13(1): 59-64.
[7] 朱积余, 梁瑞龙, 蒋 燚. 广西优良珍贵树种发展的现状、问题与对策[J]. 广西林业科学, 2007, 36(1): 1-4.
[8] 廖德志, 李志辉, 吴际友, 等. 台湾桤木无性系扦插繁殖试验[J]. 湖南林业科技, 2010, 37(6): 48-51.
[9] 邹高顺. 台湾桤木引种造林及培肥能力研究[J]. 福建林学院学报, 1995, 16(2): 112-117.
[10] 王军辉, 顾万春, 夏良放, 等. 四川省桤木种源单株生物质产量的遗传变异和选择初探[J]. 林业科学, 2005, 41(4): 55-61.
[11] 朱万泽, 王金锡, 薛建辉. 台湾桤木引种的光合生理特性研究[J]. 西北植物学报, 2004, 24(11): 2012-2019.
[12] 周小玲, 田大伦, 许忠坤, 等. 中亚热带四川桤木与台湾桤木幼树的光合生态特性[J]. 中南林业科技大学学报, 2007,27(1): 40-49,54.
[13] 王旭军, 吴际友, 程 勇, 等. 台湾桤木花粉生活力的测定方法初探[J]. 林业科学, 2007, 23(9): 201-20.
[14] 朱万泽, 薛建辉, 王金锡. 台湾桤木种源对水分胁迫的光合响应及其抗旱性[J]. 水土保持学报, 2004, 18(4): 170-173, 181.
[15] 卓仁英, 陈益泰. 五种桤木属植物的光合特性初步研究[J].浙江林业科技, 2004, 24 (6): 1-4.
[16] Weng J H, Chen Y N, Liao T S. Relationship between chlorophyll fluorescence parameters and photochemical reflectance index of tree species adapted to different temperature regimes [J].Functional Plant Biology, 2006, 33 (3): 241-246.
[16] 朱万泽, 王金锡. 引种台湾桤木的蒸腾特性及其影响因子[J].四川林业科技, 2005, 26(1): 9-14.
[17] 陈建华, 纪程灵, 吴际友, 等. 台湾桤木无性系的生长性状比较[J]. 中南林业科技大学学报, 2012, 32 (4): 7-9,31.
[18] 熊大国, 王海琼, 梁红梅, 等. 元坝区台湾桤木年生长节律与生物量研究[J]. 四川林业科技, 2006, 27 (6): 55-58.
[19] 殷祚云. Logistic曲线生长研究[J]. 数理统计与管理, 2002,(1): 41-46.
[20] 庞世龙, 黄志玲, 郝海坤, 等. 大叶栎苗期生长模型及分析[J]. 林业科技开发, 2011, 25(5): 36-39.
[21] 黄志玲, 庞世龙, 郝海坤, 等. 广西珍贵乡土速生树种任豆苗木生长节律研究[J]. 西南林业大学学报, 2011, 31(6): 6-9.
[22] 刘 磊. 逻辑增长曲线模型的一种参数估计方法[J]. 湖北工业大学学报, 2008, 23(1): 83-84,94.
[23] 包 珩. 墨西哥柏幼苗生长规律及光合特性研究[D]. 南京:南京林业大学, 2012.
Analysis on growth rhythm and biomass increment of an introduced variety of Alnus formosana
PENG Yu-hua1,2,3, HUANG Zhi-ling1,2,3, SHEN Wen-hui1,2,3, ZHU ji-yu1,2,3, HAO Hai-kun1,2,3
(1.Guangxi Forestry Research Institute; 2. Central South Key Lab. of Fast-growing Tree Cultivation Attached to Forestry Ministry;3.Guangxi Key Lab. of Superior Timber Trees Cultivation. Nanning 530002 ,Guangxi,China)
∶ Annual growth dynamics of an introduced variety of Alnus formosana were investigated, as well as its biomass accumulation and distribution, and the correlation between growth indexes and biomass components. The results showed that the growth patterns were consistent with Logistic curve, and the derived models of height, basal diameter and time demonstrated signif i cant consistency (R2>0.94).Based on two points of the greatest curvature on the curve, plant growth processes could be divided into 3 periods: cumulating, fastgrowing and moderate growth. During the fast-growing period, both the increments of height and basal diameter were more than 70%of the total year increments. The biomass increase was closely correlated with height and basal diameter, and the aboveground biomass(dry weight) accounted for 78.06% of a plant, branches and leaves together weighted 53.95% of the aboveground biomass, the moisture content was 1.39 times of the dry-mass of a plant. The lateral roots (>5 mm) averaged 9 for one plant. This analysis may inform the cultivation and domestication of introduced Alnus formosana in Guangxi.
∶ Alnus formosana; growth rhythm; Logistic curve; biomass; moisture content
S792.94
A
1673-923X(2014)06-0007-06
2013-09-09
广西林业科技项目“台湾桤木引种与区域试验示范”(桂林科字[2012]第12号);国家发改委重大项目“广西主要珍贵树种种质资源收集库建设项目”(桂发改农经[2010]599号)
彭玉华(1963-),女,工程师,研究方向:森林培育与生态林业工程研究;E-mail:pyh112233456789@126.com
[本文编校:吴 彬]
猜你喜欢
农业灾害研究(2022年2期)2022-05-31
南方农业·下旬(2022年4期)2022-05-24
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
农村百事通(2019年17期)2019-10-08
林业与生态(2019年6期)2019-07-02
现代园艺(2018年1期)2018-03-15
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
