
红椿幼苗对干旱胁迫及复水生理响应的典型相关分析
2015-01-07 07:26李志辉吴际友陈彩霞吴志华黄明军
西北农林科技大学学报(自然科学版) 2015年10期
刘 球,李志辉,吴际友,陈彩霞,李 艳,吴志华,程 勇,黄明军
(1 中南林业科技大学 林学院,湖南 长沙 410004;2 湖南省林业科学院,湖南 长沙 410004;3 国家林业局桉树研究开发中心,广东 湛江 524022 )
红椿幼苗对干旱胁迫及复水生理响应的典型相关分析
刘 球1,2,李志辉1,吴际友2,陈彩霞1,李 艳1,吴志华3,程 勇2,黄明军1
(1 中南林业科技大学 林学院,湖南 长沙 410004;2 湖南省林业科学院,湖南 长沙 410004;3 国家林业局桉树研究开发中心,广东 湛江 524022 )
【目的】 探求干旱胁迫和复水条件下红椿盆栽幼苗叶片生理特性的整体相关性,为深入研究红椿的抗旱性奠定基础。【方法】 在人工遮雨棚中对2年生红椿盆栽幼苗进行干旱胁迫及复水试验,测定叶片相对含水量、相对电导率、叶绿素含量、SOD活性、POD活性、MDA含量、游离脯氨酸含量及可溶性蛋白含量8个叶片生理指标,并运用典型相关法对未干旱(干旱胁迫前)、干旱胁迫及复水条件下3组指标两两之间的整体相关性进行分析。【结果】 (1)干旱胁迫前与干旱胁迫条件下,红椿盆栽幼苗叶片生理特性之间存在显著相关关系,干旱胁迫前及干旱胁迫下红椿幼苗生理响应典型相关分析的第1对典型变量(U1,V1)的相关系数达到0.996 5,叶片相对电导率可以反映干旱胁迫前的生理特性,叶绿素含量可以反映干旱胁迫的生理特性。(2)干旱胁迫与复水后红椿盆栽幼苗叶片生理特性有显著相关关系,干旱胁迫及复水条件下红椿幼苗生理响应典型相关分析的第1对典型变量(W1,Z1)的相关系数达到0.998 3,反映干旱胁迫和复水后生理特性的典型变量均主要由叶片相对电导率决定。(3)干旱胁迫下叶绿素含量与复水时的POD活性和MDA含量均呈显著正相关关系,说明干旱胁迫下红椿幼苗光合作用的保持能力强,则复水时表现出来的抗氧化酶活性也高。(4)干旱胁迫下可溶性蛋白含量与复水后的POD活性呈极显著正相关关系,相关系数达到0.803,说明复水时红椿幼苗植株叶片生理特性的恢复状况与干旱胁迫时受创的严重程度密切相关。(5)干旱迫胁前与复水后叶片生理指标的相关性表明,复水后红椿幼苗植株的耗水量、抗旱性与正常供水时植株的抗旱潜力直接相关。【结论】 干旱胁迫伤害程度不重,则对复水后红椿幼苗植株体内叶绿素含量和POD活性的恢复具有积极作用。适当干旱能促进植株抗逆性的提高,过度缺水则会对植株造成伤害,且修复慢,符合生物节水原理。
干旱胁迫;红椿;复水;生理响应;生物节水;典型相关分析
水分是植物生存的必要条件,植物在干旱胁迫条件下生存生长,必须有相应的水分利用策略[1]。目前,在全球气候变化的大背景下,我国一些地区陆续出现极端持续高温天气,而这种天气给植物的正常生长带来了巨大的挑战。湖南地处大陆性中亚热带气候区,在2012年夏季经历了持续50多天的极端干旱天气,许多植物遭受了不同程度的伤害。因此,在湖南开展植物抗旱性研究就显得尤为必要。
红椿(ToonacilliateRoem.)属楝科香椿属半常绿乔木,国家Ⅱ级重点保护植物,是我国热带、亚热带地区特有的珍贵速生用材树种。红椿木材红褐色,花纹美丽,质地坚硬,密度高,为上等家具用材,素有“中国桃花心木”之美誉[2]。湖南省是红椿天然分布省份之一,虽然只是天然零星分布,但发展潜力巨大。目前红椿已成为我国南方低山地区重要的速生用材林造林树种和营造针阔混交林进行树种结构调整的首选树种之一[3]。
国内对红椿已经开展了许多研究,包括形态特征、生物学特性、生长规律、育苗技术及造林技术等[4-7]。近年来,湖南省林业科学院红椿科研团队开始不断发掘红椿在抗逆方面的潜力,已经发表多篇有关红椿对水分胁迫的生理生态响应的研究报道[8-11]。本研究对干旱及复水条件下红椿叶片的生理特性指标进行测定,应用典型相关分析方法,分析干旱胁迫前、干旱胁迫和复水条件下红椿3组生理指标两两之间的整体相关性及生理指标的整体关系,深入研究红椿幼苗在不同水分环境条件下的水分生理状况及相互关系,以期为进一步科学合理地进行红椿人工林培育提供理论基础和指导。
1 材料与方法
1.1 试验地概况
试验地位于湖南省林产品质量检验检测中心苗圃内,地理坐标为东经112°59′、北纬28°05′,年平均气温16.8 ℃,平均日照时数1 496~1 850 h,年降水量1 400~1 900 mm,无霜期264 d,属中亚热带季风湿润气候区,光照充足,雨量丰沛;海拔为110 m,土壤为砂岩发育的红壤,土层厚达60 cm以上;土壤肥力中等,pH值为6.2。
1.2 试验材料
本试验以2年生红椿盆栽幼苗为研究对象。2012-03于桃源县白家育苗圃统一育苗并进行田间管理;2014-03选择长势均匀、健康的红椿幼苗30株,于湖南省林产品质量检验检测中心苗圃完成入盆,盆钵口径25 cm,深20 cm,每盆装土5 kg(V(黄心土)∶V(泥炭土)=2∶1),配统一盆垫,正常浇水管护;2014-06,选择18株长势均匀的红椿盆栽幼苗布置干旱胁迫试验,受试盆栽苗苗高平均1.0 m,胸径1.8 cm。
1.3 试验设计
试验采用完全随机设计,每重复6盆幼苗,重复3次,共18盆幼苗。为确保试验基础条件基本一致,于2014-06-04-2014-06-06对试验苗木断水3 d,06-07-06-09为受试植株每盆每天浇水500 mL,以保证基本一致的试验起始条件。2014-06-10正式开始干旱胁迫,于06-20干旱胁迫结束并开始复水,3 d复水1次,06-29复水结束。整个干旱胁迫及复水试验从2014-06-10持续到06-29,共19 d。干旱胁迫试验在人工遮雨棚中进行,保证正常的光照条件,且空气流通顺畅,棚中温度与室外基本一致。
1.4 指标测定及方法
未干旱(干旱胁迫前)、干旱胁迫及复水处理红椿叶片的采样时间分别是06-10、06-20和06-29,每次采样均在早上08:00-09:00进行,每次采取每株幼苗同一轮成熟叶片2~3片作为试验测定样品,用冰块保鲜,迅速带回实验室进行指标测定,重复测定3次取平均值。
叶片生理指标测定参照文献[12]的方法,其中叶片相对含水率的测定采用烘干法,质膜相对透性测定采用电导率法,叶绿素含量测定采用浸提法,丙二醛(MDA)含量测定采用硫代巴比妥酸法,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑法,游离脯氨酸含量测定采用酸性茚三酮法。过氧化物酶(POD)活性和可溶性蛋白含量测定采用陈建勋等[12]建立的方法。
1.5 数据处理及统计分析
1.5.1 数据处理 (1)干旱胁迫前与干旱胁迫条件下红椿盆栽幼苗叶片生理特性典型变量。将数据分成2类,即干旱胁迫前生理指标和干旱胁迫生理指标,干旱胁迫前生理指标包括干旱处理前叶片的相对含水量、相对电导率、叶绿素含量、SOD活性、POD活性、MDA含量、游离脯氨酸含量、可溶性蛋白含量,分别用子变量x1、x2、x3、x4、x5、x6、x7、x8表示,从而构成因变量U;干旱胁迫生理指标包括干旱胁迫后叶片的相对含水量、相对电导率、叶绿素含量、SOD活性、POD活性、MDA含量、游离脯氨酸含量、可溶性蛋白含量, 分别用子变量y1、y2、y3、y4、y5、y6、y7、y8表示,从而构成因变量V。采用典型相关分析研究2类变量(U,V)间的相关性。
(2)干旱胁迫与复水条件下红椿盆栽幼苗叶片生理特性典型变量。将数据分成2类,即干旱胁迫生理指标和复水后生理指标。干旱胁迫生理指标包括干旱胁迫后叶片的相对含水量、相对电导率、叶绿素含量、SOD活性、POD活性、MDA含量、游离脯氨酸含量、可溶性蛋白含量,分别用子变量x1′、x2′、x3′、x4′、x5′、x6′、x7′、x8′表示,从而构成因变量W;复水后生理指标包括复水后叶片的相对含水量、相对电导率、叶绿素含量、SOD活性、POD活性、MDA含量、游离脯氨酸含量、可溶性蛋白含量,分别用子变量y1′、y2′、y3′、y4′、y5′、y6′、y7′、y8′表示,从而构成因变量Z。采用典型相关分析研究2类变量(W,Z)间的相关性。
1.5.2 数据统计分析 运用Excel 2007进行数据整理,并运用DPS 7.05进行典型相关分析。
2 结果与分析
2.1 干旱胁迫前与干旱胁迫条件下红椿盆栽幼苗叶片生理特性的典型相关分析
从表1可以看出,第1对典型变量的相关系数达到0.996 5,而在P=0.05的显著性水平下,8对典型变量中只有第1组典型相关系数达到显著水平,其余7对相关性均不显著。因此,可以取第1对典型变量来分析2类性状之间的相互关系。
用2组典型变量的系数建立典型模型,其中对干旱前生理特性的第1典型变量,有:
U1=0.341x1-0.578 9x2+0.019 3x3-0.157 1x4-0.473 2x5-0.044x6-0.386 1x7-0.484 1x8;
对干旱胁迫生理特性的第1典型变量,有:
V1=0.412 1y1-0.078 4y2-0.870 1y3-0.212y4-0.539y5-0.279 6y6-0.359 8y7-0.312y8。
来自干旱胁迫前生理特性的第1典型变量U1在x2上载荷值的绝对值最大,其次是x8和x5,反映干旱前生理特性的典型变量主要由叶片相对电导率决定,其次是可溶性蛋白含量和POD活性。各生理特性指标对第1典型变量U1的贡献大小顺序为:x2(干旱前叶片相对电导率)>x8(干旱前可溶性蛋白含量)>x5(干旱前POD活性)>x7(干旱前游离脯氨酸含量)>x1(干旱前叶片相对含水量)>x4(干旱前SOD活性)>x6(干旱前MDA含量)>x3(干旱前叶绿素含量),其中除x1和x3所起作用为正相关外,其他均为负相关,且x3系数小于0.1,作用最小。说明植株水分供给充分时叶片相对含水量较高,而水分供给充分与否直接影响叶片生理性状,这与客观事实相符。

表1 干旱胁迫前与干旱胁迫条件下红椿盆栽幼苗叶片生理特性典型相关分析时相关系数的显著性检验Table 1 Significance test on coefficients of canonical correlation analysis on leaf physiological characteristics of potted Toona cilliate Roem.seedlings among control and drought stress
来自干旱胁迫生理特性的第1典型变量V1在y3上载荷值的绝对值最大,其次为y5,说明反映干旱胁迫生理特性的典型变量主要由叶绿素含量决定,其次是POD活性。各生理特性指标对第1典型变量V1的贡献大小顺序为:y3(干旱胁迫叶绿素含量)>y5(干旱胁迫POD活性)>y1(干旱胁迫叶片相对含水量)>y7(干旱胁迫游离脯氨酸含量)>y8(干旱胁迫可溶性蛋白含量)>y6(干旱胁迫MDA含量)>y4(干旱胁迫SOD活性)>y2(干旱胁迫叶片相对电导率),其中除y1为正贡献外,其他指标均为负贡献,且y3贡献尤其突出,y2系数小于0.1,作用最小。说明干旱胁迫导致叶片叶绿素含量显著下降,植株体内抗氧化酶活性等指标也发生变化,如POD活性贡献排名第二,说明干旱胁迫导致植株体内产生的大量活性氧物质已造成伤害,POD活性的增加是为了清除植株体内的活性氧,是植株遭受干旱胁迫时体内抗氧化酶的一种应激适应反应。
从典型冗余分析可以看出,第1组典型变量U1可以解释13.57%的组内变异,并解释另一组19.53%的变异;而典型变量V1可以解释19.67%的组内变异,并解释另一组13.48%的变异。
2.2 干旱胁迫与复水条件下红椿盆栽幼苗叶片生理特性的典型相关分析
从表2可以看出,第1对典型变量的相关系数达到0.998 3。统计检验显示,在P=0.05的显著性水平下,8对典型变量中只有第1组典型相关系数达到显著水平,其余7对相关性均不显著。因此,可以取第1对典型变量来分析2类性状之间的相互关系。
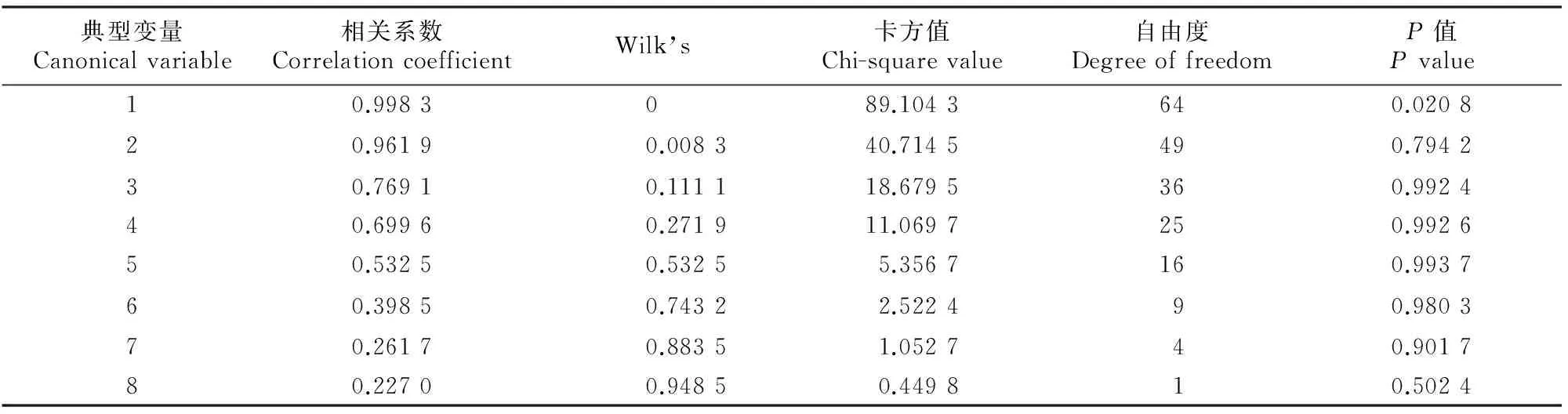
表2 干旱胁迫与复水条件下红椿盆栽幼苗叶片生理特性典型相关分析时相关系数的显著性检验Table 2 Significance test on coefficients of canonical correlation analysis on leaf physiological characteristics of potted Toona cilliate Roem.seedlings under drought stress and rewatering
用2组典型变量的系数建立典型模型,其中对干旱胁迫生理特性的第1典型变量,有:
W1=-0.263 5x1′-0.754 1x2′-0.291 6x3′+0.454 5x4′-0.140 8x5′-0.348 7x6′-0.029 2x7′-0.275 3x8′;
对复水后生理特性的第1典型变量,有:
Z1=0.082y1′-0.487 4y2′-0.086 1y3′-0.123 1y4′-0.426y5′-0.261 2y6′+0.180 6y7′+0.393 9y8′。
来自干旱胁迫生理特性的第1典型变量W1在x2上载荷值的绝对值最大,反映干旱胁迫生理特性的典型变量主要由叶片相对电导率决定。各生理特性指标对第1典型变量W1的贡献大小顺序为:x2′(干旱胁迫叶片相对电导率)>x4′(干旱胁迫SOD活性)>x6′(干旱胁迫MDA含量)﹥x3′(干旱胁迫叶绿素含量)>x8′(干旱胁迫可溶性蛋白含量)>x1′(干旱胁迫叶片相对含水量)>x5′(干旱胁迫POD活性)>x7′(干旱胁迫游离脯氨酸含量),其中除x4′所起作用为正相关外,其他均为负相关,且x7′系数小于0.1,作用最小。说明植株受干旱胁迫时,叶片相对电导率的变化为生理指标变化的最直观体现。植株在受到胁迫时,膜系统遭受破坏,体内渗透失衡,从而导致相对电导率发生明显变化,符合客观事实。
来自复水后生理特性的第1典型变量Z1在y2′上载荷值的绝对值最大,反映复水后生理特性的典型变量主要由叶片相对电导率决定,这与干旱胁迫时生理特性的变化保持一致;各生理特性指标对第1典型变量Z1的贡献大小顺序为:y2′(复水后叶片相对电导率)>y5′(复水后POD活性)>y8′(复水后可溶性蛋白含量)>y6′(复水后MDA含量)>y7′(复水后游离脯氨酸含量)>y4′(复水后SOD活性)>y3′(复水后叶绿素含量)>y1′(复水后叶片相对含水量),其中除y1′、y7′和y8′为正贡献外,其他指标均为负贡献,且y1′、y3′系数小于0.1,作用很小可忽略不计。复水后生理特性的第1典型变量中,叶片相对电导率系数较其在干旱胁迫生理特性的第1典型变量中减小了约35%,表示胁迫造成的渗透失衡有所恢复,说明复水使植株所受伤害有所缓解;然而其贡献值仍然排在第1位,并未恢复到干旱前水平,说明复水虽然在一定程度上缓解了干旱胁迫的伤害,但是短时内并未修复彻底,伤害完全修复尚需更长的时间和过程。
2.3 红椿盆栽幼苗叶片生理指标在3种试验条件下两两之间的相关关系分析
2.3.1 正常供水(干旱前)与干旱胁迫 如表3所示,干旱前生理指标中,红椿的可溶性蛋白含量与SOD活性的相关系数达到0.885,呈极显著正相关关系。干旱胁迫生理指标中,可溶性蛋白含量与叶绿素含量的相关系数达0.542,说明在干旱胁迫下,可溶性蛋白含量的变化与叶绿素含量的变化呈显著正相关趋势,叶片因缺水导致的光合作用受阻可能直接影响叶片生理性状的变化。
干旱胁迫时的叶绿素含量与干旱前叶片相对电导率以及POD活性的相关系数分别达0.478和0.507,说明红椿植株在正常供水条件下的叶片相对电导率和POD活性与干旱胁迫下叶片叶绿素含量的变化具有正相关关系。
2.3.2 干旱胁迫与复水 如表4所示,干旱胁迫条件下,红椿幼苗叶片各生理指标之间仅可溶性蛋白与叶绿素含量达到显著正相关关系,相关系数为0.542。复水条件下,MDA含量与叶绿素含量呈显著正相关,相关系数为0.524,复水后植株叶片回绿需要一段时间,叶绿素含量增加不大时,MDA含量变化亦不大;MDA含量与POD活性呈极显著正相关,相关系数为0.666,复水使植株胁迫伤害有所修复,因而叶片POD活性大幅度下降时,MDA含量也随着明显下降,这符合正常规律。MDA含量为脂质过氧化指标,一般直接用来反映植株的受伤程度[11]。
干旱胁迫时叶片的叶绿素含量与复水条件下的POD活性和MDA含量均呈显著正相关关系,说明植株在干旱胁迫下叶绿素含量越高,即其在逆境下保持光合作用的能力越强,而经过了干旱胁迫这个过程后,复水时表现出来的抗氧化酶活性也较高,即一定程度的干旱有助于进一步挖掘抗旱潜力较高植株的抗旱性能;干旱胁迫下可溶性蛋白含量与复水条件下的叶绿素含量和POD活性呈极显著正相关,其中与后者的相关系数达到0.803,同时与MDA含量也达到显著相关。干旱胁迫后复水时植株叶片生理特性的恢复情况与干旱胁迫时各生理指标受创的严重程度直接相关。植株在遭受干旱胁迫时,可溶性蛋白含量会有所下降[11]。干旱胁迫越严重,则可溶性蛋白含量越低。因此复水后叶绿素含量和POD活性越低,说明干旱胁迫越严重,对植株造成的伤害就越大,复水后修复的效率也越低。同时也说明,过度干旱胁迫无法提高植株的抗旱性,反而会对植株造成更大的伤害。

表3 红椿盆栽幼苗干旱胁迫前与干旱胁迫下叶片生理指标的相关系数Table 3 Correlation coefficients of leaf physiological indicators of potted Toona cilliate Roem. seedlings between control and drought stress
注:“*”表示在α=0.05时具有显著相关性;“**”表示在α=0.01时具有极显著相关性。下表同。
Note:“*” shows significant correlation at the level ofα=0.05;“**” shows extremely significant correlation at the level ofα=0.01.The same below.
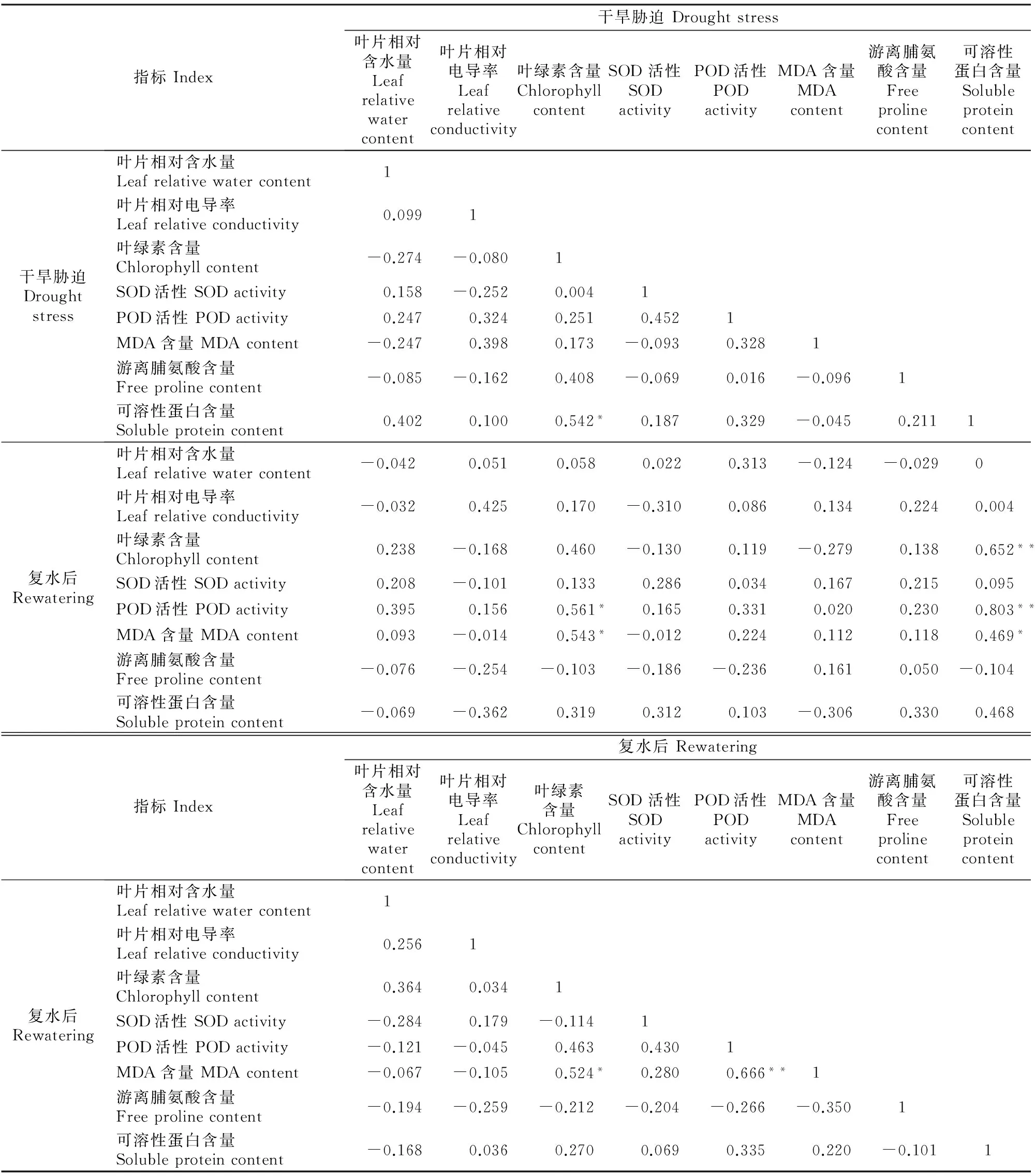
表4 红椿盆栽幼苗干旱胁迫与复水后叶片生理指标的相关系数Table 4 Correlation coefficients of leaf physiological indicators of potted Toona cilliate Roem. seedlings between drought stress and rewatering
2.3.3 正常供水(干旱前)与复水 如表5所示,复水后的叶片相对含水量与干旱胁迫前的SOD活性、可溶性蛋白含量呈显著正相关;复水后的POD活性、MDA含量与干旱胁迫前的POD活性呈显著正相关;复水后的游离脯氨酸含量与干旱胁迫前的游离脯氨酸含量呈显著正相关;复水后的可溶性蛋白含量与干旱胁迫前的MDA含量呈显著负相关。说明干旱胁迫后复水时植株的耗水量、抗旱性与正常供水植株的抗旱潜力直接相关。因此,可在正常供水条件下就植株个体差异进行优良单株选择,从而为高抗旱性红椿植株的选育提供参考。
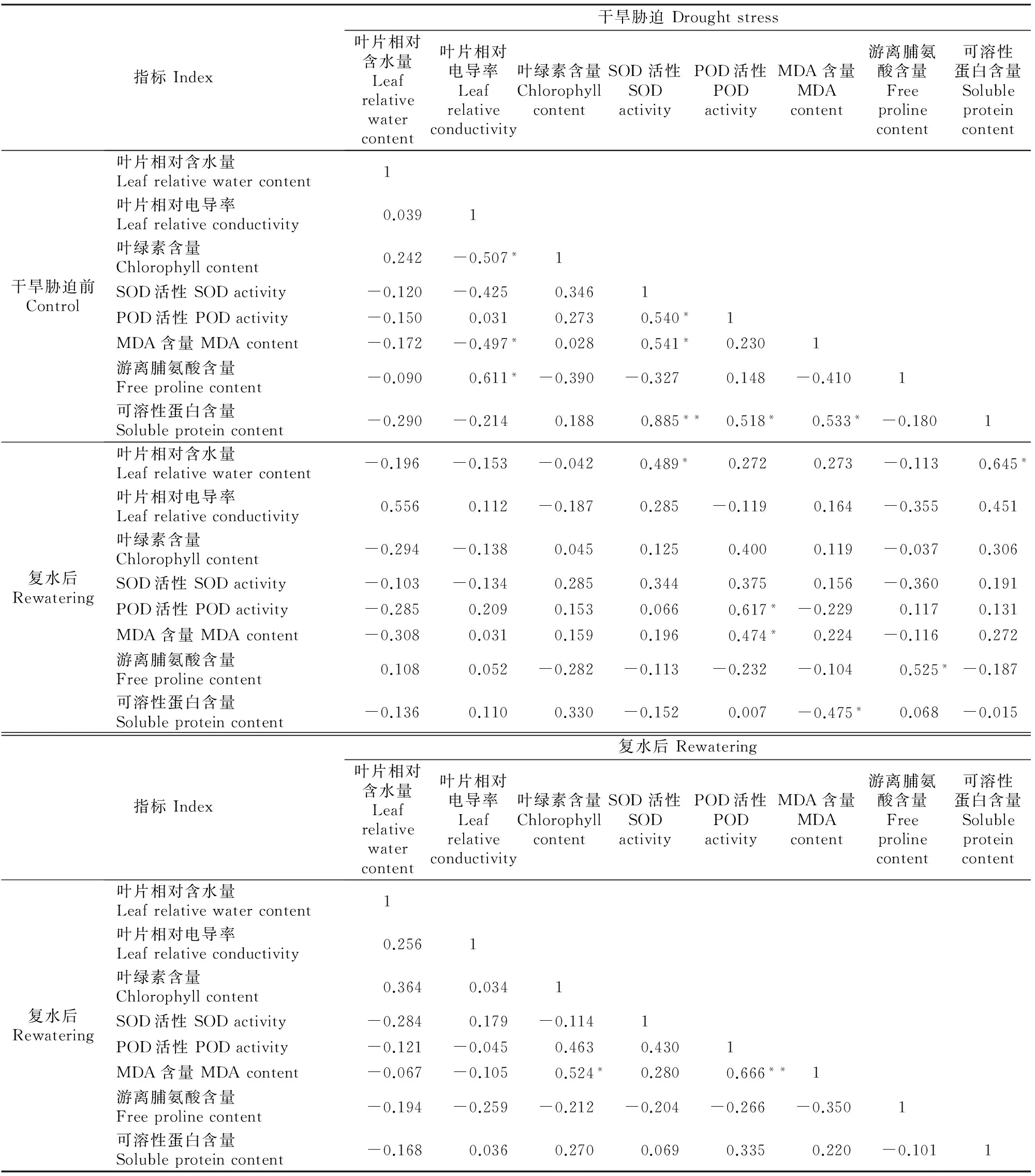
表5 红椿盆栽幼苗干旱胁迫前生理指标与复水后生理指标的相关系数Table 5 Correlation coefficients of leaf physiological indicators of potted Toona cilliate Roem.seedlings between contrast and rewatering
3 讨论与结论
3.1 3种水分条件下红椿幼苗叶片生理特性的典型相关分析
在正常供水与干旱胁迫条件下红椿盆栽幼苗叶片生理特性的典型相关分析中,干旱胁迫生理特性的第1典型变量表明,植株在受到干旱胁迫时,叶片叶绿素含量的下降贡献最大,该结果符合正常规律,因为没有水分供给,植株因无法进行正常的生长活动而造成伤害,包括叶绿素含量降低,这与吴际友[13]的研究结论一致。同时,抗氧化酶活性等生理指标亦发生响应变化,如POD活性提高。有研究表明,POD、SOD、CAT等抗氧化酶活性在水分胁迫下有增加趋势[14-15]。植物在逆境下,体内的活性氧代谢系统失衡,导致大量活性氧物质积累,从而对植株造成伤害。而超氧化物歧化酶(SOD)可消除植株体内的超氧自由基因子,将植株体内有害的超氧自由基转化为过氧化氢,再通过过氧化物酶POD和过氧化氢酶CAT将其分解为水。而当植株受到严重胁迫时,其体内产生的活性氧物质超出了抗氧化酶系统所能承受的范围,导致抗氧化酶活性降低,植株氧化加速[16]。
在干旱胁迫及复水红椿盆栽幼苗叶片生理特性的典型相关分析中,干旱胁迫以及复水后生理特性的第1典型变量一致表明,反映各时期生理指标的典型变量均主要由叶片相对电导率决定。细胞膜不仅是区分细胞内部和外部环境的动态屏障,更是细胞内外物质交换和信息传递的通道。在逆境胁迫时,膜结构受伤导致膜功能受损,使得膜透性增大,细胞内水溶性物质开始外渗,从而相对电导率提高,MDA的积累也相应增加。相对电导率越高,膜相对透性越大,则表示细胞膜在逆境下受伤害程度越大[17]。
复水能使植株的受伤害程度有所缓解。然而,复水虽然能在一定程度缓解已经造成的干旱胁迫伤害,但是伤害完全修复尚需更长的时间和过程,这与有关研究[10]的观点一致。刘锦春等[18]对柏木“干旱-复水”环境适应机制的研究表明,干旱胁迫初期复水后,MDA含量可以下降到对照水平,而干旱胁迫中期和末期后复水则不能降至对照水平。
干旱胁迫下叶绿素含量越高即表明植株在逆境下保持光合作用的能力越强,则在复水时表现出来的抗氧化酶活性越高,即一定程度的干旱有利于进一步挖掘抗旱潜力较高植株的抗旱性;同时也说明,过度的干旱胁迫不但无法提高植株抗旱性,反而会造成更大的伤害。这与山仑院士的生物节水原理不谋而合:山仑等[19]指出,水分亏缺不等于完全对植物产生危害,而是植物从适应干旱到产生伤害的一个过程,这就是生物节水原理;他认为,适度缺水后复水有可能会促进植物生长。
3.2 红椿幼苗叶片生理指标在3种水分条件下两两之间的相关关系
复水时植株的耗水量、抗旱性与正常供水条件下植株的抗旱潜力直接相关。若干旱胁迫伤害程度不重,则对复水时植株体内的叶绿素含量和POD活性恢复具有积极作用,适当的干旱能促进植株抗逆性的提高。一些相关研究也得出过类似的结论,如刘吉利等[20]研究发现,苗期适度干旱可减少花生的耗水量,提高单位产量的水分利用效率;闫永銮等[21]指出,冬小麦拔节期适宜的轻度干旱胁迫有利于提高植株整体的抗旱能力,减少耗水量,明显提高水分利用效率。
综上所述,适当的水分亏缺有利于植株抗逆性的提高,而过度缺水则会对植株造成伤害,且恢复速度较慢、耗时较长。因此,挖掘植物抗旱潜力、精确运用植物水分利用规律并完善干旱预防机制,较寻找植株遭受干旱胁迫后的伤害修复措施更具有现实意义。
[1] 林 雯,何 茜,苏 艳,等.干旱胁迫对欧洲云杉水分生理特征的影响 [J].西北农林科技大学学报:自然科学版,2014,42(6):1-9.
Lin W,He Q,Su Y,et al.Effects of drought stress on water physiological characteristics ofPiceaabies[J].Journal of Northwest A&F University:Nat Sci Ed.,2014,42(6):1-9.(in Chinese)
[2] 中国树木志编委会.中国主要树种造林技术 [M].北京:中国林业出版社,1981:613-615.
China Sylva Editorial Board.Afforestation technology of main tree species in China [M].Beijing:China Forestry Publishing House,1981:613-615.(in Chinese)
[3] 姜 芸,吴际友,姚贤清,等.湖南省珍贵乡土用材树种研究现状与发展对策 [J].湖南林业科技,2013,40(3):1-4.
Jiang Y,Wu J Y,Yao X Q,et al.Research and development countermeasures on precious indigenous timber tree species in Hunan Province [J].Hunan Forestry Science and Technology,2013,40(3):1-4.(in Chinese)
[4] 邹高顺.珍贵速生树种红椿与毛红椿引种栽培研究 [J].福建林学院学报,1994,14(3):271-276.
Zou G S.Study on introduction and cultivation of precious and fast-growing tree speciesToonaciliatevar.pubescens [J].Journal of Fujian College of Forestry,1994,14(3):271-276.(in Chinese)
[5] 吴莉莉,王鸣凤,陈柏林.红椿树的生物学特性及人工栽培试验研究 [J].安徽农学通报,2006,12(7):168-169.
Wu L L,Wang M F,Chen B L.Study on biological characteristics and artificial cultivation test ofToonaciliate[J].Anhui Agri Sci Bull,2006,12(7):168-169.(in Chinese)
[6] 赵汝玉,李光友,徐建民.红椿育苗及造林技术 [J].广西林业科学,2005,34(3):155-156.
Zhao R Y,Li G Y,Xu J M.Cultivation and afforestation technology ofToonaciliate[J].Guangxi Forestry Science,2005,34(3):155-156.(in Chinese)
[7] 文卫华,吴际友,陈明皋,等.红椿优树子代苗期生长表现 [J].中国农学通报,2012,28(34):36-39.
Wen W H,Wu J Y,Chen M G,et al.Seedling growth performance ofToonaciliateelite trees progeny [J].Chinese Agricultural Science Bulletin,2012,28(34):36-39.(in Chinese)
[8] 陈彩霞,王瑞辉,吴际友,等.持续干旱条件下红椿无性系幼苗的生理响应 [J].中南林业科技大学学报:自然科学版,2013,33(9):46-49.
Chen C X,Wang R H,Wu J Y,et al.Physiological responses ofToonaciliateclone seedlings under continuous drought conditions [J].Journal of Central South University of Forestry & Technology,2013,33(9):46-49.(in Chinese)
[9] 吴际友,李志辉,刘 球,等.干旱胁迫对红椿无性系幼苗叶片相对含水量和叶绿素含量的影响 [J].中国农学通报,2013,29(4):19-22.
Wu J Y,Li Z H,Liu Q,et al.Response of drought stress on relative water content of leaf and cholorophyll content of youngToonaciliateRoem.clone seedlings [J].Chinese Agricultural Science Bulletin,2013,29(4):19-22.(in Chinese)
[10] 刘 球,陈彩霞,吴际友,等.红椿无性系幼苗叶片抗氧化酶指标对干旱胁迫的响应 [J].中南林业科技大学学报:自然科学版,2013,33(11):73-76.
Liu Q,Chen C X,Wu J Y,et al.Response of drought stress on anti-oxydant enzyme in leaves of youngToonaciliateclone seedlings [J].Journal of Central South University of Forestry & Technology:Nat Sci Ed,2013,33(11):73-76.(in Chinese)
[11] 陈彩霞,王瑞辉,吴际友,等.淹水胁迫对红椿家系幼苗几种生理生化指标的影响 [J].西北林学院学报,2014,29(3):21-26.
Chen C X,Wang R H,Wu J Y,et al.Effects of waterlogging stress on lipid peroxidation and antioxidant system in leaves ofToonacilliateseedlings [J].Journal of Northwest Forestry University,2014,29(3):21-26.(in Chinese)
[12] 陈建勋,王晓峰.植物生理学实验指导 [M].广州:华南理工大学出版社,2006.
Chen J X,Wang X F.Plant physiology experiment guidance [M].Guangzhou:South China University of Technology Press,2006.(in Chinese)
[13] 吴际友.红椿生理特性与家系选择研究 [D].长沙:中南林业科技大学,2012.
Wu J Y.Research on physiological characteristics and family selection ofToonaciliate[D].Changsha:Central South University of Forestry & Technology,2012.(in Chinese)
[14] 曹 慧,王孝威,韩振海,等.水分胁迫诱导平邑甜茶叶片衰老期间内肽酶与活性氧累积的关系 [J].中国农业科学,2004,37(2):274-279.
Cao H,Wang X W,Han Z H,et al.Relationship between changes of endopeptidases activity and active oxygen inMalushupehensisleaves during senescence induced by water stress [J].Scientia Agricultura Sinica,2004,37(2):274-279.(in Chinese)
[15] 周瑞莲,王 刚.水分胁迫下豌豆保护酶活力变化及脯氨酸积累在其抗旱中的作用 [J].草业学报,1997,6(4):39-43.
Zhou R L,Wang G.Role of protective enzymes changes and proline accumulation ofPisumsativumin drought resistance under water stress [J].Acta Prataculturae Sinica,1997,6(4):39-43.(in Chinese)
[16] 李少峰,李志辉,刘友全,等.水分胁迫对椿叶花椒抗氧化酶活性等指标的影响 [J].中南林业科技大学学报:自然科学版,2008,28(2):29-33.
Li S F,Li Z H,Liu Y Q,et al.Effect of water stress on activities of anti-oxidase and several physiological indexes ofZanthoxylumailanthoidesSieb.et Zucc [J].Journal of Central South University of Forestry & Technology:Nat Sci Ed,2008,28(2):29-33.(in Chinese)
[17] 欧 毅,王 进,吴天强,等.水分胁迫对桃形李叶片含水量、质膜透性和抗氧化酶活性的影响 [J].西南农业学报,2007,20(5):982-985.
Ou Y,Wang J,Wu T Q,et al.Effects of water stress on water content,relative membrane permeability and the activity of antioxidant enzymes of plum leaves [J].Southwest China Journal of Agricultural Sciences,2007,20(5):982-985.(in Chinese)
[18] 刘锦春,钟章成,何跃军.干旱胁迫及复水对卡斯特地区柏木幼苗活性氧清除系统的影响 [J].应用生态学报,2011,22(11):2836-2840.
Liu J C,Zhong Z C,He Y J.Effects of drought stress and re-watering on the active oxygen scavenging system ofCupressusfunebrisseedlings in Karst area [J].Chinese Journal of Applied Ecology,2011,22(11):2836-2840.(in Chinese)
[19] 山 仑,邓西平,张岁岐,等.生物节水研究现状及展望 [J].中国科学基金,2006(2):1-4.
Shan L,Deng X P,Zhang S Q,et al.Current research situation and prospects of biological water saving [J].Bulletin of National Natural Science Foundation of China,2006(2):1-4.(in Chinese)
[20] 刘吉利,赵长星,吴 娜,等.苗期干旱及复水对花生光合特性及水分利用效率的影响 [J].中国农业科学,2011,44(3):469-476.
Liu J L,Zhao C X,Wu N,et al.Effects of drought and rewatering at seedling stage on photosynthetic characteristics and water use efficiency of peanut [J].Scientia Agricultura Sinica,2011,44(3):469-476.(in Chinese)
[21] 闫永銮,郝卫平,梅旭荣,等.拔节期水分胁迫-复水对冬小麦干物质积累和水分利用效率的影响 [J].中国农业气象,2011,32(2):190-195.
Yan Y L,Hao W P,Mei X R,et al.Effects of water stress-rewatering at jointing stage on dry matter accumulation and WUE of winter wheat [J].Chinese Journal of Agrometeorology,2011,32(2):190-195.(in Chinese)
Canonical correlation analysis on leaf physiological responses ofToonacilliateRoem. seedlings to drought stress and rewatering
LIU Qiu1,2,LI Zhi-hui1,WU Ji-you2,CHEN Cai-xia1,LI Yan1, WU Zhi-hua3,CHENG Yong2,HUANG Ming-jun1
(1CollegeofForestry,CentralSouthUniversityofForestry&Technology,Changsha,Hunan410004,China;2HunanForestryAcademy,Changsha,Hunan410004,China;3ChinaEucalyptResearchCentre,Zhanjiang,Guangdong524022,China)
【Objective】 The study discovered the correlation of leaf physiological characteristics ofToonacilliateRoem.potted seedlings among drought stress and rewatering to build knowledge foundation for investigating drought tolerance ofToonacilliateRoem.【Method】 The drought stress and rewatering experiment was conducted in artificial-rain-shelter with 2 years oldToonacilliateRoem.potted seedlings.Eight leaf physiological indicators including leaf relative water content,leaf relative conductivity,chlorophyll content,SOD activity,POD activity,MDA content,free proline content and soluble protein content were determined.Canonical correlation was adopted to analyze the overall correlation of leaf physiological characteristics between control,drought stress and rewatering cases.【Result】 (1) There was significant correlation in leaf physiological characteristics ofToonacilliateRoem.potted seedlings between control and drought stress cases.The first coefficient of canonical correlation analysis was 0.996 5,Leaf relative conductivity and chlorophyll content mainly reflected the leaf physiological characteristics before and after drought stress,respectively.(2) There was significant correlation in leaf physiological characteristics ofToonacilliateRoem. potted seedlings between drought stress and rewatering cases. The first coefficient of canonical correlation analysis was 0.998 3.Leaf relative conductivity mainly reflected leaf physiological characteristics both in drought stress and rewatering cases.(3) There was significant positive correlation between chlorophyll content under drought stress and POD activity and MDA content in rewatering case,indicating that higher photosynthetic capacity under drought stress caused higher antioxidant enzyme activity in rewatering.(4) Soluble protein content under drought stress extremely correlated with POD activity in rewatering with the correlation coefficient of 0.803,indicating that the damage severity under drought stress closely influenced the reparation of leaf physiological characteristics in rewatering.(5) Water consumption and drought tolerance ofToonacilliateRoem.after rewatering were directly related to drought-resistant potential in normal watering case.【Conclusion】 Slight drought damage could positively promote the recovery of chlorophyll content and POD activity ofToonacilliateRoem.seedlings after rewatering.Thus,proper water deficit could improve the drought resistance ability of plants,while excessive water shortage could harm the plants and the reparation would last for a long period.
drought stress;ToonacilliateRoem.;rewatering;physiology response;biological water saving;canonical correlation analysis
时间:2015-09-09 15:41
10.13207/j.cnki.jnwafu.2015.10.006
2014-09-29
湖南省林业科学院青年科研创新基金项目(2013-13)
刘 球(1985-),女,湖南安化人,助理研究员,在读博士,主要从事森林培育和林木抗逆机理研究。
S792.990.1
A
1671-9387(2015)10-0035-10
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20150909.1541.012.html
猜你喜欢
小学生作文(低年级适用)(2022年10期)2022-10-31
现代临床医学(2021年1期)2021-01-26
食品安全导刊(2020年33期)2020-12-26
湖南饲料(2019年4期)2019-10-17
中国生物医学工程学报(2019年5期)2019-07-16
Coco薇(2017年5期)2017-06-05
山东农业科学(2017年4期)2017-05-18
中国火炬(2015年1期)2015-07-25
现代电生理学杂志(2015年4期)2015-07-18
江苏农业科学(2014年10期)2014-11-22
