
小陇山百花林场次生松栎混交林群落及优势种群空间结构特征1)
2015-03-08 05:45张岗岗王得祥张丛珊张宋智刘文桢欧阳君祥
东北林业大学学报 2015年8期
张岗岗 王得祥 张丛珊 张宋智 刘文桢 欧阳君祥
(西北农林科技大学,杨凌,712100) (甘肃省小陇山林业科学研究所) (国家林业局调查规划设计院)
责任编辑:潘 华。
森林结构是森林动态变化过程中测度时点的林分状态的高度概括和度量,是对林分发展过程如更新方式、竞争、自然稀疏和经历的干扰活动的综合反映[1],也是影响森林生态系统多样性、决定森林生态系统稳定性和功能效率的主要因素[2],已成为分析和管理森林生态系统的重要因子。树木在林地上的分布格局及其属性在空间上的排列方式构成了林分空间结构,决定了树木之间的竞争势及其空间生态位,且在很大程度上决定了林分的稳定性、发展的可能性和经营空间大小[3-4],直接关系到森林生态系统功能的发挥和生物多样性的保存[1],而且林分的空间结构越优,林分的功能越强,稳定性也越高。因此,对林分空间结构特征进行准确描述、合理解释和应用研究方兴未艾。种群是群落结构和功能的基本单元,群落的优势种对其结构的形成起着主导作用,要理解群落如何建成,需了解种群(特别是优势种种群)结构如何影响群落结构,才能客观体现群落的发展、演变趋势[5],但目前对优势种群和群落空间结构特征关系的研究却鲜见报道。林木大小、格局和树种是森林经营中可操作的主要结构因素[6],直接与种间、种内竞争,幼苗更新、生长以及植物对环境的综合利用紧密相关[7],因此,基于相邻木之间空间关系的结构调整可以作为次生林经营的有效手段。一元分布或均值可以独立地描述林分单方面结构特征或平均生长状态[4,8-9],而二元分布从两个方面刻画林分的结构特征,克服了一元分布在表达结构方面的缺陷,提供更加丰富、直观的有效信息[7,10-12]。由此可见,借助一元分布和二元分布探讨群落和主要种群结构关系并指导次生林经营,理论上正确、操作上可行、方法上合理,是深入地认识相邻木之间空间关系,实施次生林可持续经营战略的必然选择。
甘肃小陇山林区是我国主要的次生林分布区,林区的天然林主要是次生林,因受气候、地形等环境影响,森林组成结构复杂,其中以华山松、油松与锐齿栎、辽东栎、红桦等形成的针阔混交林占次生林面积的75%[13]。次生林作为该区森林资源的主体,对其种群结构、多样性、更新及其合理经营一直是研究的焦点[8,13-14],而对该区域次生松栎混交林群落及优势种群空间结构特征的研究却鲜见报道。本研究采用一元分布、二元分布从不同角度量化分析小陇山次生松栎混交林林群落及其主要种群空间结构一元分布、二元分布特征,以期为小陇山林区次生林精细的结构分析和可持续经营提供借鉴。
1 研究区概况
研究区域位于秦岭西段小陇山林区百花林场,北纬34°16'~34°25',东经106°15'~106°30',境内地势由西北向东南倾斜,山体相对高差在100~550 m,坡度为26°~45°,海拔为1 442.0~2 489.2 m,年平均气温10.9 ℃,年均降水量800 mm,年平均蒸发量1 420 mm,湿润度0.36~0.45,相对湿度69%,无霜期184.8 d,年均日照时间2 098.7 h,土壤以山地棕壤和山地褐土为主,pH 值5.0~7.0 地貌类型为土石侵蚀剥蚀中山山地,属暖温湿润一中温带半湿润大陆性季风气候类型。根据全林调查资料和往年历史资料,百花林场大干子沟2 号小班于1976—1979年经过择伐作业后形成次生松栎针阔混交林,林分中除主要建群种油松(Pinus tabulaeformis)、华山松(Pinus armandii)、锐齿栎(Quercus aliena var.acuteserrata)外,主要伴生树种有华榛(Corylus chinensis)、兴山榆(Ulmus bergmanniana)、椴树(Tilia tuan)、漆(Toxicodendron vernicifluum)、山核桃(Carya cathayensis)、水榆花楸(Sorbus alnifolia)等共计44 个树种,具有典型的次生林特征和区域代表性。
研究样地位于百花林场英崖管护区大干子沟,样地面积50 m×50 m,海拔1 376 m,坡向东,坡度36°,中下坡位,样地内林木总计767 株,总蓄积39.981 5 m3,总断面积7.230 7 m2,平均胸径11.6 cm,平均树高10.9 m。树种组成为4 锐齿栎4 油松1 华山松1 阔杂,其中锐齿栎种群林木259 株,占总株数量的33.8%,平均胸径11.1 cm,平均树高10.2 m;油松种群林木212 株,占总株数量的27.6%,平均胸径11.6 cm,平均树高11.3 m。
2 研究方法
2.1 野外调查及样地设置
于2012年7—9月,在百花林场选取典型地段设置50 m×50 m 固定样地进行全林调查,利用TOPCON 全站仪对样地内胸径≥5 cm 的林木进行定位,测定并记录每株树木的X、Y 坐标、树种、胸径、树高、枝下高、冠幅等,记录样地的海拔、经纬度、坡向、坡度、坡位和土壤状况等立地条件因子。为避免边缘效应的影响,在样地边缘设置5 m 缓冲区。
2.2 数据处理
基于相邻木空间关系的林分空间结构指标能够准确地描述林分空间结构特征,各指标混交度(Mi)、大小比数(Ui)及其角尺度(Wi)的计算方法和取值意义参照文献[15]。利用Winkelmass1.0 计算每株林木的角尺度、大小比数、混交度,再用Excel透视表分别统计群落及其油松、锐齿栎种群一元分布和二元分布频率,利用Sigmaplot12.5 绘制X-Y 二元分布图。
3 结果与分析
3.1 松栎林群落及优势种群一元分布特征
松栎林群落中55.8%的林木处于随机分布,均匀和不均匀的林木分别占22.0%、15.7%,处于很均匀和很不均匀的林木极少,仅为0.9%和5.6%,角尺度为0.508,落在[0.475,0.517]范围内,说明林分整体处于随机分布;不同优劣程度林木比例均接近20%,频率值介于17.7%~22.0%,大小比数平均值(ˉU)为0.503,说明林木分化比较严重,整体生长处于中庸状态;处于强度和极强度混交的林木比例较大,总计达64.0%,其次是中度混交林木(20.5%),零度混交和弱度混交林木较少,分别占总株数的3.7%、11.9%,混交度平均值(ˉM)为0.696,说明群落混交良好,周围相邻木大多为其他树种。油松种群中随机分布的林木占59.7%,均匀分布林木(20.9%)略大于不均匀分布林木(14.7%),其余分布格局林木极少,角尺度为0.508,油松种群林木分布格局属随机分布;优势和亚优势林木总计达49.6%,明显多于劣势和绝对劣势林木比例(29.4%),中庸状态林木占20.9%,大小比数平均值(ˉU)为0.419,说明该种群在群落中占有一定的竞争优势;中度混交林木最多,达31.8%,强度混交和极强度混交林木分别占27.9%、15.5%,明显多于比零度混交(5.4%)和弱度混交(19.4%)林木,混交度平均值(ˉM)为0.572,说明油松种群混交良好,大多数林木个体与其他树种相伴而生。锐齿栎种群中,中庸状态林木仅为53.3%,左右两侧林木分布比例分别为23.4%、23.3%,整体近似呈正态分布,角尺度为0.512,属随机分布;25.3%林木处于优势状态,其余优劣程度的林木比例接近,介于18.0%~19.3%,大小比数平均值(ˉU)为0.463,处于中庸状态;强度混交林木高达38.7%,中度混交次之,占26.0%,再次是弱度混交林木(15.3%),极强度混交和零度混交林木较少,混交度平均值(ˉM)为0.592,说明锐齿栎种群空间隔离程度较高。
3.2 松栎林群落二元分布特征
由图1可知,松栎混交林群落在同一大小比数等级上,频率最大值均处于随机分布轴,介于9.9%~12.7%,总计达55.8%,并向两侧逐渐减小,说明同一优劣程度林木个体大多数随机分布;同一分布格局的林木各大小比数等级上的频率分布差异不明显,即相同分布格局林木个体处于不同优劣等级的近似相等。随着角尺度的增大,同一混交等级对应频率值先增大后减小,且在Wi=0.5 时取得最大值,即相同混交等级的林木大多处于随机分布;同一分布格局(除Wi=0)对应频率值随着混交度取值的增大而逐渐增大,并在Wi=0.50 且Mi=0.75、1.00时取得较大值,分别为16.2%、18.8%,即林分中较为常见的结构单元参照树周围相邻木大多为其它树种且呈随机分布。群落中同一优劣程度对应的频率值随着混交度等级的增大逐渐增大,且混交良好(Mi=0.75、1.00)的林木株数高达64%,而混交较差的林木较少;同一混交等级上,频率值随着大小比数等级的增大差异不明显,即相同混交等级林木处于不同优劣等级的株数比例大致相等。

图1 松栎混交林群落不同林分空间结构下的二元分布
3.3 油松种群二元分布特征
由图2可知,油松种群中同一优劣程度对应的频率值则随着角尺度等级的增大呈现先增大后较小的趋势,且在Wi=0.5 时取得最大值,整体近似呈正态分布,即相同优劣等级的林木大多处于随机分布,处于其它分布格局的林木较少;随机分布(Wi=0.50)的林木随着大小比数的增大呈下降趋势,但不明显,其余分布格局的林木则基本无变化,即同一分布格局的林木在不同优劣等级分布比例相近。油松种群中同一混交等级上,频率值随着角尺度等级的增大先增大后减小,说明同一混交等级的林木大多处于随机分布,其它分布格局的林木较少;Wi=0.75、0.50、0.25 的林木随着混交度等级的增大逐渐增大后略有下降,Wi=0或1.00 的林木总计不足5%,其中Wi=0.5 且Mi=0.5、0.75 的林木总计达35.7%,即呈随机分布且处于中度混交或强度混交的林木构成的结构单元在林分中较常见。油松种群中同一优劣程度的林木在中度混交轴(Mi=0.50)上取得最大值,且向两侧逐渐减少,其中,中度混交且中庸状态的林木高达9.3%,即同一优劣程度的林木大多处于中度混交,其他混交状况的较少;Mi=0、0.25 的林木随着大小比数等级的增大无明显变化,其余混交等级的林木则趋于平缓减少,即油松种群中处于优势(Wi=0、0.25)且混交良好(Mi=0.75、1.00)的林木占有较大比例,为30.2%。
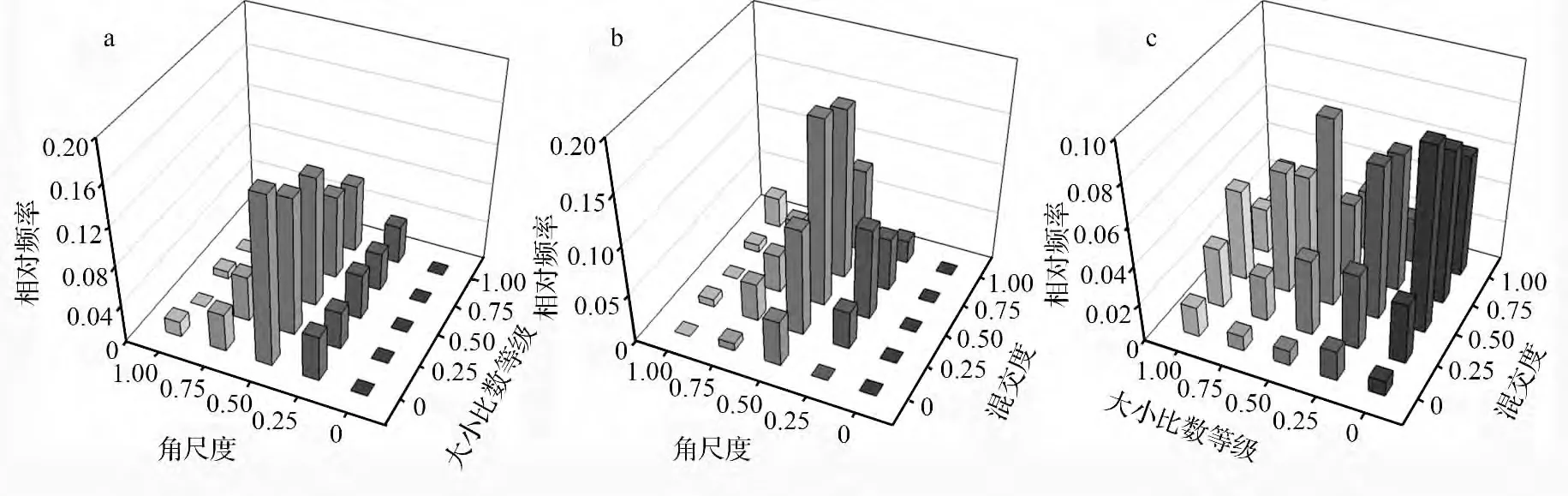
图2 油松种群不同林分空间结构下的二元分布
3.4 锐齿栎种群二元分布特征
由图3可知,锐齿栎种群中同一优劣程度的林木随着角尺度等级的增大先增大后较小,且在角尺度为0.5 时取得最大值,频率值介于7.3%~14.0%,整体近似呈正态分布,即同一优劣等级的林木大多处于随机分布,处于其它分布格局的林木较少;Wi=0、0.75、1.00 的林木在不同优劣等级分布比例相近,随机分布(Wi=0.50)的林木随着大小比数的增大逐渐减少,均匀分布(Wi=0.25)的林木则先下降后上升,但整体变化趋势不明显。锐齿栎种群中同一混交等级上,频率值随着角尺度等级的增大先增大后减小,近似呈正态分布,说明同一混交等级的林木大多处于随机分布,其它分布格局的林木较少;同一分布格局(除Wi=1.00)对应的频率值先增大后减小,其中Wi=0.5 且Mi=0.75 的林木达19.3%,即林分中常见结构单元参照树周围相邻木处于强度混交且呈随机分布。锐齿栎种群中同一优劣程度的林木在强度混交轴(Mi=0.75)上取得最大值,且向两侧逐渐减少,即相同优劣程度的林木大多处于强度混交;同一混交等级的林木随着大小比数等级的增大无明显变化,说明同一优劣等级的林木处于各优劣等级的林木比例相等。

图3 锐齿栎种群不同林分空间结构下的二元分布
4 结论与讨论
小陇山油松、锐齿栎种群结构参数一元分布规律与整个群落的大致相同,即角尺度频率分布均呈正态分布,角尺度分别为0.508、0.508、0.512,说明油松、锐齿栎种群和群落中多数林木和各自总体均处于随机分布,符合顶级群落及其优势种群分布格局特征;大小比数分化不明显且频率分布较均匀,ˉU 分别为0.419、0.463、0.503,说明不同生长状态的林木株数比例相差不大,林木分化程度较高,整体处于中庸状态,油松和锐齿栎种群中优势和亚优势林木比例略高,原因在于油松和锐齿栎优势程度较大,具有明显的竞争势[16],能够占据主林层有利的生态位,林木生长受周围相邻木干扰较小,其它树种基本处于林冠中下层,对资源和空间的竞争势较弱,趋于劣势,致使林分总体大小比数分布较为均匀;油松和锐齿栎种群混交度分布先增大后减小,中度混交和强度混交的林木较多,混交度平均值(ˉM)分别为0.572、0.592,树种空间隔离程度较高,群落混交度分布则呈递增趋势,混交良好(Mi=1.00、0.75、0.50)的林木较多,混交度平均值(ˉM)为0.696,明显高于油松和锐齿栎种群,这主要是因为林分中有华山松、千金榆等伴生树种共计42 种,物种丰富但株数和断面积比例均比较低[16],往往散生于群落中,大多与其它树种相伴而生,促使群落中极强度和强度混交林木比例高达64.0%,而该区域择伐作业后进行了以油松为主的团块状造林[16],锐齿栎则具备较强的萌蘖能力,这在一定程度上影响着种群的混交程度,但经过长期的自然演替,整体混交良好。油松、锐齿栎作为次生林群落建群种和优势种,株数和断面积比例占绝对优势[16],与华山松、千金榆、鹅耳枥等伴生种经过不断地竞争、适应和选择,群落中林木间关系复杂多样、高度协调发展,混交度逐步提升[17],水平分布格局趋于随机分布[18],形成了典型的天然次生针阔混交林,说明油松、锐齿栎种群对群落结构和环境的形成有明显的控制作用,在很大程度上决定了群落的结构特征和发展趋势,也侧面反映出不同层次物种对群落的贡献是不等价的,尚需进一步探讨其它伴生树种的结构特征及其环境因素的影响,才能更好地反映群落总体结构的数量特征。
小陇山次生松栎混交林群落及油松和锐齿栎种群角尺度—大小比数、角尺度—混交度、大小比数—混交度二元分布特征基本一致,即同一优劣程度或混交等级的林木大多处于随机分布,同一分布格局或混交等级的林木处于不同优劣程度的林木比例近似相等;林分中较为常见的结构单元参照树周围相邻木大多为随机分布的其它树种;但值得注意的是在群落中处于相同优劣等级的林木大多混交良好,油松种群中相同优劣程度的林木频率大多处于中度混交,锐齿栎种群中相同优劣程度的林木则大多处于强度混交,这可能是因为该区域择伐作业后在该区域中、下部以及局部进行了以油松为主的林冠下和团块状造林[16],局部形成大小不等的团状、块状纯林呈镶嵌状分布,其次,锐齿栎的竞争能力比油松强[19],林木分化突出,导致自疏和他疏,出现的林隙有利于其他伴生种的侵入和生长,随着群落不同个体的生长发育和激烈的种间竞争,使得同种个体趋于分散,混交度逐步提高,致使锐齿栎种群中相同优劣程度的林木空间隔离程度较高。除了鄂椴、千金榆、三桠乌药,这几个树种有“零度混交”状态出现,其它伴生树种的混交度分布频率主要集中在“强度混交”与“极强度混交”的状态[16],使得群落总体中相同优劣程度的林木混交良好。张连金等[1]发现林分中极强度混交的林木大都处于劣势,本文研究的次生林群落中混交良好的林木处于劣势(Mi、Ui=0.75、1.00)的占25.9%,明显多于优势木,进一步统计发现群落中仅仅有锐齿栎、油松、华山松、漆、小叶钓樟(Lindera aggregata)的相对多度≥0.5%(株数量≥38 株),其余树种个体数量极少,大多与其他树种伴生,且处于林分中下层,使得这些树种在群落中混交良好但处于劣势地位,而这种结构特性的林木在优势种群油松、锐齿栎中分别仅占10.8%、18.0%,明显多于劣势木,但在局部环境中并不一定处于优势。因此,区分树种(尤其是优势种群)分析林木分布状况和数量特征是对群落结构特征的进一步细化和深入探讨,有助于阐明群落结构建成和稳定性影响因素,同时,这些独特的结构特征对经营次生林具有指导意义,若该林分以生态防护为目标,则应适当选取混交良好林木中“霸王木”予以伐除,以释放林冠中下层被压木,提高物种多样性和空间隔离程度,否则应伐除这些劣势干扰木,以培育中大径材林木。
小陇山林区是我国西北地区重要的天然林区,锐齿栎天然次生林为其主要的林分类型,合理地经营次生林是实现当地林业可持续发展的关键[14]。就目前的认识水平来看,完全可以将未经人为干扰的天然林或经过微弱干扰已得到恢复的天然林结构特征或原始林、顶级群落的共同特征作为同地段现有森林的经营方向[15],而通常认为松类树种终将为栎类植物所取代[19-20],因此,小陇山林区锐齿栎天然林群落作为当地森林演替的顶级群落,可作为锐齿栎次生林经营的模式林分[9]。小陇山林区王安沟100 多a 未经人为干扰的锐齿栎天然林林木分布格局呈随机分布,各树种呈高度混交,锐齿栎占优势地位[14],符合森林演替顶级群落结构特征[17-18]。本研究结果与锐齿栎天然林结构特征较为一致,说明该次生林林分结构目前处于较为合理的状态,应加速其顺向演替的进程,使其尽快处于稳定的顶级群落阶段。而小陇山响潭沟锐齿栎次生林总体结构特征表现为进展演替群落,但林木分布格局呈轻微团状分布,混交较差,建群种锐齿栎处于亚优势的状态,白营西沟次生林林木的分布格局为随机分布,但树种混交程度较低,建群种锐齿栎在林分中处于亚优势状态[14],虽然锐齿栎原始林各树种呈高度混交,锐齿栎在林分中占绝对优势,但其林木分布格局呈轻微团状分布[9],这些林分结构特征仍与锐齿栎天然林有较大差距,迫切需要以同地段顶级群落锐齿栎群落特征为模式,充分利用结构参数一元分布和二元分布在表达结构上的频率优势[7],从单方面或两方面量化分析不同水平层次林木的分布状况和数量特征的基础上,采取恰当的经营措施有针对性地调整次生林分的水平分布格局、树种组成和各树种的竞争关系,从而实现小陇山林区次生林精细的结构调整,加速其顺向演替进程,使次生林分尽早恢复到健康稳定的顶级天然林群落阶段。
[1] 张连金,胡艳波,赵中华,等.北京九龙山侧柏人工林空间结构多样性[J].生态学杂志,2015,34(1):60-69.
[2] Polyakov M,Majumdar I,Teeter L.Spatial and temporal analysis of the anthropogenic effects on local diversity of forest trees[J].Forest Ecology and Management,2008,255(5):1379-1387.
[3] Pretzsch H.Analysis and modeling of spatial stand structures.Methodological considerations based on mixed beech-larch stands in Lower Saxony[J].Forest Ecology and Management,1997,97(3):237-253.
[4] 安慧君.阔叶红松林空间结构研究[D].北京:北京林业大学,2003.
[5] 何建敏,王震洪.雷公山灌木林群落结构及其优势种种群特征关系研究[J].广东农业科学,2014,41(6):178-182.
[6] Kint V.Structural development in ageing temperate Scots Pine stands[J].Forest Ecology and Management,2005,214(1/3):237-250.
[7] 李远发.林分空间结构参数二元分布的研究[D].北京:中国林业科学研究院,2013.
[8] 袁士云,张宋智,刘文桢,等.小陇山辽东栎次生林的结构特征和物种多样性[J].林业科学,2010,46(5):27-34.
[9] 郭小龙,刘文桢,张宋智,等.小陇山林区锐齿栎原始林群落的空间结构特征[J].西北农林科技大学学报:自然科学版,2014,42(11):106-112,120.
[10] Wang M L,Rennolls K,Tang S Z.Bivariate distribution modeling of tree diameters and heights:Dependency modeling using copulas[J].Forest Science,2008,54(3):284-293.
[11] Li Y F,Hui G Y,Zhao Z H,et al.The bivariate distribution characteristics of spatial structure in natural Korean pine broadleaved forest[J].Journal of Vegetation Science,2012,23(6):1180-1190.
[12] Li Y F,Hui G Y,Zhao Z H,et al.Spatial structural characteristics of three hardwood species in Korean pine broad-leaved forest-Validating the bivariate distribution of structural parameters from the point of tree population[J].Forest Ecology and Management,2014,314:17-25.
[13] 王晓春,王金叶,江泽平.甘肃小陇山次生林经营技术研究[J].西北林学院学报,2008,23(3):142-146.
[14] 赵中华,白登忠,惠刚盈,等.小陇山不同经营措施下次生锐齿栎天然林物种多样性研究[J].林业科学研究,2013,26(3):326-331.
[15] 惠刚盈,赵中华,胡艳波.结构化森林经营技术指南[M].北京:中国林业出版社,2010.
[16] 袁一超.结构化经营方法和技术在甘肃小陇山典型针阔混交林中的应用研究[D].杨凌:西北农林科技大学,2014.
[17] 胡艳波,惠刚盈.优化林分空间结构的森林经营方法探讨[J].林业科学研究,2006,19(1):1-8.
[18] 张家城,陈力,郭泉水,等.演替顶级阶段森林群落优势树种分布的变动趋势研究[J].植物生态学报,1999,23(3):256-268.
[19] 徐化成.油松[M].北京:中国林业出版社,1990.
[20] Barbéro M,Loisel R,Quézel P,et al.Pines of the Mediterranean Basin[M]//Richardson D M.Ecology and Biogeography of Pinus.Cambridge:Cambridge University Press,1998.
猜你喜欢
现代园艺(2018年2期)2018-03-15
现代农业研究(2017年11期)2018-01-12
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
现代园艺(2017年20期)2017-02-03
甘肃林业(2016年4期)2016-11-07
甘肃林业(2016年4期)2016-11-07
甘肃林业(2016年6期)2016-02-27
