
2套半同胞籼粳交重组自交系粒形性状的相关和遗传分析
2015-03-24 02:27叶乃忠肖应辉
作物研究 2015年2期
叶乃忠,曾 盖,唐 伟,肖应辉
(湖南农业大学农学院,长沙 410128)
(续表3)
2套半同胞籼粳交重组自交系粒形性状的相关和遗传分析
叶乃忠,曾 盖,唐 伟,肖应辉*
(湖南农业大学农学院,长沙 410128)
利用以粳稻品种日本晴为背景与不同粒形供体亲本构建的2套半同胞籼粳交重组自交系(RILs)为材料,分析了连续2年田间种植的水稻籽粒粒长、粒宽和粒厚等粒形性状的表型分布特征。结果显示,所有粒形性状均表现为单峰或多峰连续分布特征。采用主基因+多基因混合遗传模型,分析了各性状的遗传方式,结果表明:在日本晴/特大籼RILs中,粒长由2对显性上位作用的独立主基因+多基因控制,粒宽由2对重叠作用的独立主基因控制,粒厚由2对等加性作用的连锁主基因控制;在日本晴/R1126 RILs中,粒长表现为2对互补或抑制作用的独立主基因+多基因遗传模式,粒宽遗传由2对互补作用的独立主基因控制,粒厚由4对主基因+多基因控制。
水稻;半同胞;重组自交系;粒形性状;遗传模型
籽粒的粒长、粒宽和粒厚等粒形性状是水稻重要的经济性状,与产量和品质均有密切关系。一方面,水稻的这些粒形性状直接决定其粒重,进而影响水稻产量[1];另一方面,水稻粒形也是决定稻米外观品质和商用品质的重要因素[2],粒长、粒宽及长宽比是稻米外观品质性状的重要指标[3],粒长和长宽比更是影响整精米率的关键因子[4],所有这些都直接关系到稻米的商品和市场价值。为了降低杂交水稻种子生产成本,近年科学家们提出了混播制种的杂交制种技术[5~7],选育粒形性状差异显著、适合机械分离加工的不育系和恢复系是该杂交制种技术得以应用的技术关键。因此,加强水稻籽粒粒形的遗传机理研究,对于通过遗传调控培育符合粒形目标要求的水稻品种具有重要意义。
关于水稻粒形性状间的相互关系,国内外开展了大量研究,因研究材料不同,结论存在较大差异。一般认为,水稻籽粒粒长与粒宽、粒厚呈负相关;粒宽与粒厚呈正相关[8]。鉴于水稻籽粒大小和粒重在产量形成中的重要作用,其遗传机制和调控机理方面的研究一直备受关注。多数研究观察到不同自然分离世代粒形性状表现为连续正态分布特征,并存在超亲分布现象,推断水稻粒形性状属于多基因控制的数量性状[9~11]。近20年来,科学家们利用不同类型的遗传群体,累计报道了400多个与水稻籽粒大小和形态有关的QTL[12],但迄今仅有GW2[13]、GS3[14]、GW5[15]、GS5[16]、GW8[17]、qGL3(qGL3.1)[18]、GIF1[19]、TGW6[20]等8个与籽粒大小有关的QTL被分离克隆。其中,GS3是第一个被克隆的粒形基因,主要影响粒长、长宽比和粒重;qGL3(qGL3.1)对粒长、粒宽、粒厚、粒重和灌浆速率均有影响;GW8与GS5功能相似,通过促进细胞分化和加快籽粒灌浆来增大粒宽和粒重;GW5和GW2则通过增大胚乳细胞使粒宽和粒重增大;GIF1和TGW6主要通过影响籽粒灌浆速率来调控粒重,对粒宽和粒厚等性状不起作用。
不断发掘新的粒形相关基因/QTL,通过分子技术进行精细定位和分离克隆,阐明粒形性状的遗传调控机制,是通过分子设计塑造不同粒形水稻品种的前提。本研究利用小粒粳稻品种日本晴为背景亲本,分别以大粒籼稻品系特大籼(TDX)和中粒形籼稻品系R1126为供体亲本构建的2套半同胞籼粳交重组自交系为材料,旨在探索粒形性状在不同背景中的遗传规律,为进一步发掘新的粒形相关基因并精细定位提供依据。
1 材料与方法
1.1 试验材料
亲本品种(系)包括小粒形粳稻品种日本晴(NPB)、大粒形籼稻品种特大籼(TDX)、中粒形籼稻品系R1126。重组自交系群体(RILs)2套,其中日本晴/特大籼(NPB/TDX)RILs(F11)含240个株系,日本晴/R1126(NPB/R1126)RILs(F12)含217个株系,2套群体均是在杂种F1自交分离获得F2群体的基础上通过单粒传法构建而成。
1.2 种植方法
田间试验分别于2012年和2013年在湖南农业大学水稻试验基地进行。2年均于当年5月25日播种所有材料,秧龄25 d左右移栽。每个株系种植1行,每行10株,株行距为20 cm×20 cm。田间管理和病虫防治按当地水稻常规栽培技术执行,期间未发生鸟鼠和病虫为害。
1.3 粒形性状调查方法
于成熟期拔除各株系中的异株和杂株,在每株系中随机选取非边行的5株主穗混合收种,籽粒晒干后用于后续分析。每个株系随机选取10粒饱满种子,用电子游标卡尺(精确到0.01 mm)分别测定籽粒粒长、粒宽和粒厚,取10粒平均值作为该株系相应性状的表型值。
1.4 数据分析软件

2 结果与分析
2.1 亲本及重组自交系群体的粒形性状分布特征
3个亲本品种(系)中,粒长、粒宽和粒厚3个性状均呈现出特大籼>R1126>日本晴的趋势(表1),尤其是品种间的粒长差异明显,特大籼和R1126的粒长分别约为日本晴的2倍和1.5倍。同一品种的同一粒形性状在不同年份略有差异,所有性状变化趋势一致,2013年各性状表型值小于2012年的对应值。与粒长相比,粒宽和粒厚性状的年际间变异相对较大。

表1 亲本在不同年份的粒形性状表现
在NPB/TDX和NPB/R1126重组自交系群体中,水稻粒形性状均呈连续分布,并且大多出现了超亲分布现象,暗示2群体中所有粒形性状均可能是多基因控制的数量性状(图1)。同一群体同一粒形性状在不同年份的分布特征基本相似,具有较好的重演性。同一粒形性状在不同群体中的分布特征各有差异,这种差异依具体性状而异。如NPB/TDX RILs中,粒长表现为双峰分布,而在NPB/R1126中则呈单峰分布。与之相反,粒宽性状在NPB/TDX RILs中呈单峰分布,而在NPB/R1126中则呈多峰分布。粒厚性状总体表现为单峰分布,但在不同群体中表现有一定差异。粒长、粒宽和粒厚性状在不同群体中的分布差异,暗示在不同遗传背景中控制这些性状的遗传机制有差异。

图1 2套半同胞重组自交系粒形性状的频率分布Fig. 1 Frequency distribution of the grain shape traits in the two sets of RILs
2.2 重组自交系群体的粒形性状间的相关分析
对2套半同胞重组自交系的粒长、粒宽和粒厚性状分年度进行了相关分析,2年数据具有较好的一致性(表2)。在NPB/TDX重组自交系群体2年的数据中,粒长、粒宽和粒厚间均呈正相关,粒长—粒宽间相关系数达0.05显著水平,粒长—粒厚、粒宽—粒厚均在0.01水平上呈显著正相关。对于NPB/R1126重组自交系群体,粒长—粒宽不存在显著的相关关系,而粒长—粒厚、粒宽-粒厚均在0.01水平上呈显著正相关。
粒长、粒宽和粒厚不同性状间的相关关系,在2套群体中表现大体一致。相对于粒长,粒宽和粒厚间的相关性更为密切,推断两者在遗传上可能存在一定的连锁关系,有待于进一步深入研究。
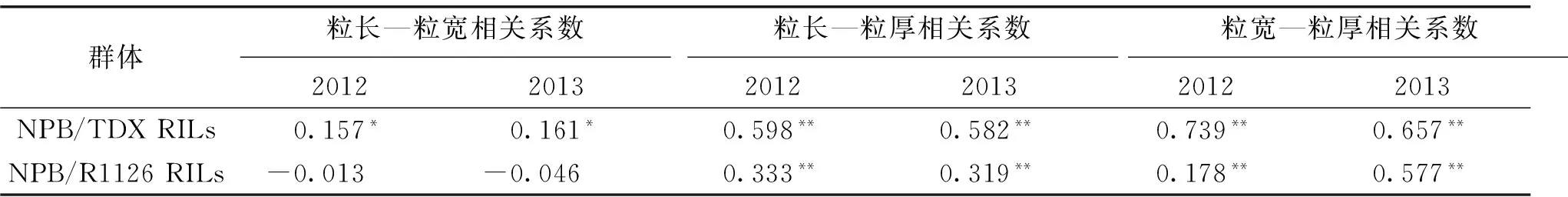
表2 不同粒形性状间的相关系数
注:*,** 分别表示在5%和1%水平上差异显著。
2.3 粒形性状遗传分析
2.3.1 遗传模型的选择及适合性分析
根据盖钧镒院士等提出的植物数量性状主基因+多基因混合遗传模型,分别计算不同年份不同群体不同粒形性状各遗传模型的AIC值(未列出所有数据)。对于每群体每年度的每一性状,选取AIC值最小的2个遗传模型作为备选模型,以用于进一步检验(表3)。在NPB/TDX重组自交系群体中,粒长性状在2012年的2个备选模型为E-1-4和E-1-2,2013年的为E-1-4和E-1-1;粒宽性状在2年中2个备选模型均为B-1-8和B-2-7;粒厚性状2012年备选模型为B-1-5和B-2-3,2013年为F-3和B-2-3。在NPB/R1126重组自交系群体中,粒长性状两年备选模型相同,均为E-1-7和E-1-9;粒宽性状在2012年备选模型为B-1-7和C-1,2013年为B-1-7和B-2-5;
粒厚性状在2012年备选模型为I-7和I-8,2013年为I-7和I-6。对所有性状不同备选模型的适合性检验表明,在NPB/TDX RILs中,粒长性状2年的最优遗传模型均是E-1-4,即该重组自交系群体粒长性状由2对显性上位作用的独立主基因+多基因控制;粒宽性状的最优模型是B-1-8,即表现为2对重叠作用独立主基因遗传模型;粒厚性状的最优模型为B-2-3,即该性状由2对等加性作用的连锁主基因控制。在NPB/R1126 RILs中,E-1-7与E-1-9统计量达到显著水平的个数相等,因此,E-1-7与E-1-9均可作为该重组自交系群体粒长的最优模型,即该重组自交系群体粒长性状表现为2对互补或抑制作用的独立主基因+多基因遗传模式;粒宽性状的最优模型为B-1-7,即2对互补作用独立主基因遗传模型;粒厚性状的最优模型是I-7,即4对主基因(2对等效)加性效应+多基因加性上位性控制。

表3 2套重组自交系群体粒形性状备选模型的适合性检验
(续表3)

群体性状年份模型U21U22U23nW2DnB-2-717.298917.31950.28964.68440.87222013B-1-80.00080.13891.89894.79890.8882B-2-70.00080.13891.89894.79890.8882粒厚2012B-2-30.00010.02500.35394.75400.9560B-1-50.21670.206113.09804.75490.95572013B-2-32.55422.75480.20184.84890.9530F-33.17791.23336.06154.85630.9521NPB/R1126RILs粒长2012E-1-71.94879.334346.43573.79610.8628E-1-91.94879.334346.43573.79610.86282013E-1-70.00002.956547.23993.77030.8242E-1-90.00002.956547.23993.77030.8242粒宽2012B-1-70.06790.00051.19764.36520.1402C-13.338711.3747423.01694.37650.37942013B-1-736.660534.66040.00984.35320.9754B-2-597.464872.379417.68474.34670.9820粒厚2012I-70.48130.195419.84994.39080.8894I-876.810765.90392.16384.40970.86562013I-70.09041.955645.66934.40150.8913I-64.149317.710980.00364.39810.8922
2.3.2 粒形性状遗传模型的参数估计
根据极大似然原理估算最优遗传模型的遗传参数(表4),NPB/TDX RILs的粒长主基因遗传率高达95%以上,多基因遗传率相对较小,且其加性效应为负效应;控制粒宽性状的主基因的遗传率在90%以上,其加性效应值表现为负值,即2对独立主基因的重叠作用使粒宽减小0.305~0.337 mm;控制粒厚性状的2对连锁主基因的遗传率相对较大,2年的贡献率分别为88.13%和91.18%。
在NPB/R1126 RILs中,控制粒长的主基因遗传率分别为88.71%和89.74%,2对主基因互补或重叠作用,使粒长减少0.592 mm和0.588 mm;控制粒宽的主基因遗传率分别为92.04%和93.39%,2对主基因的互补作用使粒宽减小0.19~0.21 mm; NPB/R1126 RILs粒厚的遗传主要表现为多基因遗传,主基因遗传效应只占44.88%和32.92%,其中遗传效应相对较大的2对等效基因使水稻粒厚减少0.071~0.086 mm。

表4 2套RILs群体粒形性状最优模型的遗传参数估计值
3 结论与讨论
前人利用各种不同的遗传群体,对水稻粒形性状的遗传规律开展了大量的研究,研究结论各有差异。本研究采用以粳稻品种日本晴为背景构建的2套半同胞籼粳交重组自交系,进行了粒形性状的遗传分析,结果发现同一粒形性状即使在具有同一背景亲本的不同群体中的分布特征也不相同,遗传模型分析进一步证实了在不同群体中控制这些性状的遗传机制是不同的。这一现象也暗示,本研究的2套重组自交系群体中的大粒亲本特大籼和中粒形亲本R1126控制粒形性状的基因可能具有较大差异。
本研究中,2套重组自交系群体的粒长、粒宽以及NPB/TDX RILs的粒厚均表现为2对主基因+多基因的遗传模型,其主基因的遗传效应均在88%以上,然而,控制不同性状的2对主效基因间作用方式不一样,如控制NPB/TDX RILs的粒长、粒宽和粒厚的主效基因间的互作方式分别为显性上位作用、重叠作用和等加性连锁,即使是控制同一性状的2对主效基因,在不同群体中的作用方式也不尽相同,如在NPB/TDX RILs和NPB/R1126 RILs中,控制粒宽的均为2对主效基因,但前者表现为重叠作用,后者表现为互补作用。值得注意的是,在NPB/R1126 RILs中,粒厚性状的遗传模式相对复杂,该性状的遗传以多基因遗传方式为主,4对主基因中即使是遗传效应相对较大的2对等效基因,也只能使水稻粒厚减少0.071~0.086 mm。
本研究结果将为进一步利用这2套群体进行粒形性状的基因定位提供有益信息,尤其是为后续的基因精细定位策略提供参考。如对于粒厚性状的基因定位,在遗传群体的选择上,NPB/TDX RILs优于NPB/R1126 RILs,利用前者进行基因定位有可能快速准确地将基因定位于目标染色体区段。又如,利用NPB/TDX RILs群体进行粒厚基因的精细定位时,回交构建近等基因系的过程中必须充分考虑到2对主效基因的连锁效应。
[1] 石春海,申宗坦.籼稻粒形及产量性状的加性相关和显性相关分析[J].作物学报,1996,22(1):36-42.
[2] 徐正进,陈温福,马殿荣,等.稻谷粒形与稻米主要品质性状的关系[J].作物学报,2004,30(9):894-900.
[3] 林建荣,石春海,吴明国.不同环境条件下粳型杂交稻稻米外观品质性状的遗传效应[J].中国水稻科学,2003,17(1):16-20.
[4] 徐富贤,熊 洪,朱永川,等.杂交中稻整精米率的影响因子及其间接鉴定方法研究[J].西南农业学报,2013,26(1):1-6.
[5] 刘延斌,杨远柱,刘建丰,等.杂交水稻亲本混播机械化制种研究进展[J].作物研究,2012,26(1):85-87,102.
[6] 汤国华,谢红军,余应弘.杂交水稻机械化制种研究的现状问题与对策[J].湖南农业科学,2012,(3):133-136.
[7] 王 杰,邓华凤,张武汉,等.杂交水稻机械化制种技术的研究现状与发展策略[J].安徽农业科学,2013,41(13):5682-5683,5686.
[8] 张颖慧,谢永楚,董少玲,等.利用水稻籼粳重组自交系群体研究粒型性状与千粒重的相关性[J].江苏农业学报,2012,28(2):231-235.
[9] 王松凤,李 辉,刘 喜,等.水稻粒形相关性状及千粒重QTL的稳定性分析[J].南京农业大学学报,2008,31(3):1-7.
[10] 张志勇.水稻粒型和粒重性状的主效QTL定位研究[D].厦门:厦门大学硕士学位论文,2009.
[11]于 波,高冠军,张庆路.水稻外观品质性状和千粒重的QTLs分析[J].湖北农业科学,2012,51(19):4187-4193.
[12]GRAMENE (2015).GRAMENE Database.[2015-01-31]http://www.gramene.org/.
[13]Song X,Huang W,Shi M,et al.A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J].Nat Gene,2007,39:623-630.
[14]Mao H,Sun S,Yao J,et al.Linking differential domain functions of theGS3 protein to natural variation of grain size in rice[J].Proc Natl Acad Sci USA,2010,107:19579-19584.
[15]Weng J,Gu S,Wan X,et al.Isolation and initial characterization ofGW5,a major QTL associated with rice grain width and weight[J].Cell Res,2008,18:1199-1209.
[16]Li Y,Fan C,Xing Y,et al.Natural variation inGS5 plays an important role in regulating grain size and yield in rice[J].Nat Genet,2011,43:1266-1269.
[17]Wang S,Wu K,Yuan Q,et al.Control of grain size,shape and quality byOsSPL16 in rice[J].Nat Genet,2012,44:950-54.
[18]Qi P,Lin Y,Song X,et al.The novel quantitative trait locusGL3.1 controls rice grain size and yield by regulating Cyclin-T1;3[J].Cell Res,2012,22:1666-1680.
[19]Wang E,Wang J,Zhu X,et al.Control of rice grain-filling and yield by a gene with a potential signature of domestication[J].Nat Genet,2008,40:1370-74.
[20]Ishimaru K,Hirotsu N,Madoka Y,et al.Loss of function of the IAA-glucose hydrolase geneTGW6 enhances rice grain weight and increases yield[J].Nat Genet,2013,45:707-711.
[21]何小红,盖钧镒.回交自交系群体数量性状遗传体系的分离分析方法[J].作物学报,2006,32(2):210-216.
[22]王金社,李海旺,赵团结,等.重组自交家系群体4对主基因加多基因混合遗传模型分离分析方法的建立[J].作物学报,2010,36(2):191-201.
Genetic and Correlation Analysis of the Grain Shape Traits with Two Set of Half-Sib Recombinant Inbred Lines Derived fromIndica/JaponicaRice Cross
YE Nai-zhong,ZENG Gai,TANG Wei,XIAO Ying-hui*
(College of Agronomy,Hunan Agricultural University,Changsha,Hunan 410128,China)
Two sets of half-Sib recombinant inbred lines (RILs),which with the japonica cultivar Nipponbare (NPB) as the common genetic background and crossed donor parents with different grain shape,were developed to research the genetic of grain shape traits.The frequency distribution of the grain shape traits including grain length (GL),grain width (GW) and grain thickness (GT) of rice seed harvested from the two sets of RILs were analyzed.The results showed that all the grain shape traits showed continuous distribution with single or multi peak.By using the mixed inheritance model with major genes plus polygenes,genetic analysis of the three grain shape traits of the sets of RILs were conducted.Genetic analysis showed that GL was controlled by two independent major genes with epistatic dominance plus polygenes,GW was controlled by two independent major genes with duplicate effect,GT was controlled by two linkage major genes with equal additive effect.In the NPB/R1126 RILs,GL was controlled by two major genes with complementary or inhibiting effect plus polygenes,GW was controlled by two independent major genes,whereas,GT was controlled by four major genes plus polygenes.The above results would provide useful information for the further gene mapping of grain shape traits with the two sets of RILs.
rice;half-Sib; recombinant inbred lines;grain shape trait;genetic model
2014-10-16
叶乃忠 (1989-),男,福建武夷山人,硕士研究生,Email:477583984@qq.com。
*通信作者:肖应辉,博士,教授,Email:xiaoyh@hunau.edu.cn。
教育部创新团队发展计划项目(IRT1239);湖南省高校科技创新团队支持计划项目。
S511.032
A
1001-5280(2015)02-0111-06
10.3969/j.issn.1001-5280.2015.02.01
猜你喜欢
区域治理(2022年40期)2022-11-27
四川蚕业(2022年2期)2022-11-19
作物研究(2020年5期)2020-12-08
世界科学技术-中医药现代化(2020年2期)2020-07-25
四川蚕业(2020年3期)2020-07-16
北京农学院学报(2020年1期)2020-03-09
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
北京农学院学报(2019年1期)2019-02-22
