
海雀稗对镉胁迫的生理响应及积累特性
2015-03-26 10:57吴朝波郭建春符少萍李瑞梅段瑞军
江苏农业学报 2015年6期
吴朝波, 郭建春, 符少萍, 刘 娇, 李瑞梅, 王 蕾,3, 段瑞军
(1.中国热带农业科学院/热带生物技术研究所,海南 海口 571101;2.海南大学农学院,海南 海口 571101;3.海南大学园林园艺学院,海南 海口 570228)
中国受重金属污染威胁的耕地面积约2×107hm2[1],2014年由环保部和国土资源部联合发布的《全国土壤污染状况调查公报》显示全国镉(Cd)污染超标率达7%[2]。镉毒性强,毒性持久,会对植物造成伤害。在镉胁迫下植物主要表现出叶茎黄化,叶片脱落,株高、叶长、叶宽、茎粗、根长等减少,过高的镉浓度使植物生长停滞,甚至出现枯死。在重金属胁迫下,植物体内会产生大量的活性氧(O·-2),使细胞膜脂过氧化加强,破坏膜的结构和功能[3]。抗氧化系统是植物体内镉伤害的主要解毒机制[4-5],由超氧化物歧化酶、过氧化物酶、过氧化氢酶等构成植物体内的主要活性氧清除系统[6-7],清除植物体内过多的活性氧(O·-2)。郑爱珍等[8]研究结果表明,白菜在镉处理下增加了植物叶片SOD、CAT的活性,有利于对活性氧的清除,因而增加了白菜对镉的耐性;彭艳[9]研究铝对小麦保护酶活性的影响,指出一定范围内保护酶活性随镉胁迫强度的增加而上升,SOD、POD、CAT活性的增强是维持小麦耐铝胁迫的重要生理基础;李丽峰等[10]研究镉胁迫对芦苇叶片SOD、POD活性影响的结果表明,当镉处理浓度<2 mg/kg时,SOD、POD活性随镉浓度增加而升高,能有效调节芦苇体内的活性氧平衡,此浓度内镉胁迫对芦苇的生长影响不大。海雀稗(Paspalum vaginatum Sw.)是一种可以用于草坪、饲料及盐碱地改良的多用途草种[11],有研究结果表明海雀稗可用于重金属(铅、锌)污染土壤的植物修复[12-15]。本研究以海雀稗为材料,研究其在镉胁迫下的生长、膜质损伤、抗氧化酶活性及其对镉的富集特征,旨在为应用海雀稗修复重金属镉污染土壤提供理论依据,为土壤镉污染植物修复技术的植物筛选提供参考依据[15]。
1 材料与方法
1.1 材料
海雀稗,采自海南岛文昌滨海滩涂。主要试剂: CdCl2·2.5H2O(分析纯)、α-萘胺、对氨基苯磺酸、亚硝酸钠、三氯乙酸(TCA)、硫代巴比妥酸(TBA)、甲硫氨酸(Met)、氮蓝四唑(NBT)、EDTA-Na2、核黄素等,均购自上海生工生物工程技术服务有限公司;主要仪器:电导率仪(DDS-11A型)、紫外可见光分光光度计(岛津UV-2501PC型);培养基质(土∶沙∶有机质=1∶1∶1,体积比)。
1.2 方法
1.2.1 试验材料处理 本研究于中国热带农业科学院热带生物技术研究所内进行,设置6个镉浓度,3次重复。将15 kg培养基质放入泡沫箱,分别用0 mol/L、1×10-5mol/L、1×10-4mol/L、5×10-4mol/L、1×10-3mol/L、2×10-3mol/L CdCl2·2.5H2O水溶液12 L进行处理,充分搅拌混匀,使每箱土壤中的镉浓度分别达到0 mg/kg、1 mg/kg、10 mg/kg、50 mg/kg、100 mg/kg、200 mg/kg,静置7 d,将海雀稗截取相同的茎段,每个茎段含2个节间,3个芽点,种植于处理好的土壤中,每箱12株,70 d后,测定其形态、生理指标。
1.2.2 形态指标测量 用直尺测量不同处理下的海雀稗株高(cm)、根长(cm);用精度为1/1000电子天平测定海雀稗根、茎、叶鲜质量(g),3次重复。
1.2.3 叶绿素含量测定 叶绿素含量测定使用紫外分光光度法。称取0.1 g鲜叶,剪成1 mm宽的细条,放入试管中,加10 ml提取液(乙醇∶丙酮=1∶1),密封,在黑暗条件下浸提24 h(至叶片脱色变白)。分别在663 nm、645 nm、440 nm下比色测定吸光值,3次重复。
1.2.4 根系活力测定 参照张宪政等[16]的方法,将待测根系吸干附着水后称取1 g,加入40 mg/L的α-萘胺溶液和磷酸缓冲液各25 ml,静置10 min后,吸取2 ml溶液,其余在25℃震荡3 h后再吸取2 ml溶液,在2次所吸取的测定液中各加入10 ml蒸馏水,1 ml 1%对氨基苯磺酸和1 ml 100 mg/L亚硝酸钠,静置显色5 min,再加入11 ml蒸馏水,混匀,测定510 nm波长下的吸光值,试验以不加根系的样品作为空白对照,3次重复。
1.2.5 丙二醛(MDA)含量测定 参照郑炳松[17]的方法测定MDA含量,将待测根、茎、叶吸干附着水后各称取0.5 g,加10ml 10%三氯乙酸(TCA)和少量石英砂研磨至匀浆,匀浆离心(2 000 g,15 min),取上清液2 ml,加0.6%硫代巴比妥酸(TBA) 2 ml,于沸水浴反应15 min,迅速冷却后离心(3 000 g,15 min)。取上清液测定532 nm、600 nm和450 nm波长的吸光度值,3次重复。
1.2.6 超氧化物歧化酶(SOD)活性测定 参照李和生等[18]、郝建军等[19]的方法测定SOD活性,将待测根、茎、叶的水分吸干后各称取0.5 g,液氮冰浴研磨,加入5 ml 0.05 mol/L磷酸缓冲液(pH 7.8),12 000 r/min冷冻离心20 min,获得酶粗提液,进行SOD活性测定,反应体系见表1,空白置于暗处,对照管与样品管置于4 000 lx荧光灯下显色反应20 min,以空白调零,测定569 nm波长的吸光值,各3个重复。
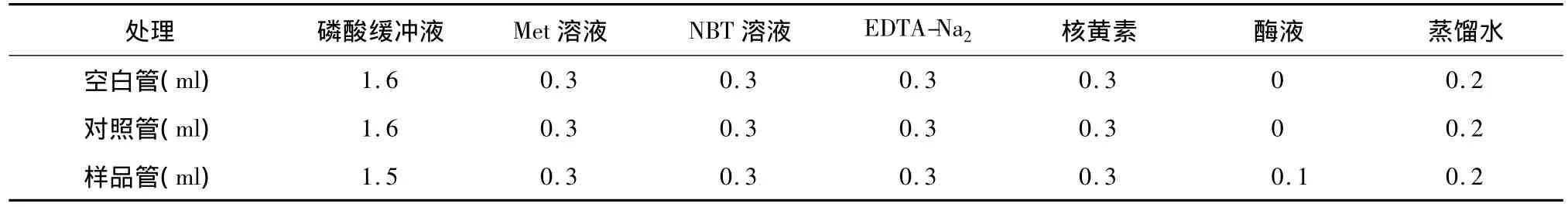
表1 SOD活性测定反应体系Table 1 The reaction system for SOD activity determination
1.2.7 过氧化氢酶(CAT)活性测定 采用彭志英等[20]的方法测定CAT活性,酶粗提液制备与SOD酶粗提液制备方法相同,取0.1 ml酶粗提液,加入2.5 ml反应液(0.1 mol/L的H2O2与0.1 mol/L pH 7.0的磷酸缓冲液按体积比为1∶4混合),于240 nm下比色,每隔1 min读数1次,共读3次,3次重复。
1.2.8 叶片细胞膜透性测定 参照刘宁等[21]、陈建勋等[22]的方法测定质膜相对透性,称取0.1 g叶片,用无离子水洗净,剪碎加入10 ml无离子水,自然浸泡6 h,用DDS-11A型电导率仪测定溶液的电导率EC1,沸水浴10 min,冷却后测定电导率EC2,无离子水电导率值为 EC0,质膜透性 =(EC1-EC0)/(EC2-EC0)×100%。
1.2.9 镉含量测定 将镉处理的海雀稗取出,用去离子水冲洗干净,清除表面水分,于105℃下杀青20 min,然后在80℃下烘干至恒质量,称取样品0.5 g,置于100 ml聚四氟乙烯材料的微波消解罐中,加入4 ml硝酸(北化65%BV-III微电子级)和2 ml双氧水(北化30%BV-III微电子级),盖好密封,按照预设的微波消解条件进行消化处理,将消解后的溶液移至石英试管中,在天平上定质量至20 g,摇匀待测,利用美国生产的安捷伦ICP-MS7500ce电感耦合等离子体质谱仪测定镉含量。
1.2.10 数据分析 试验数据用Excel 2003整理、绘制图标;用SPSS13.0进行方差分析和多重比较。
2 结果与分析
2.1 镉胁迫对海雀稗生长的影响
为了研究镉胁迫对海雀稗生长的影响,本研究测定了不同镉浓度胁迫下海雀稗的形态指标(株高、根长、茎叶鲜质量、根鲜质量)。结果(表2)发现:镉浓度0~50 mg/kg时,各性状指标下降不显著;镉浓度为100 mg/kg时,株高、根长未受到显著抑制,根鲜质量、茎叶鲜质量和分枝数较对照显著降低,相对于对照分别下降了28.76%、46.62%、63.57%;当镉浓度为200 mg/kg时,各项生长指标显著下降,海雀稗生长受显著抑制,但仍能完成生活史。通过分析可知,分枝数与根鲜质量、茎叶鲜质量存在极显著相关性,相关系数分别为0.963**和0.977**,分枝数的减少在一定程度上严重影响海雀稗生物量。

表2 镉胁迫对海雀稗生长的影响Table 2 Effect of Cd stress on biomass of Paspalum vaginatum Sw.
2.2 镉胁迫对海雀稗膜脂过氧化产物丙二醛(MDA)含量的影响
丙二醛(MDA)作为生物膜氧化的产物,其含量的高低代表植物受逆境胁迫伤害程度的大小。根中MDA积累量随镉处理浓度的增加而上升,当镉浓度≤1 mg/kg时海雀稗根中MDA积累量未显著增加;当镉浓度≥10 mg/kg时海雀稗根中MDA积累量随镉浓度的增加显著升高;当镉浓度达200 mg/kg时,海雀稗根中MDA积累量为对照的29.51倍。当镉浓度≤50 mg/kg时茎中MDA未显著积累;当镉浓度达到100 mg/kg时海雀稗茎中MDA含量显著增加;在海雀稗茎中,当镉浓度增加到200 mg/kg,MDA积累量是对照的4.52倍。海雀稗叶片MDA积累量在镉浓度≤10 mg/kg时无显著变化;当镉浓度为50 mg/kg时,MDA在海雀稗叶片迅速积累;当镉浓度达到200 mg/kg时,海雀稗叶片中MDA积累量达到最大,是对照的1.59倍(图1)。

图1 镉胁迫下海雀稗根、茎、叶中丙二醛(MDA)含量的变化Fig.1 Effect of Cd stress on MDA content in the roots,stems and leaves of P.vaginatum Sw.
2.3 镉胁迫对海雀稗叶片细胞膜透性的影响
细胞膜透性是评价植物对污染物反应的指标之一,对细胞内外物质的交换运输具有调节作用。重金属离子与胞膜相结合,会改变膜蛋白的磷脂结构,细胞膜系统受到破坏,细胞内可溶物外渗,进而电导率增加。海雀稗叶片电导率检测结果(图2)显示,在镉处理浓度≤10 mg/kg时,叶片电导率有所增加,但变化不显著;镉浓度为50 mg/kg时,电导率上升15.3%,与对照差异显著;当镉浓度增加到 200 mg/kg时其叶片电导率增加了26.0%,与对照差异显著。说明低浓度镉处理下海雀稗叶片的细胞膜系统未受到损伤,而镉浓度>50 mg/kg时,海雀稗叶片电导率显著增加,细胞膜系统受到损伤。
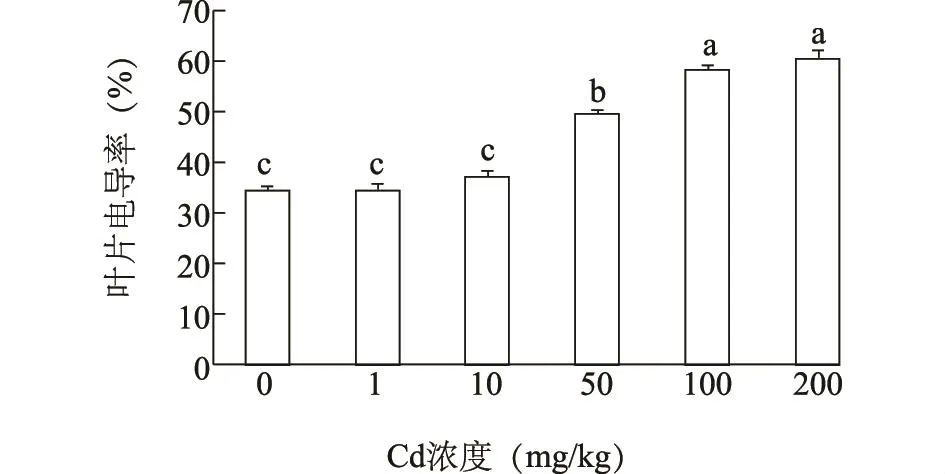
图2 镉胁迫对海雀稗叶片电导率的影响Fig.2 Effect of Cd stress on the leaf electrical conductivity of P.vaginatum Sw.
2.4 镉胁迫对海雀稗保护酶(SOD、CAT)活性的影响
2.4.1 镉胁迫对海雀稗SOD活性的影响 海雀稗中SOD活性变化见图3,镉浓度为1 mg/kg时海雀稗根、茎、叶中SOD活性增加不显著;镉浓度为10 mg/kg时SOD活性在海雀稗根、茎、叶中显著增加,SOD活性分别是对照的1.60倍、2.18倍、2.25倍;镉浓度为200 mg/kg时,海雀稗根和叶片中SOD活性达到最大值,分别是对照的4.92倍、5.58倍。而在茎中当镉浓度为50 mg/kg时,SOD活性达到最大值,其活性是对照的2.34倍,此后随镉浓度增加,茎中SOD活性显著下降。
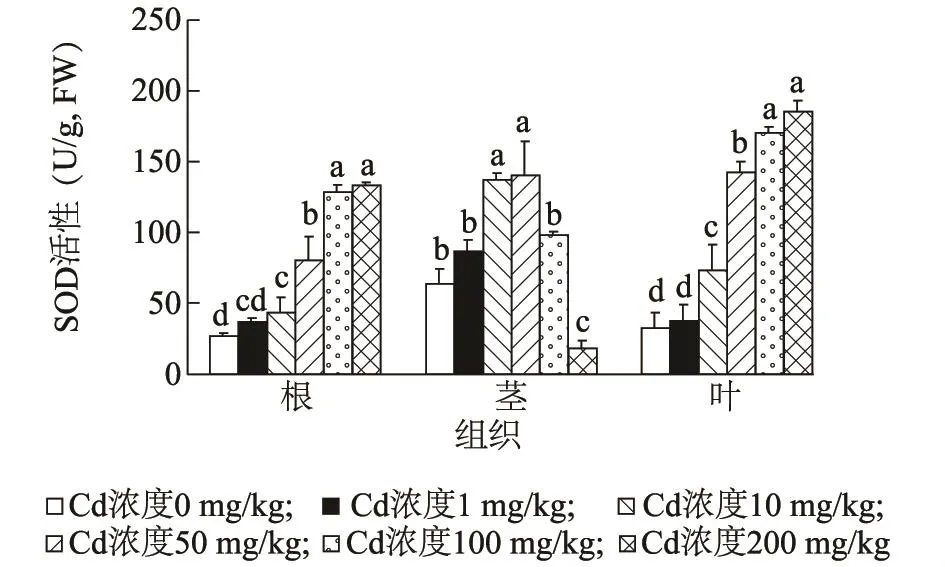
图3 镉胁迫对海雀稗SOD酶活性的影响Fig.3 Effect of Cd stress on SOD activity in P.vaginatum Sw.
2.4.2 镉胁迫对海雀稗CAT活性的影响 CAT能有效清除过氧化氢对细胞的氧化作用,是植物保护自身免受羟基自由基毒害的关键酶。通过研究镉胁迫对海雀稗CAT活性的影响,发现海雀稗根、茎、叶CAT活性随镉浓度的升高呈先升高后下降的趋势(图4)。当镉浓度为1 mg/kg时,海雀稗根和叶片中CAT活性增加不显著,而在海雀稗茎中CAT活性显著增加;当镉浓度≥10 mg/kg时,根、茎、叶中CAT活性随着镉浓度的增加而显著上升;当镉浓度达到50 mg/kg时,各组织中CAT活性达到最大,分别为对照的2.42倍、19.71倍、4.59倍。当镉浓度≥100 mg/kg时,植物体内CAT活性随镉浓度增加而显著下降,但海雀稗叶片中CAT活性较对照相比,维持在较高水平。
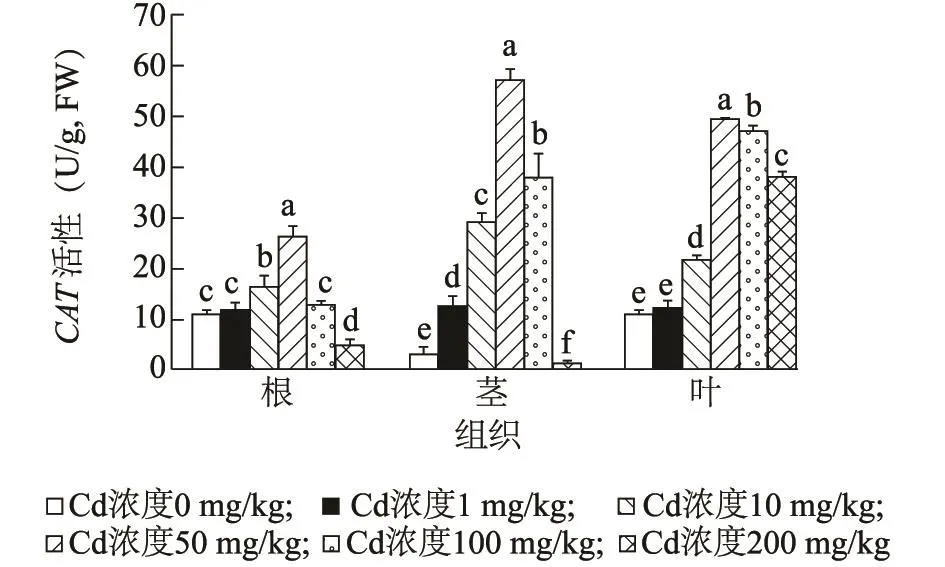
图4 镉胁迫对海雀稗CAT酶活性的影响Fig.4 Effect of Cd stress on CAT activity in P.vaginatum Sw.
2.5 镉胁迫对海雀稗根系活力的影响
根系是植物吸收养分和水分的主要器官,同时也是最先受到镉胁迫的部位,根系活力的强弱直接影响地上部分的生长。如图5所示,当镉浓度为1 mg/kg时,海雀稗根系活力显著上升,其根系活力为对照的1.20倍,此后随镉处理浓度的增加根系活力开始下降。
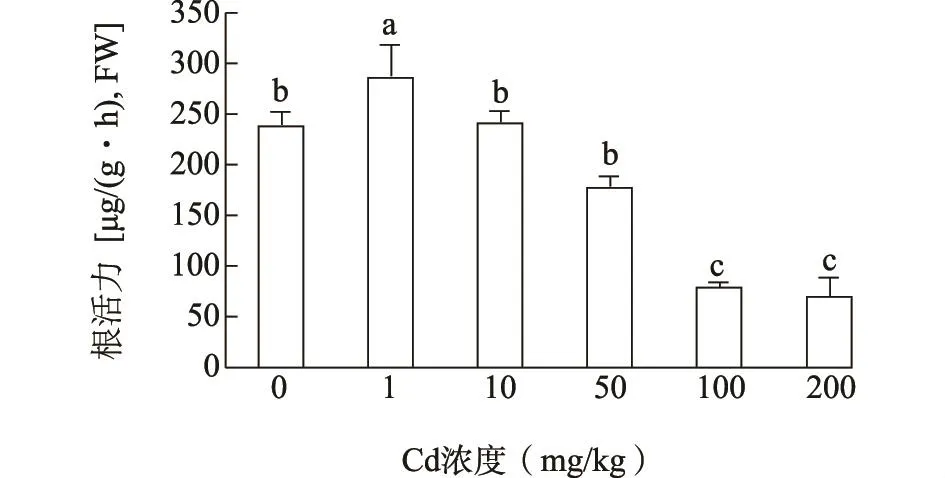
图5 镉胁迫对海雀稗根系活力的影响Fig.5 Effect of Cd stress on the root vigor of P.vaginatum Sw.
2.6 镉胁迫对海雀稗镉积累特征的影响
镉在海雀稗体内的富集特征如表3所示,随镉胁迫浓度增加,海雀稗地上部分和地下部分镉含量呈增加趋势,但地上部分的增加值远小于地下部分,地下部分镉富集量在各浓度下变化显著。地上部分当镉浓度>1 mg/kg时,富集系数为0.16~0.31,镉的富集量较低;地下部分在镉处理后,其富集系数为3.92~14.42,根中滞留率>90%,镉转移系数<0.10,大量的镉被富集于根部。

表3 镉在海雀稗体内的富集特征Table 3 Enrichment characteristics of Cd in P.vaginatum Sw.responding to Cd treatments
3 讨论
3.1 海雀稗体内MDA含量与耐镉性
重金属对植物的胁迫达到一定程度,细胞内活性氧会增加,促进膜脂过氧化作用,破坏细胞膜系统,膜脂过氧化产物丙二醛(MDA)含量增加,细胞膜透性增加,植物的生长受到抑制,表现出相应的毒害症状。如:张超兰等[23]研究镉对4种湿地植物胁迫发现MDA含量随镉浓度增加而升高;王正秋等[24]通过镉对芦荟的胁迫发现,镉胁迫会促使膜透性增大、MDA含量增加。本研究也获得了相似的研究结果,海雀稗MDA含量随镉浓度的增加呈上升趋势,镉浓度≥10 mg/kg时,根系中MDA含量显著增加,茎、叶片中MDA含量分别在镉浓度为100 mg/kg、50 mg/kg才显著增加,表明镉胁迫对海雀稗根、茎、叶的伤害程度不同,根系对重金属镉较为敏感,海雀稗各器官对镉胁迫的忍耐程度为茎>叶>根。
3.2 海雀稗体内SOD、CAT活性的变化与耐镉性
重金属胁迫导致植物活性氧代谢失衡,损伤细胞膜系统,是重金属产生毒性的一种机制,植物体内的抗氧化酶系统对于维持活性氧平衡有重要作用[25]。CAT可以和SOD偶联,彻底清除体内活性氧自由基,保护膜系统[26]。赵钢等[27]、朱启红等[28]、陈朝明等[29]、苑丽霞等[30]对苦荞、蜈蚣草、桑树、油菜的研究发现,低浓度重金属胁迫会促进SOD、CAT等保护酶活性增强,有助于清除体内活性氧自由基,减轻重金属对植物的毒害作用,高浓度重金属胁迫会使SOD、CAT等保护酶活性降低或失活,进而不能清除活性氧自由基对植物的毒害,植物生长受到抑制。这与本研究结果相似,在海雀稗根、叶片中SOD活性随镉浓度的增加而上升,而在茎中SOD活性在镉浓度为50 mg/kg时达到最大,之后开始明显下降,可能是因为茎中活性氧的过量积累抑制了SOD活性。CAT活性随镉浓度的增加呈先上升后降低的变化,镉浓度为50 mg/kg时,CAT活性达到最大,可能是低浓度镉胁迫下海雀稗体内活性氧积累增加导致CAT活性的提高,有利于对活性氧的清除,因而此时根系活力维持在较高水平,植株正常生长,海雀稗对镉的耐受能力增强。当镉浓度>50 mg/kg时,活性氧的过量积累导致CAT活性受到抑制,活性氧平衡受到破坏,从而使海雀稗表现出相应的镉毒害症状,生物量、根系活力降低,MDA含量显著增加,生长受到抑制。
3.3 海雀稗镉的富集特征
镉向海雀稗地上部分转运能力较弱,镉胁迫下海雀稗根系滞留率>90%,转移系数、地上部分富集系数、富集量较小,大量的镉积累于海雀稗根部,而从镉胁迫与植物生长来看,海雀稗对镉胁迫具有一定的耐性。蓖麻在镉浓度为200 mg/kg时,其根部镉含量仅为521.50 mg/kg[31]。有研究发现豆科植物镉迁移到茎叶中的量仅占根吸收的2%[32],这些研究结果与本研究结果相似,而有研究报道胡萝卜和烟草叶片中镉富集量高于根[33],说明镉在植物中的积累分布与植物种类有关。
综上所述,当镉浓度≤50 mg/kg时,海雀稗株高、根长、植株鲜质量未明显下降,根系活力未显著下降,植物体内保护酶活性(SOD、CAT)随镉浓度的增加而增强,能有效清除植物体内的活性氧,活性氧代谢平衡未受到破坏,膜脂过氧化产物(MDA)的积累和膜透性未明显增加,此时海雀稗细胞膜系统未受到严重损伤,海雀稗生长正常。镉浓度>50 mg/kg时,随镉浓度的增加,植物体内产生更多的活性氧,CAT活性显著下降,SOD与CAT的协同作用大大降低,不能维持植物体内的活性氧平衡,植物细胞膜受到破坏,MDA大量积累,根系活力下降,海雀稗生长受到显著抑制。海雀稗根系对镉具有很强的富集能力,可利用海雀稗固定镉污染土壤中的镉,减轻土壤中镉的淋溶及污染土壤的流失,控制镉污染的扩散。因此海雀稗可以作为重金属镉污染土壤修复的候选植物。
[1] 刘玮宁,张炜哲.罗锡文院士谈因污染而导致的食品安全问题:3亿亩地遭重金属污染广东仅一成耕地幸免[N].羊城晚报,2011-10-12(A3).
[2] 佟 讯.全国土壤污染状况调查公报发布[N].中国国土资源报,2014-4-18(1).
[3] 刘 杰.紫苏耐镉胁迫的生理响应及其土壤重金属镉修复潜力评价[D].福州:福建农林大学,2012.
[4] SHAH K,KUMAR R G,VERMA S,et al.Effect of cadmium on lipid peroxidation,superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings[J].Plant Science,2001,161(6):1135-1144.
[5] YI TING HSU,CHING HUEI KAO.Toxicity in leaves of rice exposed to cadmium is due to hydrogen peroxide accumulation[J].Plant and Soil,2007,298(1-2):231-241.
[6] CHAOUI A,MAZHOUDI S,GHORBAL M H,et al.Cadmium and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in bean(Phaseolus vulgaris L.)[J].Plant Science,1997,127(2):139-147.
[7] 孟 丽,李德生,李海茹,等.重金属镉对日本楤木叶绿素和保护酶活性的影响[J].江苏农业科学,2013,41(8):178-180.
[8] 郑爱珍,刘传平,沈振国.镉处理下青菜和白菜MDA含量、POD和SOD活性的变化[J].湖北农业科学,2005(1): 67-69.
[9] 彭 艳,李 洋,杨广笑,等.铝胁迫对不同小麦SOD、CAT、POD活性和MDA含量的影响[J].生物技术,2006,16(3): 38-42.
[10]李丽锋,卢 佳,苏芳莉.镉(Cd)胁迫对芦苇叶片SOD、POD活性影响及动力学分析[J].沈阳农业大学学报,2014,45 (3):326-330.
[11]罗小波,向佐湘,胡立群.09-1海滨雀稗草坪坪用性状评价[J].作物研究,2013,27(S1):57-61.
[12]王 恺,刘一明,王兆龙.假俭草和海滨雀稗对土壤铅污染胁迫的生理反应[J].草业科学,2010,27(2):32-38.
[13]王 恺.假俭草和海滨雀稗对土壤Pb、Cd、Zn污染胁迫的响应及耐受阈值研究[D].上海:上海交通大学,2010.
[14]朱燕华.草坪植物对铅的耐性及富集特性研究[D].扬州:扬州大学,2007.
[15]孙约兵,周启星,王 林,等.三叶鬼针草幼苗对镉污染的耐性及其吸收积累特征研究[J].环境科学,2009,30(10): 3028-3035.
[16]张宪政,陈风玉,王荣富.植物生理学实验技术[M].沈阳:辽宁科学技术出版社,1994:99-100
[17]郑炳松.现代植物生理生化研究技术[M].北京:气象出版社,2006.
[18]李合生,孙 群,赵世杰,等.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[19]郝建军,康宗利,于 洋.植物生理学实验技术[M].北京:化学工业出版社,2007.
[20]彭志英,蒋 黎.紫外速率直接法测定过氧化氢酶活性[J].华西医学,1995,10(1):4-7.
[21]刘 宁,高玉葆,贾彩霞,等.渗透胁迫下多花黑麦草叶内过氧化物酶活性和脯氨酸含量以及质膜相对透性的变化[J].植物生理学通讯,2000,36(1):11-14.
[22]陈建勋,王晓峰.植物生理学实验指导[M].2版.广州:华南理工大学出版社,2006:64-66.
[23]张超兰,陈文慧,韦必帽,等.几种湿地植物对重金属镉胁迫的生理生化响应[J].生态环境,2008,17(4):1458-1461.
[24]王正秋,江行玉,王长海.铅、镉和锌污染对芦苇幼苗氧化胁迫和抗氧化能力的影响[J].过程工程学报,2002,2(6): 558-563.
[25]PALACE V P,BROWN S B,BARON C L,et al.An evaluation of the relationships among oxidative stress,antioxidant vitamins and early mortality syndrome(EMS)of lake trout(Salvelinus namaycush)from lake ontario[J].Aquatic Toxicology,1998,43 (2):195-208.
[26]张金彪,黄维南.镉对植物的生理生态效应的研究进展[J].生态学报,2000,20(3):514-523.
[27]赵 钢,邹 亮,彭镰心,等.铅胁迫对苦荞生理特性的影响[J].江苏农业科学,2012,40(7):98-100.
[28]朱启红,夏红霞.铅胁迫对蜈蚣草抗氧化酶系统和叶绿素含量的影响[J].贵州农业科学,2012,40(4):56-58.
[29]陈朝明,龚惠群,王凯荣.Cd对桑叶品质、生理生化特性的影响及其机理研究[J].应用生态学报,1996,7(4):417-423.
[30]苑丽霞,孙 毅,杨艳君.镉胁迫对油菜生长发育中生理生化特性的影响[J].安徽农业科学,2014(9):2544-2547,2558.
[31]陈亚慧,刘晓宇,王明新,等.蓖麻对镉的耐性、积累及与镉亚细胞分布的关系[J].环境科学学报,2014,34(9):2440-2446.
[32]李彦娥,赵秀兰.植物镉积累和耐性差异研究进展[J].微量元素与健康研究,2004,21(3):53-56.
[33]CATALDO D A,GARLAND T R,RE W.Cadmium distribution and chemical fate in soybean plants.[J].Plant Physiology,1981,68(4):835-839.
猜你喜欢
四川劳动保障(2021年3期)2021-06-09
疯狂英语·初中天地(2019年12期)2020-01-04
时代人物(2019年29期)2019-11-25
疯狂英语·初中版(2019年12期)2019-01-02
小哥白尼(野生动物)(2018年3期)2018-06-15
现代园艺(2017年13期)2018-01-19
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
山东医药(2015年38期)2015-12-07
食品工业科技(2014年13期)2014-03-11
