
黄河源区沼泽湿地退化过程中植物群落特征的变化
2015-04-10 07:29任国华
草业科学 2015年8期
任国华,邓 斌,后 源
(1.山西农业大学动物科技学院,山西 太谷030801;2.草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020)
湿地是分布于水生生态系统与陆地生态系统之间的特殊生态系统[1],它不仅是陆地上的天然水库,具有重要的涵养水源和调蓄水资源功能,也是地球上最活跃和最富生物多样性的生态景观[2-3]。黄河源区由于独特的地理条件形成了类型多样的高原湿地生态景观[4],是青藏高原湿地重要的集中分布区,对流域内的生态水文过程与水循环有重要影响[5],同时它也是青藏高原生产力最高的天然草地生态系统[6]。因此,黄河源区高寒湿地资源的分布和格局变化不仅对当地社会经济产生直接影响,也对青藏高原生物多样性保护及生态平衡的维持具有重要作用。近几十年来,在全球气候变化的大背景下[7-8],加之过度放牧、随意采挖、开沟排水和鼠害肆虐等因素的干扰[9],使得源区湿地萎缩退化十分严重[10],植被类型发生更替,群落结构和生物多样性发生改变,毒杂草比例增加[11-12]。高寒湿地退化萎缩是黄河源区生态环境恶化的重要标志之一,已经引发了一系列生态环境问题[13-14]。因此,对黄河源区高寒湿地萎缩退化过程中植物群落空间分布特征的研究,有利于揭示湿地退化过程中植被分异规律,从而为高寒湿地的优化管理、保护和恢复提供基础资料。
1 研究地区与方法
1.1 自然概况
研究区位于青海省东南部果洛藏族自治州玛沁县境内的高寒草甸生态系统试验站(34°26' -34°28' N,100°12' -100°14' E),平均海拔3 800 m。该地区具有典型的高寒大陆性气候特征,年平均气温-4 ~-1 ℃,最冷月1 月和最热月7 月平均气温分别为-14 ~-12 ℃和9 ~13 ℃。年均降水量在510 ~550 mm 之间,80%以上集中在6 -9 月份,年均蒸发量2 400 mm 左右。土壤以沼泽草甸土、高山草甸土和高山灌丛草甸土为主。
1.2 群落调查
在自然和人为因素的综合作用下,青海三江源地区湿地环境呈逆向演替,尽管不同类型湿地演替序列复杂多变,但湿地及其周边植被退化演替进程一般为沼泽湿地→高寒草甸→退化高寒草甸→荒漠化地区[15]。本研究中湿地退化演替样地调查区设置在青海省玛沁县大武镇河漫滩湿地,研究区地势低平,地表径流不畅,形成大面积的沼泽湿地。20世纪80 年代中后期以来,该区连片的沼泽湿地发生了明显的萎缩退化现象,湿地面积减小甚至外围已呈零星分布[2-3]。本研究以现生长季可见的沼泽湿地中心区域向外延伸的取样方法,选取各退化序列(依次为沼泽湿地、沼泽化草甸、高寒草甸和退化高寒草地)的典型群落样地。每个退化序列设置5 个10 m×10 m 的取样样地,在每个取样样地内随机设置3 个1 m×1 m 样方,共计20 个取样样地,60 个取样样方。野外样方调查于2010 年7 ~8 月该区植物生长旺盛期进行,分别调查每个样方内物种种类、数量及每个物种的盖度和高度;地上生物量采用烘干称重法,将样方内植物距地面1 cm 处收获并烘干至恒重,并按经济类群(莎草科、禾草科和杂类草)进行分类。
1.3 多样性测定及方法
群落中植物多样性以物种丰富度指数、Simpson优势度指数、Pielou 均匀度指数和Shannon-Wiener多样性指数表示,群落中植物多样性根据物种重要值计算[16]:
重要值= (相对盖度+ 相对高度+ 相对生物量)/3;
丰富度指数:

Simpson 优势度指数:

Pielou 均匀度指数:

Shannon-Wiener 指数:
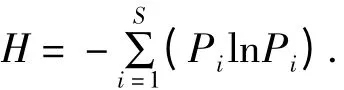
式中,S 为样方内物种数目;Pi为物种i 的相对重要值,Pi=Ni/N,Ni为物种i 的绝对重要值,N 为物种i所在样方内所有物种重要值之和。
1.4 模型介绍
(1)Null 断棍模型(Broken-Stick Model),群落中等级为i 的物种多度(Ni)可以表示为:

式中,J 表示群落中物种个体数,S 表示物种数目,x为从i 到s 中取值。
(2)Preempt 生态位优先模型(Niche Preemption Model),其等级i 物种多度(Ni)可以表示为:

式中,N1为模型拟合的群落中最丰富物种的多度,k为物种占用总生态位的份数。
(3)Log-normal 对数正态分布模型(Log-Normal Model),其假定物种个体数取对数后符合正态分布,即物种多度分布模式可以表示为:

式中,μ 为正态分布的均值,δ 为正态分布的方差,Φ为正态偏差。
(4)Zipf 模型(Zipf Model),该拟合模型实际上是对群落演替过程的一种反映,即后来定居物种相对先前到达物种更难存活。

式中,N 为群落中物种总的个体数,P1表示预测物种最丰富植物的多度占群落总个体数比例,γ 代表了物种出现可能性的平均值,为常数。
(5)Zipf-Mandelbrot 模型(Zipf-Mandelbrot Model),是在Zipf 模型中增加了一个参数C,以取代模型预测的最多物种的多度比例P1,多了参数β。
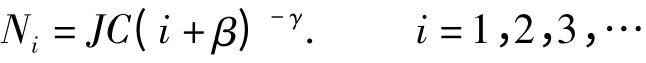
式中,J 表示群落中物种个体数,C 和β 表示为非线性参数。
1.5 数据处理
所有数据均采用Excel 录入并进行物种重要值及多样性指数计算;利用统计软件SPSS 18.0中的ANOVA 进行方差分析,Tukey’s 进行多重比较;在R-3.1.3 的vegan 软件包中进行物种多度分布模式分析并绘制拟合曲线[17-19]。
2 结果与分析
2.1 沼泽湿地退化过程中群落特征变化
调查结果显示,在4 个退化演替序列中,共记录到87 种植物,分属25 科69 属。其中菊科植物13种,龙胆科10 种,毛茛科和禾本科均为9 种,分别占14.9%、11.5%和10.3%。湿地退化过程中群落组成变化明显(表1),演替早期群落优势种为藏嵩草和华扁穗草(Blysmus sinocompressus),主要伴生种是矮金莲花(Trollius pumilus)、盘花垂头菊(Cremanthodium discoideum)、鹅绒委陵菜(Potentilla anserine)和草地早熟禾(Poa pratensis)等。随着沼泽化草甸逐渐演变为高寒草甸,原生植物在群落内的地位和作用逐渐减弱直至消失,典型的高原地带性植物矮嵩草(Kobresia humilis)和高山嵩草(K. pygmaea)在群落内居于主导地位。至退化高寒草地阶段,群落优势种为黄帚橐吾(Ligularia virgaurea),主要伴生种为铁棒锤(Aconitum pendulum)、高山嵩草、细叶亚菊(Ajania tenuifolia)和细裂叶莲蒿(Artemisia gmelinii),毒杂草种类和重要值具有明显优势,说明其在群落内的地位和作用较强,具有进一步扩张的趋势(表1)。
2.2 沼泽湿地退化过程中生产力和物种多样性变化
本研究中,沼泽湿地群落地上生物量最高,为515 g·m-2;随着湿地的退化,群落生物量迅速下降,并在高寒草甸阶段降至最低值,为307 g·m-2左右(图1)。各演替阶段,不同功能群植物生物量所占比重发生了明显变化(图1),沼泽湿地和沼泽化草甸都是莎草科植物占据绝对优势,分别占总量的87.6%和65.4%;而随着退化演替的进行,杂类草生物量所占比重逐渐增加,在高寒草甸和退化高寒草地中分别占45.6%和79.9%。
黄河源区沼泽湿地退化过程对植物群落间物种丰富度、优势度、均匀度和多样性均有影响。演替初期,群落内物种数目较少,物种丰富度、多样性和均匀度指数较低,而优势度指数较大,说明藏嵩草作为群落的主要建群种占据优势地位和作用。随着退化演替进展,群落样方内物种数目逐渐增多,均匀度和多样性指数逐渐增大,优势度指数不断减小,说明群落内物种影响群落组织水平和功能的作用差别逐渐减弱(表2)。
2.3 沼泽湿地退化过程中物种-多度分布模式变化
沼泽湿地退化过程中植物群落物种多度等级分布拟合如图2 所示,根据每个模型的最小信息准则(Akaike’s Information Criteria,AIC)选择最优模型(表3)。沼泽湿地退化过程影响群落物种多度等级分布模式,演替早期对数正态分布模型与实际物种多度分布吻合较好;生态位优先占领模型和Zipf-Mandelbrot 模型更适合描述高寒草甸阶段群落物种多度分布格局;而在退化草地阶段符合Zipf-Mandelbrot 模型(表3 和图2)。

表1 黄河源区沼泽湿地退化过程中主要物种重要值变化Table 1 Changes of species composition and important values along the successional gradient of the swamp wetland in the Yellow River Source

图1 黄河源区沼泽湿地退化过程中群落生产力及不同功能群植物所占比例变化趋势Fig.1 Changes of productivity and the percentage of plant functional groups along the successional gradient of theswamp wetland in the Yellow River Source
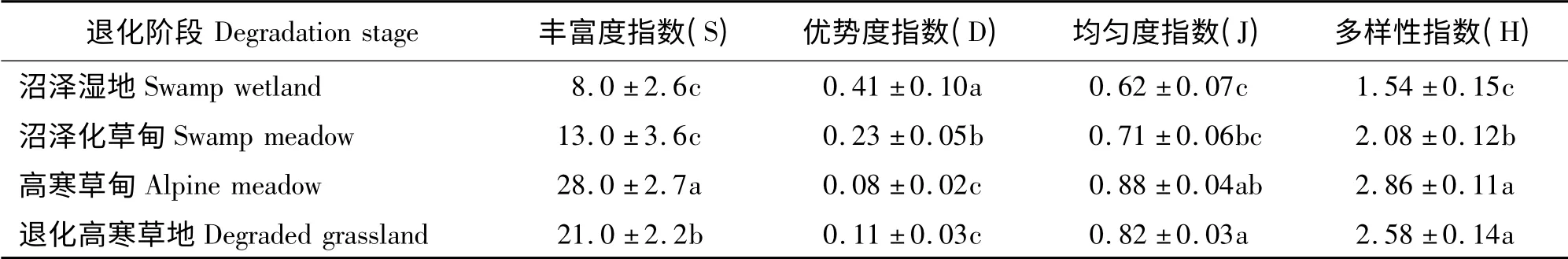
表2 沼泽湿地退化过程中物种丰富度指数(S)、优势度指数(D)、均匀度指数(J)和多样性指数(H)的变化Table 2 Changes of species number(S),domination index (D),evenness index (J)and Shannon-Weiner index (H)along the successional gradient of the swamp wetland in the Yellow River Source
3 讨论
3.1 群落结构和多样性变化
黄河源区高寒湿地的3 种主要类型(河漫滩湿地、山前湿地和湖泊湿地)在全球气候变化和人类活动干扰下均出现了不同程度的退化[2-3]。在湿地退化过程中,群落组分更替明显,总体上物种组成呈现水生→湿生→中生→旱生的演化模式[6]。本研究中原建群种藏嵩草和华扁穗草在群落内的地位随着沼泽湿地的退化逐渐减弱,而矮嵩草和小嵩草等旱中生植物则演变为群落主要组分种,群落物种丰富度和多样性迅速提高。这与黄河首曲湿地[6]和祁连山海北湿地[20]退化过程中的物种多样性和植物群落结构的变化趋势相一致,说明高寒湿地退化后形成的暖干生境已经不适宜原生湿生性植物的生长,却为生长潜势被抑制的中旱生植物生长创造了有利条件[21-23]。随着湿地退化程度的继续加剧,群落原生物种被杂类草取代并逐渐退出群落,因此植物多样性相对于演替中期的群落而言略有下降。

表3 黄河源区沼泽湿地退化过程中5 种模型拟合的AIC 值Table 3 AIC of five models at different succession stages of the swamp wetland in the Yellow River Source

图2 黄河源区沼泽湿地退化过程中物种-多度模式分布及模型拟合Fig. 2 Species-abundance distribution and fitting of five models at different succession stages of the swamp wetland in the Yellow River Source
3.2 生产力变化
高寒湿地群落内物种随着退化演替时间的推移而有所更替,不仅影响群落结构和多样性特征,而且影响生产力的变化[24]。退化演替早期阶段,植物群落中以克隆植物藏嵩草、华扁穗草和粗喙苔草等莎草科植物为主,包含少量的草地早熟禾,莎草和禾草所占比例为95.7%,杂草种类和生物量较小,植被盖度高,放牧干扰少,其累积的地上生物量较高[25]。随着沼泽湿地积水面积的减少,放牧干扰逐渐增大,牲畜啃食与践踏导致植被高度和盖度减小,地上生物量也随之减少[26];至退化草地阶段,由于原生植物的基本退出和地表秃斑裸地的增多[16,27],群落总生物量值最低,而禾本科和莎草科牧草生物量所占比例仅为20.1%,草地质量下降,生产性能丧失。
3.3 物种多度格局变化
在群落物种组成多度变化分析中,通常采用代表不同生态机理的数学模型来拟合物种的这种分布模式,以此揭示群落内物种多度格局产生和维持的生态学过程[17,28-29]。湿生性植物由于主要分布在排水不畅的潮湿地带,对水淹环境和低温具有较高的抗逆性,具有较好的资源优势和适应性,因而在沼泽湿地阶段符合对数正态模型。随着沼泽湿地水分的减少和土壤暖干化程度的加剧,中生性植物的生境适合度增加,占领生境中的空间和资源,生境过滤和生态位分化作用增强[30],代表生态位过程的Zipf-Mandelbrot 模型和生态位占领模型对此阶段物种多度分布格局有较好的拟合。当演替至退化草地阶段时,Zipf-Mandelbrot 模型拟合物种多度格局效果最好,该假说认为在植物群落中,一个物种的存在取决于生境先前的物理条件和群落中已经存在的物种,先锋物种入侵付出的代价较少,即要求的先决条件很少;其后到达的物种侵入过程则要付出较高的代价,对时间、能量以及生态系统的结构都有所要求,所以个体数目相对稀少,即后来定居物种相对先前到达物种更难存活,实际上是对群落演替过程的一种反映[17-18]。Zipf-Mandelbrot 模型较好地说明了在沼泽湿地演替后期,退化草地次生毒杂草逐渐取代中生性先锋种而成为群落优势种的生态学过程[22,31]。总体来说,在黄河源区高寒沼泽湿地萎缩退化过程中,群落物种多度分布模式由对数正态型分布经生态位优先占领型逐渐转变为Zipf-Mandelbrot 型分布。
4 结论
黄河源区沼泽湿地退化过程中生境逐渐干暖化,部分中旱生植物的侵入致使群落内物种丰富度和多样性指数增加,但降低了优势度指数并改变了群落内物种多度分布格局,即湿地退化增加了群落的植物多样性,但增加的植物多为毒杂草,从而会导致高寒湿地向杂草型退化方向演替。因此,沼泽湿地一些排水、改良等工程尽管短期内会增大牧草供给功能,促进畜牧业发展,但长期开发利用会加剧湿地的退化,不利于草地的良性发展和其他服务功能(如水源涵养和调节功能)的正常发挥。
[1] Costanza R,d’Arge R,de Groot R,Farber S,Grasso M,Hannon B,Limburg K,Naeem S,O’Neill R V,Paruelo J,Raskin R G,Sutton P,van den Belt M.The value of the world’s ecosystem services and natural capital[J].Nature,1997,387:253-260.
[2] 潘竟虎,王建,王建华.长江、黄河源区高寒湿地动态变化研究[J].湿地科学,2007,5(4):298-304.
[3] 王根绪,李元寿,王一博,陈玲.近40 年来青藏高原典型高寒湿地系统的动态变化[J].地理学报,2007,62(5):481-491.
[4] 陈桂琛,黄志伟,卢学峰,彭敏.青海高原湿地特征及其保护[J].冰川冻土,2002,24(3):254-230.
[5] 王莺,李耀辉,孙旭映.气候变化对黄河源区生态环境的影响[J].草业科学,2015,32(4):539-551.
[6] 后源,郭正刚,龙瑞军.黄河首曲湿地退化过程中植物群落组分及物种多样性的变化[J].应用生态学报,2009,20(1):27-32.
[7] 张轶鸥,丑洁明,董文杰.中国关于气候变化归因和责任的研究进展与展望[J].干旱气象,2013,31(1):174-181.
[8] 罗磊.青藏高原湿地退化的气候背景分析[J].湿地科学,2005,3(3):190-199.
[9] 李春秀,孙海松.青海省西藏嵩草沼泽化草甸资源及保护[J].草业科学,2009,26(9):77-83.
[10] 陈永富,刘华,邹文涛,张怀清.三江源高寒湿地动态变化趋势分析[J].林业科学,2012,48(10):70-76.
[11] 吴海艳,马玉寿,董全民,孙小弟,施建军,王彦龙,盛丽.黄河源区藏嵩草沼泽化草甸地上生物量及营养季节动态研究[J].草业科学,2009,26(1):8-12.
[12] 张伟,孙海松.黄河源区藏嵩草沼泽化草甸群落结构特征的研究[J].青海大学学报(自然科学版),2015,33(1):1-13.
[13] Dong Q M,Zhao X Q,Wu G L,Shi J J,Ren G H.A review of formation mechanism and restoration measures of“black-soiltype”degraded grassland in the Qinghai-Tibetan Plateau[J].Environmental Earth Sciences,2013,70(5):2359-2370.
[14] Liu J,Xu X,Shao Q. Grassland degradation in the“Three-River Headwaters”region,Qinghai Province[J]. Journal of Geographical Sciences,2008,18(3):259-273.
[15] 黄桂林.青海三江源区湿地状况及保护对策[J].林业资源管理,2005(4):35-39.
[16] 张蕊,王媛,马丽娜,桑潮,王力,郭瑞英,汪海波,尚占环.三江源区退化人工草地、“黑土滩”和天然草地植物群落物种多样性[J].草地学报,2014,22(6):1171-1178.
[17] 程佳佳,米湘成,马克平,张金屯.亚热带常绿阔叶林群落物种多度分布格局对取样尺度的响应[J].生物多样性,2011,19(2):168-177.
[18] 闫琰,张春雨,赵秀海. 长白山不同演替阶段针阔混交林群落物种多度分布格局[J]. 植物生态学报,2012,36(9):923-934.
[19] 文淑均,李伟,杜国祯.围栏封育对青藏高原东缘高寒草甸种子雨的影响[J].草业科学,2012,29(3):333-340.
[20] 李英年,赵新全,赵亮,王启基,沈振西.祁连山海北高寒湿地气候变化及植被演替分析[J].冰川冻土,2003,25(3):243-249.
[21] 赵新全,马玉寿,王启基,刘伟,周立,周华坤,赵亮,施建军,徐世晓,董全民.三江源区退化草地生态系统恢复与可持续管理[M].北京:科学出版社,2011:1-5.
[22] 李媛媛,董世魁,朱磊,温璐,李小艳,王学霞.青藏高原高寒草甸退化与人工恢复过程中植物群落的繁殖适应对策[J].生态学报,2013,33(15):4683-4691.
[23] 邓自发,谢晓玲,王启基,周兴民. 高寒草甸藏嵩草种群繁殖对策的研究[J]. 应用与环境生物学报,2001,7(4):332-334.
[24] Wang C,Long R,Wang Q,Liu W,Jing Z,Zhang L.Fertilization and litter effects on the functional group biomass,species diversity of plants,microbial biomass,and enzyme activity of two alpine meadow communities[J].Plant and Soil,2010,331(1):377-389.
[25] Wu G L,Ren G H,Wang D,Shi Z H,Warrington D.Above-and below-ground response to soil water change in an alpine wetland ecosystem on the Qinghai-Tibetan Plateau,China[J].Journal of Hydrology,2013,476:120-127.
[26] 庞晓攀,贾婷婷,李倩倩,雒眀伟,肖玉,赵旭,郭正刚.高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响[J].生态学报,2015,35(3):873-884.
[27] 尚占环,龙瑞军,马玉寿.江河源区“黑土滩”退化草地特征、危害及治理思路探讨[J].中国草地学报,2006,28(1):69-74.
[28] 李巧,涂璟,熊忠平,卢志兴,刘春菊.物种多度格局研究概况[J].云南农业大学学报,2011,26(1):117-123.
[29] 宋晓谕,张仁懿,李新娥,袁建立,储诚进,王刚.甘南亚高山草甸弃耕演替过程中的物种多样性与生产力变化模式及相互关系研究[J].草业学报,2010,19(6):1-8.
[30] Maire V,Gross N,Börger L,Proulx R,Wirth C,da Silveira Pontes L,Soussana J F,Louault F.Habitat filtering and niche differentiation jointly explain species relative abundance within grassland communities along fertility and disturbance gradients[J].New Phytologist,2012,196:497-509.
[31] 周华坤,赵新全,温军,陈哲,姚步青,杨元武,徐维新,段吉闯.黄河源区高寒草原的植被退化与土壤退化特征[J].草业学报,2012,21(5):1-11.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
环境工程技术学报(2022年3期)2022-06-05
西部资源(2021年1期)2021-11-26
灌溉排水学报(2021年8期)2021-09-02
活力(2019年21期)2019-04-01
西北林学院学报(2018年5期)2018-10-12
下一代英才(酷炫少年)(2017年9期)2017-11-27
河北林业科技(2016年5期)2016-11-08
剑南文学(2016年11期)2016-08-22
