
弗吉尼亚栎幼苗对高温胁迫的生理响应
2015-05-15 05:59杨舒婷李谦盛
江西农业大学学报 2015年1期
杨舒婷,曲 博,2,李谦盛,邓 敏
(1.中国科学院 上海辰山植物研究中心/上海辰山植物园,上海 201602;2.上海应用技术学院 生态学院,上海 201418)
弗吉尼亚栎(Quercus virginiana Mill)为壳斗科栎属常绿乔木,原产于美国东南部,因树冠浓密宽大、容易栽培、幼树生长速度快,在美国东南部城市森林和绿化中普遍栽培[1]。因其根系发达、树干韧性强而抗风耐盐,我国作为沿海防护林树种引进后在长江三角洲沿海栽培示范适应性良好[2]。我国在研究推广过程中重点对弗吉尼亚栎的盐胁迫生理[3-4]和干旱胁迫生理[5]进行了较多研究,而鲜有对弗吉尼亚栎温度胁迫生理的研究报道。温度是影响植物生长的重要环境因子,随着全球气候变暖,夏季极端高温度数和天数都在上升,高温胁迫也成为植物面临的最主要的非生物胁迫之一[6]。树木对高温胁迫的耐受性和适应性是选择园林树种的重要依据之一。本研究通过人工气候箱模拟高温胁迫,对弗吉尼亚栎幼苗叶片的含水量以及生物膜相关的几个生理指标开展研究,探讨其耐热性能,旨在为其人工栽培繁殖及园林应用提供基础资料。
1 材料与方法
1.1 试验材料
种子于2012年11月采自上海辰山植物园内引种的弗吉尼亚栎,选取籽粒饱满、大小基本一致、无虫孔的种子用0.5%的高锰酸钾溶液消毒30 min后晾干,贮存于4℃种子保存箱。2012年12月20日播于32孔林木专用穴盘,基质为80%泥炭+20%珍珠岩的混合基质。2013年2月底开始出苗,4月中旬每穴施用2~4粒18N-6 P2O5-12K2O的控释肥。7月中旬挑选长势基本一致的幼苗作为试验材料。
1.2 试验方法
1.2.1 材料处理 将供试植株从温室转入人工气候箱,在25℃,光照12 h,黑暗12 h,70%湿度条件下培养7 d。选取长势一致的植株,分为3组,一组作为对照(CK),继续置于25℃光照培养箱中生长,其余2组分别置于35℃和45℃光照培养箱中进行高温胁迫处理12,24,48 h,处理过程中保证水分补给充足,各时长处理的1/5幼苗(各4盆)在每个时段的高温处理结束后立即用于测定生理指标,剩余4/5幼苗分别在各处理结束后重新置于25℃、光照12 h、黑暗12 h下恢复生长12,24,48,72 h(各4盆),用于取样测定相关生理指标。取叶片测定生理指标后的植株仍保留于温室内栽培观察。以上试验处理重复3次。
1.2.2 测定方法 叶片含水量的测定:取不同处理相同部位的叶片称鲜质量M1,用信封装好后放入80℃烘箱,烘至恒重,称取干质量M2,每个处理3次重复。叶片含水量=(M1-M2)/M1×100%。电解质外渗率的测定:参考孙国胜等[7]的方法测定叶圆片相对电解质外渗率。丙二醛(MDA)的测定:采用硫代巴比妥酸法测定[8]。脯氨酸的测定:采用酸性茚三酮法测定[8]。
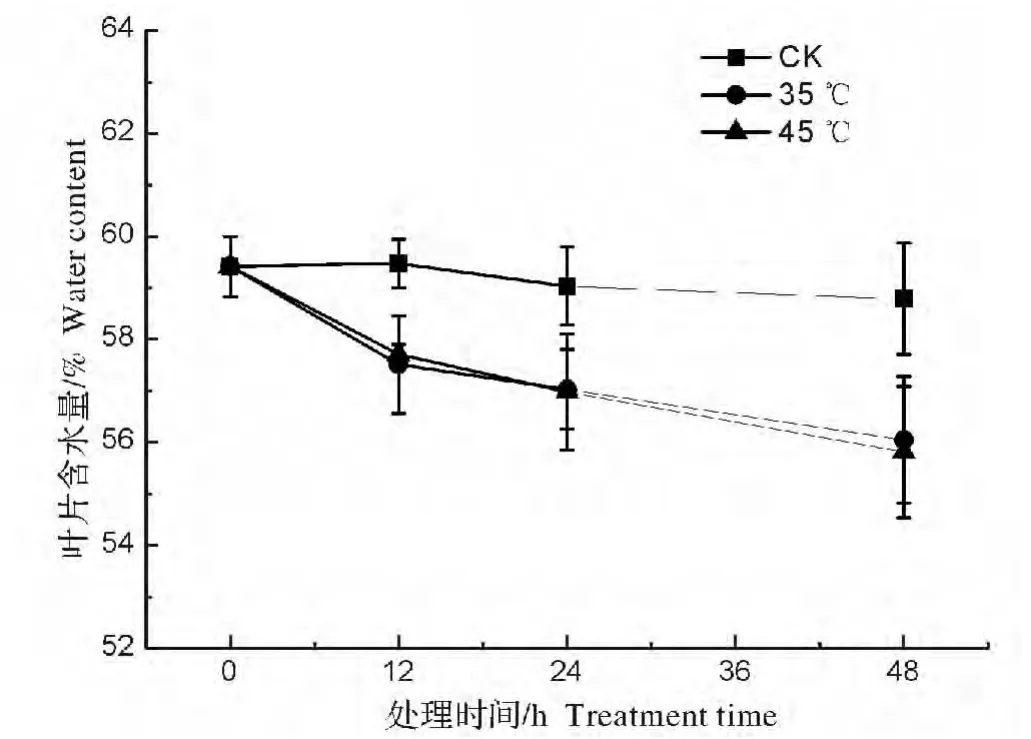
图1 高温胁迫对弗吉尼亚栎叶片含水量的影响Fig.1 Effect of heat stresses on leaf water content of Quercus virginiana
2 结果与分析
2.1 高温胁迫及恢复生长对弗吉尼亚栎幼苗叶片含水量的影响
弗吉尼亚栎幼苗在高温胁迫后,叶片含水量显著下降,并且随着胁迫时间的延长而递减;在高温处理48 h后,叶片含水量从59.42%分别降到了 56.05%(35 ℃)和 55.81%(45℃),显著低于对照组(CK)(图1)。但相同处理时间下两个胁迫温度间的叶片含水量没有显著差异,说明35℃和45℃的胁迫处理对叶片含水量的影响基本一致。
将高温胁迫处理后的植株置于25℃下恢复生长后,弗吉尼亚栎植株的叶片含水量逐渐恢复(图2),经过72 h的恢复生长后其叶片含水量基本可达到处理前水平。但经过45℃高温胁迫处理后的植株(图2B)相比35℃相同时间处理的植株(图2A)其叶片含水量的恢复能力稍弱一些,在恢复生长的这段时间里所测定的数值波动也相对较大。

图2 不同处理阶段弗吉尼亚栎叶片含水量的变化Fig.2 Leaf water content change of Quercus virginiana at different treatment phase

图3 不同温度高温胁迫处理对弗吉尼亚栎叶片电解质外渗率的影响Fig.3 Effect of heat stress on the electrolyte leakage of Quercus virginiana leaves
2.2 高温胁迫及恢复生长对弗吉尼亚栎幼苗电解质外渗率的影响
高温胁迫12 h后,叶片的电解质外渗率均显著高于对照组(图3)。随着胁迫处理时间的延长,植株的电解质外渗率略有回落,但均维持显著高于对照的水平,而35℃和45℃处理之间的电解质外渗率没有显著差异。
将高温处理过的弗吉尼亚栎植株置于25℃下进行恢复生长后,所有处理后的植株在恢复生长的72 h内电解质外渗率一直与胁迫处理后水平相近,没有恢复(图4),表明高温胁迫对弗吉尼亚栎叶片电解质渗透率的伤害是不可逆的。
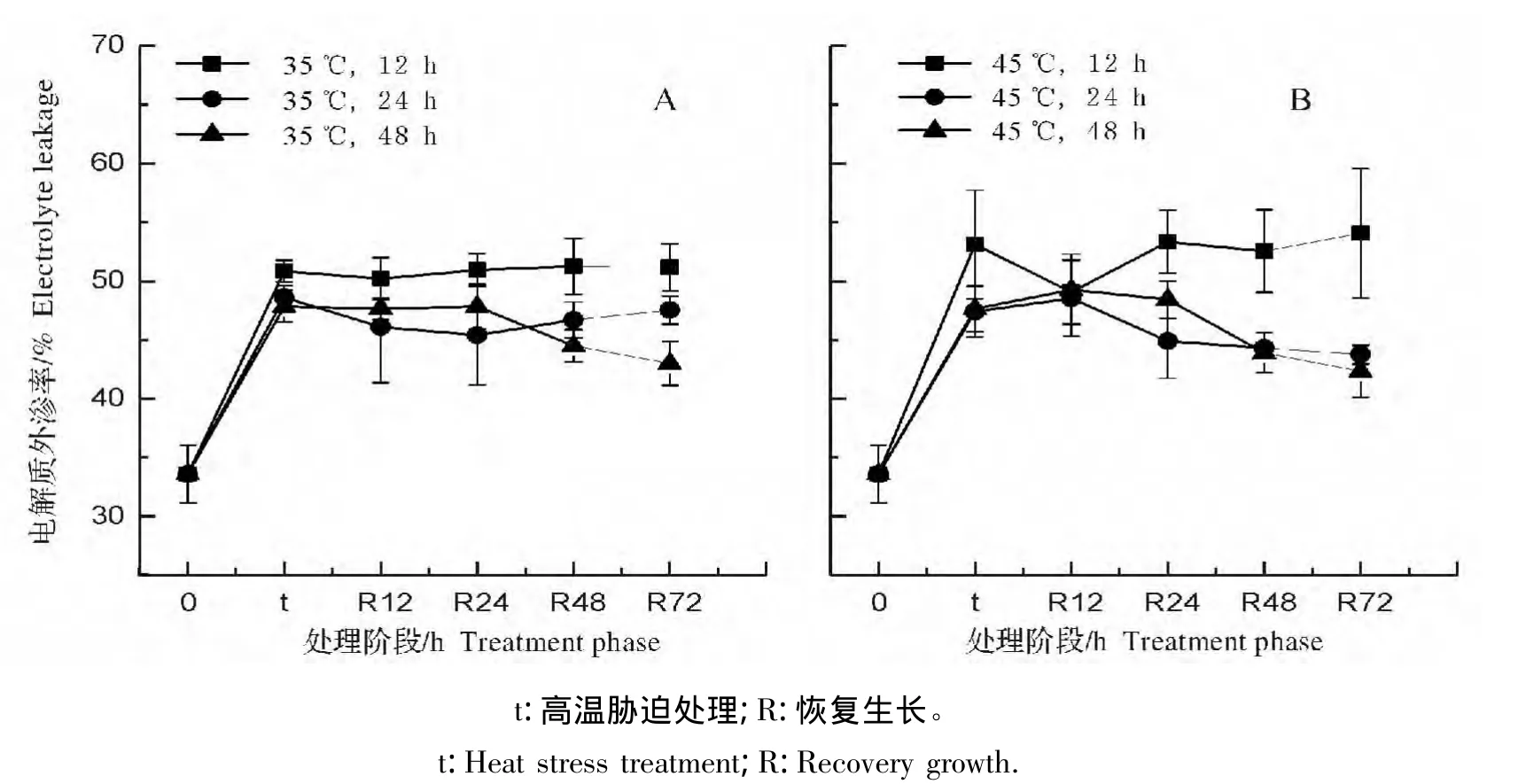
图4 不同处理阶段弗吉尼亚栎叶片电解质外渗率的变化Fig.4 Change of the electrolyte leakage in Quercus virginiana at different treatment phase
2.3 高温胁迫及恢复生长对弗吉尼亚栎幼苗MDA含量的影响
高温胁迫处理后叶片MDA含量均出现先升高后降低的变化,这可能是植株逐渐适应高温环境过程中,激活抗氧化系统,清除掉了一些MDA(图5)。但在35℃高温处理后,MDA含量变化幅度相对较小,随着处理时间延长逐渐升高,在胁迫处理24 h时 MDA 含量达到最高(0.046 μmol/g),在48 h时又下降至对照组相近水平;而在45℃极端温度胁迫下,叶片中的MDA含量在胁迫处理12 h就达到了一个很高的水平(0.082 μmol/g),而后随着处理时间增加,MDA含量又逐步下降到与对照组相当的水平。
在恢复生长期,两种温度处理所表现出来的变化亦有所不同。35℃胁迫处理的植株中MDA含量在恢复生长过程中均逐步下降,其中35℃高温胁迫处理12 h和48 h的植株在经过72 h的恢复生长后,植株中的MDA含量与对照组相比没有显著性差异,而35℃高温处理24 h的植株在经过72 h的恢复生长后植株中的MDA含量仍显著高于其他两组,说明其恢复相对缓慢一些(图6A)。45℃高温胁迫处理12 h和24 h的植株在恢复生长过程中MDA含量的变化趋势分别与35℃高温胁迫处理12 h和24 h的处理组检测结果相似(图6B)。尽管经过45℃,48 h的高温胁迫后,植株出现了一定的适应性,MDA的含量较处理12 h和24 h的植株有所降低,但高温胁迫处理48 h的植株在恢复生长12 h后其MDA含量才从处理后的0.034 μmol/g急升至0.094 μmol/g,随后才回落恢复到与45℃胁迫处理24 h的植株相当的含量,说明45℃极端高温处理后会引起MDA的大量积累,并有一定的延后性。
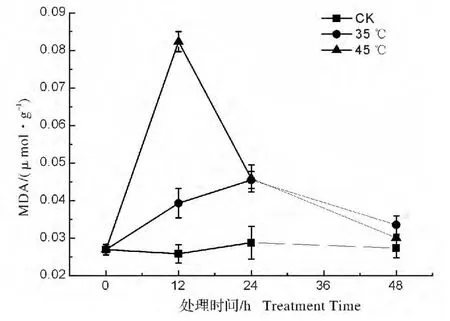
图5 高温胁迫处理对弗吉尼亚栎叶片MDA含量的影响Fig.5 Effect of heat stress on MDA content in Quercus virginiana leaves

图6 不同处理阶段弗吉尼亚栎叶片中MDA含量的变化Fig.6 Change of MDA in Quercus virginiana leaves at different treatment phase
2.4 高温胁迫及恢复生长对弗吉尼亚栎幼苗脯氨酸含量的影响
在高温处理48 h内,弗吉尼亚栎植株中的脯氨酸含量均表现出先升高后降低的趋势,并且脯氨酸含量的最高点均出现在处理24 h后,但45℃胁迫下植株中的脯氨酸含量和增幅均显著高于35℃处理(图7)。这表明45℃处理对弗吉尼亚栎幼苗叶片伤害更大。
35℃胁迫处理后的植株中脯氨酸的含量在恢复生长过程中迅速降低(图8A),其变化趋势基本与MDA含量的变化一致。而45℃胁迫处理12 h和24 h的植株中的脯氨酸含量在恢复生长的12 h仍在增加,在恢复生长72 h之后植株中的脯氨酸含量都基本上恢复到了处理前水平(图8B)。

图7高温胁迫处理对弗吉尼亚栎叶片脯氨酸含量的影响Fig.7 Effect of heat stress on proline content in Quercus virginiana leaves
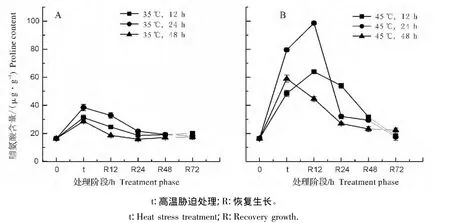
图8 不同处理阶段弗吉尼亚栎叶片中脯氨酸含量的变化Fig.8 Change of proline content in Quercus virginiana leaves at different treatment phase
3 讨论与结论
植物体中的各种代谢活动都需要水分的参与,在不同环境的胁迫下植物体内的水分均会受到不同程度的影响,植物会出现脱水现象[9]。植物在逆境胁迫下的水分含量变化在一定程度上也反应了植物体的生存状态和抗逆性能。弗吉尼亚栎幼苗的叶片含水量随着胁迫处理时间的延长逐渐下降,但总体变化幅度较小。45℃胁迫下,幼嫩叶片边缘出现了一定程度的脱水变焦。含水量在胁迫处理后较短时间内可基本恢复至处理前水平(图2)。
细胞膜系统是植物在逆境胁迫条件下最主要的受害部位。植物在受到高温胁迫时,叶表温度上升,可能导致抗氧化酶活性降低[10],自由基大量积累,进而导致膜脂过氧化水平增高,MDA含量增加,膜脂成分改变,不饱和指数下降,膜透性增大,相对电导率升高,膜结构和功能改变,从而引起一系列生理代谢变化[11]。弗吉尼亚栎叶片的电解质外渗率在高温胁迫12 h后即出现了不可逆的显著上升,但并没有随胁迫时间延长而持续增加(图4),暗示了弗吉尼亚栎幼苗叶片细胞膜可能通过一些自身的其他生理生化特征调节形成了较良好的热稳定性,从而防止了长时间的高温胁迫对细胞膜的进一步破坏。45℃高温处理组的弗吉尼亚栎幼苗叶片中MDA含量和脯氨酸含量增加要显著大于35℃处理组(图5和图7),表明了胁迫温度越高,膜脂过氧化水平越高,植株受胁迫的程度也相应加剧。但同时,我们也发现随着胁迫时间的增加,在胁迫处理前中期(12~24 h),MDA和脯氨酸含量基本呈现增长趋势,但到48 h后均出现了一定程度降低,暗示植株可能通过自身生理代谢变化对高温胁迫形成了一定的抗逆性,植株受胁迫程度得以缓和。经胁迫处理后的植株经过72 h的恢复生长后,除细胞液渗漏不可逆外,叶片水分可以有效恢复,MDA和脯氨酸含量均降低到接近处理前水平,进一步说明了弗吉尼亚栎对高温具有良好的适应性。此外,我们对不同实验处理后的弗吉尼亚栎幼苗在常规温室下进行了2个月的栽培观察,植株均没有出现死亡,且基本恢复了正常生长,说明高温胁迫处理并没有对其造成致命伤害。
耐热性好的植物经过长期的历史演化发展出了一系列形态结构、生理及生化特征,通过规避伤害和提高耐性两大机制来实现自身在高温逆境下的生存[6]。首先,弗吉尼亚栎幼苗叶片较小,受空气边界层的阻力也小于大叶植物,因而在水分充足情况下面临极端高温时,通过加大蒸腾作用,更容易向周围环境散发自身热量;再则弗吉尼亚栎的叶片表面具有较厚的蜡质和表皮毛[12],可以减少高温辐射造成的伤害[10],从而为其应对逆境提供了较好的保护。
本研究通过对弗吉尼亚栎幼苗在高温胁迫下和恢复生长过程中叶片水分以及与细胞膜稳定性相关生理指标的测定,证实了弗吉尼亚栎能通过自身的耐热机制抵御高温胁迫对其造成的伤害。弗吉尼亚栎对高温胁迫有良好耐受性和适应性的探明,是其除抗风[2]、耐盐[3,4]、耐旱[5]等优良特性外的又一重要优良生理特性,这将为弗吉尼亚栎作为一种优良的园林和防护林树种的进一步推广应用提供有力的理论支撑。
[1]Bartens J,Grissino-Mayer H D,Day S D,et al.Evaluating the potential for dendrochronological analysis of live oak(Quercus virginiana Mill.)from the urban and rural environment—An explorative study[J].Dendrochronologia,2012,3(1):15-21.
[2]陈益泰,陈雨春,黄一青,等.抗风耐盐常绿树种弗吉尼亚栎引种初步研究[J].林业科学研究,2007,20(4):542-546.
[3]王树凤,陈益泰,孙海菁,等.盐胁迫下弗吉尼亚栎生长和生理生化变化[J].生态环境,2008,17(2):747-750.
[4]王树凤,胡韵雪,李志兰,等.盐胁迫对弗吉尼亚栎生长及矿质离子吸收、运输和分配的影响[J].生态学报,2010,30(17):4609-4616.
[5]王树凤,孙海菁,陈益泰,等.模拟干旱胁迫下弗吉尼亚栎苗木叶片相关生理参数的分析[J].南京林业大学学报:自然科学版,2011,35(6):6-10.
[6]Bita C E,Gerats T.Plant tolerance to high temperature in a changing environment:Scientific fundamentals and production of heat stress-tolerant crops[J].Front Plant Sci,2013,4:273.
[7]孙国胜,邓敏,李谦盛,等.牛耳朵幼苗对低温胁迫的生理响应[J].南方农业学报,2013,44(6):918-923.
[8]李玲.植物生理学模块实验指导[M].北京:科学出版社,2010.
[9]Levitt A D.Responses of plant to environmental stresses[M].New York:Academic Press,1990.
[10]Hasanuzzaman M,Nahar K,Alam M M,et al.Physiological,biochemical,and molecular mechanisms of heat stress tolerance in plants[J].Int J Mol Sci,2013,14(5):9643-9684.
[11]张庆峰,徐胜,李建龙.高温胁迫下高羊茅生理生化特性研究[J].草业科学,2006,23(4):26-28.
[12]Mittler R.Abiotic stress,the field environment and stress combination[J].Trends Plant Sci,2006,11(1):15-19.
[13]Hardin J W.Terminology and classification of Quercus trichomes[J].Journal of the Elisha Mitchell Scientific Society,1976,92(4):151-161.
猜你喜欢
中老年保健(2022年3期)2022-11-21
英美文学研究论丛(2021年2期)2021-02-16
百科探秘·海底世界(2020年8期)2020-07-29
世界科学(2020年1期)2020-02-11
科学与财富(2019年15期)2019-10-21
中国生物医学工程学报(2019年5期)2019-07-16
发酵科技通讯(2018年2期)2018-07-06
Coco薇(2017年5期)2017-06-05
湖南林业科技(2017年1期)2017-02-06
现代电生理学杂志(2015年4期)2015-07-18
