
干旱胁迫对6 种藤本植物光合作用及叶绿素荧光参数的影响
2015-07-13 02:06邓沛怡周杰良陶抵辉刘坤吴易雄
湖南农业大学学报(自然科学版) 2015年3期
邓沛怡,周杰良,陶抵辉,刘坤,吴易雄,2*
(1.湖南生物机电职业技术学院藤本植物研究所,湖南 长沙410127;2.湖南广播电视大学,湖南 长沙 410004)
近年来,随着全球气温的回升,中国南方大部分地区在盛夏都出现了高温干旱天气,这对植物的生长产生了严重影响。藤本植物是园林绿化的重要组成部分,其种类繁多,生态适应性强,生长迅速,在绿化引种及边坡生态防护中有独特的优势[1–4]。中国南方地区应用较多的藤本植物有:扶芳藤(Euonymus fortunei),为常绿蔓性灌木,生长旺盛,是庭院绿化的主要植物;金银花(Lonicera japonica),为半常绿木质藤本植物,根系繁密发达,萌蘖性强,茎蔓着地即能生根;绿爬山虎(Parthenocissus laetevirens),为葡萄科地锦属植物,攀援于其他植物上,生长快,攀援力极强;络石(Trachelospermum jasminoides),为木质藤本植物,常攀援于树上、墙壁或岩石上,在园林中多作地被;常春油麻藤(Mucuna sempervirens),为豆科油麻藤属常绿藤本植物,适于攀附建筑物、陡坡、岩壁等处生长,是良好的垂直绿化植物[5];常春藤(Hedera nepalensis),为五加科常绿木质藤本植物,在庭院中可用以攀援假山、岩石或在建筑阴面做垂直绿化材料[6]等。目前,叶绿素荧光动力学检测技术已被广泛应用于各种胁迫对植物生理生化影响的研究[7–8]中。有关干旱胁迫对扶芳藤、常春油麻藤等藤本植物生理生化指标影响的研究[1,6]已有报道,而干旱胁迫对扶芳藤、金银花、绿爬山虎等藤本植物的光合特性及叶绿素荧光参数影响的研究尚少。笔者对此进行了研究,现将结果报道如下。
1 材料与方法
1.1 供试材料
扶芳藤(Euonymus fortunei (Turcz.) Hand.–Mazz.)、金银花(Lonicera japonica Thunb.)、绿爬山虎(Parthenocissus laetevirens Rehd.) 、 络 石(Trachelospermum jasminoides (Lindl.) Lem.)、常春油麻藤(Mucuna sempervirens Hemsl.)、常春藤(Hedera nepalensis K. Koch var. sinensis (Tobl.) Rehd.) 6 种藤本植物均为湖南生物机电职业技术学院藤本植物研究所1年生扦插苗。
1.2 试验设计
试验于2014年6—9月在湖南生物机电职业技术学院内进行。选取长势一致的扦插苗,移栽到规格为15 cm×15 cm×12 cm 装有基质的塑料盆中。基质的田间持水量为26.3%,其中含有机质11.1g/kg,全氮0.5g/kg,全磷3.7mg/kg,速效钾100.2mg/kg。干旱胁迫设4个水分处理:正常供水(CK)、轻度干旱(LS)、中度干旱(MS)、重度干旱(SS),其土壤的含水量分别为田间持水量的90%~85%、75%~70%、55%~50%、35%~30%。用称重法控制不同处理土壤水分恒定。播种后每天于18:00 称重,并及时补充失去的水分,使各处理保持设定的土壤含水量[9]。土壤田间持水量=(当天盆重-土壤干重)/ (浇透水盆重-土壤干重)。土壤在85℃下烘干后装盆,称重即为土壤干重;在下午浇透水过夜,使多余的水流掉,第二天早上称重即为浇透水盆重。根据设定的田间持水量可以推算出当天盆重。为了减少水分蒸发,在盆口套塑料袋[10]。每个处理9 盆,每种植物36 盆,共计216 盆。
1.3 测定项目与方法
干旱处理1个月后依次测定藤本植物光合参数、叶绿素SPAD 值和叶绿素荧光参数。
1) 光合特性指标的测定。2014年8月上旬连续3 d 的8:30—11:00 选取植株倒3 叶,用Li–6400xt便携式光合仪(Li–COR USA)测定叶片的净光合速率(Pn)、气孔导度(Gs)、细胞间CO2浓度(Ci)及蒸腾速率(Tr)。水分利用效率(WUE)=净光合速率/蒸腾速率,测定时用红蓝光源,光强设为1 000 μmol/(m2·s),CO2气体由小钢瓶提供,控制浓度为400 μmol/(m2·s) (下同)。每个处理测定3 株。重复3次,结果取平均值。
2) 光合–光响应特征参数的测定。光合有效辐射梯度设定为2 400、2 100、1 800、1 500、 1 200、900、600、200、150、100、75、50、25、0 μmol/(m2·s),用6400–LED 仪测定光合–光响应特征参数。光饱和点(LSP)、光补偿点(LCP)等光合指标根据叶子飘等[11]的直角双曲线修正模型进行拟合计算。每个处理测定3 株。每株测定3 片叶。重复3次,结果取平均值。
3) 相对叶绿素含量的测定。采用SPAD–502 叶绿素测定仪(日本Konica 公司产品),在测定光合作用的同时测定叶绿素相对含量。测定时,在每个叶片主脉两侧中部从叶尖到叶基部均等取9个点测量。每个处理测定3 株。每株测定3 片叶。重复3次,结果取平均值。
4) 叶绿素荧光参数的测定。采用Li–6400xt 便携式光合仪(Li–COR,USA),在凌晨破晓前,按照操作手册对经过充分暗适应的叶片照射检测光,测定Fo(初始荧光),然后在施加饱和脉冲为7 200 μmol/(m2·s)的光强下0.8 s 测得暗适应下最大荧光Fm;于当天8:00 用叶室内活化光将叶片活化30min以上测定光化学量子产量(Y)和电子传递速率(ETR)等参数。荧光参数的计算参照Roháček 的方法[12]。每个处理测定3 株。每株测定3 片叶。重复3次,结果取平均值。
1.5 数据统计与分析
用Excel 2007 处理数据;用SPSS 17.0 软件进行方差分析。
2 结果与分析
2.1 干旱对6 种藤本植物相对叶绿素含量(SPAD值)的影响
由图1 可知,在CK 和LS 下,常春藤SPAD值高于其他植物,且差异显著(P<0.05);在MS和 SS下,常春藤SPAD 值低于扶芳藤及络石,但高于其他植物;在所有处理中,绿爬山虎的SPAD 值均低于其他植物,且差异达到显著水平 (P<0.05)。在 MS 下,与CK 相比,扶芳藤、金银花、绿爬山虎、络石、常春油麻藤、常春藤的SPAD 值分别降低了8.58 %、7.48%、9.41%、7.70%、10.61%、19.21%;在SS 下,SPAD 值分别降低了10.25%、17.04%、16.58%、23.28%、25.35%、30.33%,说明中度及重度干旱均对常春藤SPAD 值影响较大,重度干旱对扶芳藤SPAD 值影响较小。
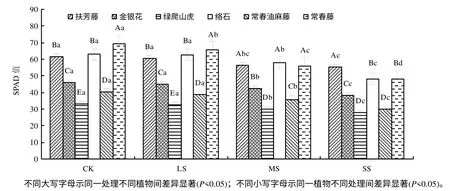
图1 干旱胁迫下6 种藤本植物的SPAD 值 Fig.1 Relative chlorophyll content of six kinds of lianas in drought stress
2.2 干旱对6 种藤本植物光合参数的影响
2.2.1 对净光合速率的影响
图2 表明,在CK、LS、MS 处理下,金银花叶片的Pn最大;在SS 下,金银花的Pn高于常春藤叶片的Pn,但低于其他4 种植物叶片的Pn。在4种处理下,常春藤叶片的Pn与其他5 种植物比较差异达到显著水平 (P<0.05)。在MS 下,与CK 相比,扶芳藤、金银花、绿爬山虎、络石、常春油麻藤、常春藤的Pn分别降低了25.75%、35.58%、26.08%、22.28%、26.59%、24.18%,且差异达到显著水平 (P<0.05);在SS 下,与CK 相比,扶芳藤、金银花、绿爬山虎、络石、常春油麻藤、常春藤的Pn分别降低了59.93%、77.20%、65.16%、57.35%、56.04%、51.26%,且差异均达到极显著水平(P<0.01),说明在SS 下对金银花和绿爬山虎叶片的Pn影响较大,Pn降幅由小到大的6 种藤本植物顺序为常春藤、常春油麻藤、络石、扶芳藤、绿爬山虎、金银花。
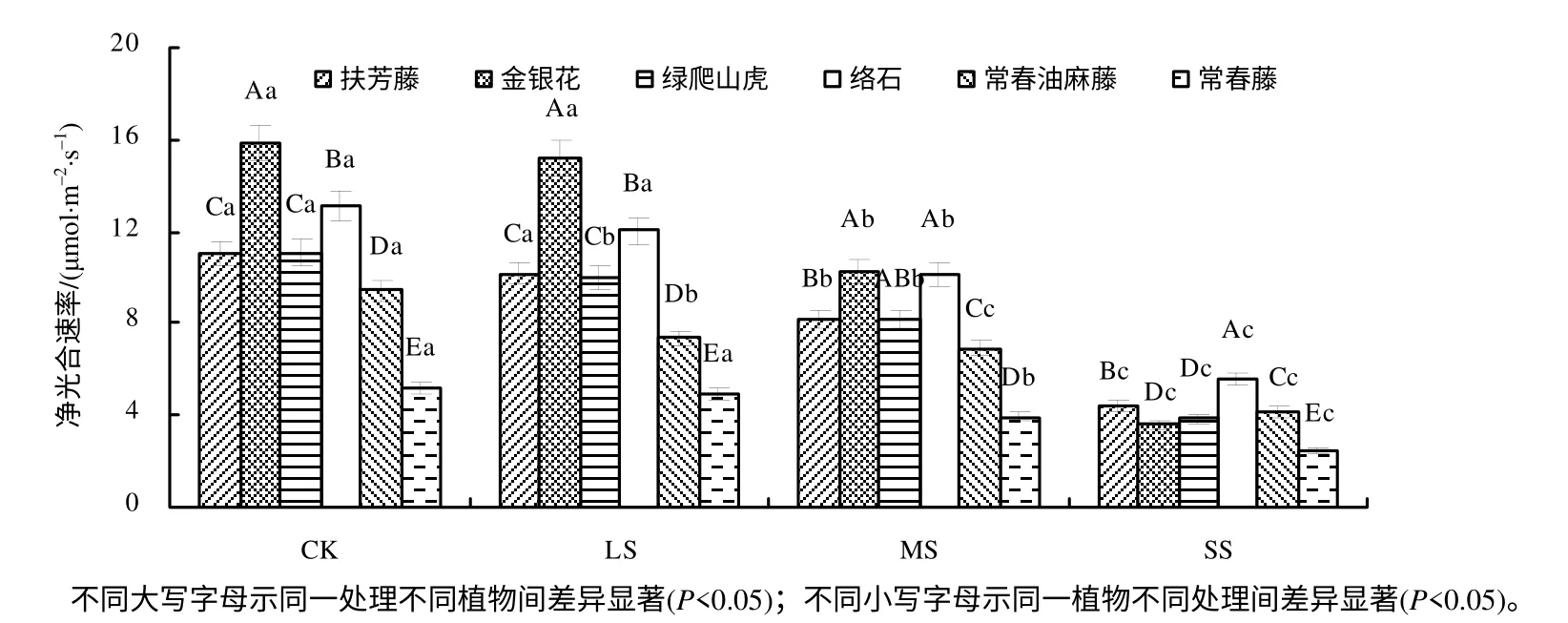
图2 干旱胁迫下6 种藤本植物的净光合速率 Fig.2 Net photosynthetic rates of six kinds of lianas in drought stres s
2.2.2 对气孔导度的影响
从图3 可以看出,Gs的整体变化趋势与Pn的一致,在CK、LS、MS 下,6 种植物中金银花的Gs最大,在 SS 下,金银花的 Gs较小,为 0.03 mmol/(m2·s),说明重度干旱对金银花叶片的Gs影响较大;常春藤叶片的Gs均低于其他藤本植物叶片的Gs。在SS 下,与CK 相比,扶芳藤、金银花、绿爬山虎、络石、常春油麻藤、常春藤的Gs分别降低了80.95 %、94.12%、73.33%、86.96%、85.00%、87.50%,且差异达到极显著水平(P<0.01),其中,绿爬山虎的Gs降幅最小,金银花的Gs下降幅度最大,其他4 种植物的Gs降幅介于绿爬山虎与金银花之间。
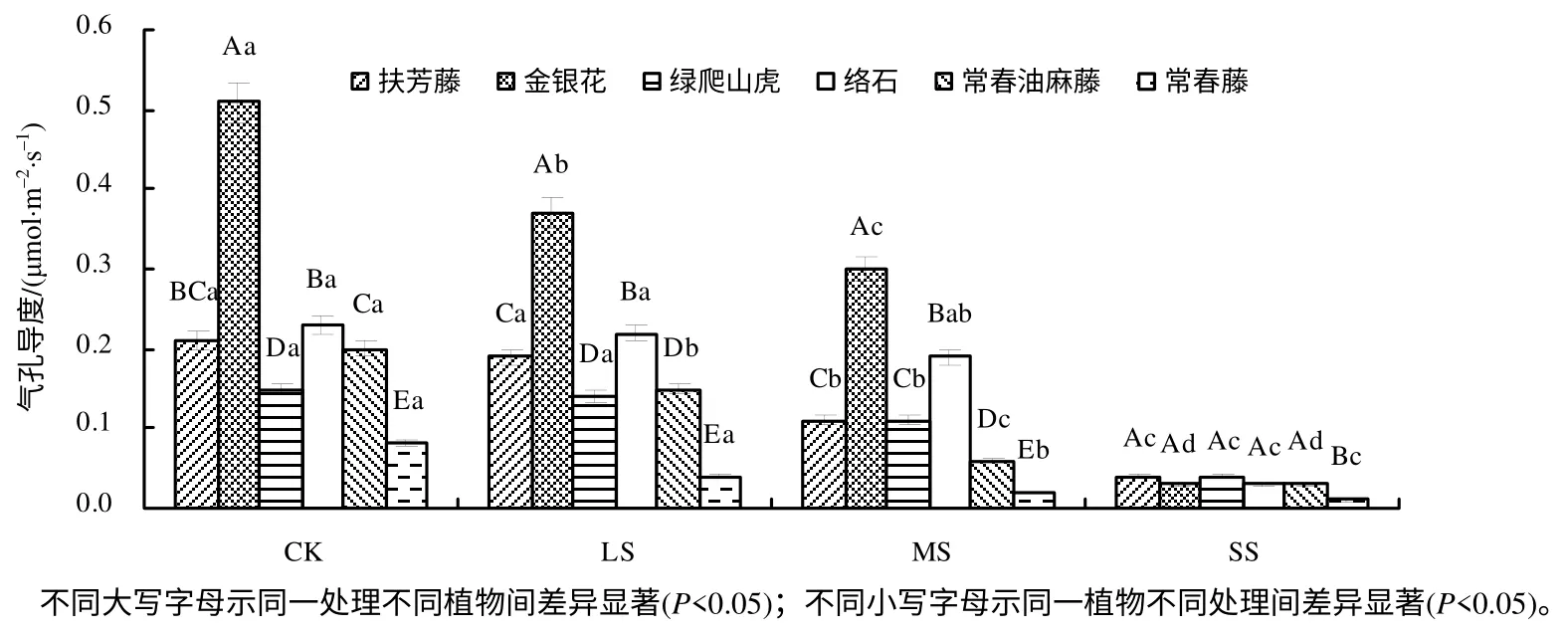
图3 干旱胁迫下6 种藤本植物的气孔导度 Fig.3 Stomatal conductances of six kinds of lianas in drought stress
2.2.3 对蒸腾速率的影响

图4 干旱胁迫下6 种藤本植物的蒸腾速率 Fig.4 Transpiration rates of six kinds of lianas in drought stress
由图4 可知,Tr的变化趋势与Pn和Gs的基本一致,在CK、LS 及MS 下,金银花的Tr均高于其他5 种藤本植物的Tr;在SS 下,扶芳藤、金银花、 绿爬山虎、络石、常春油麻藤、常春藤的Tr分别为1.16、0.87、1.01、0.70、0.78、0.42 mmol/(m2·s),说明在重度干旱条件下,扶芳藤和绿爬山虎的Tr较高,常春藤的Tr最低。
2.2.4 对胞间CO2浓度的影响
由图5 可知,Ci的变化与Pn、Gs及Tr的变化无明显相关性,在4 种处理下,常春藤的Ci均低于其他植物的Ci;在CK、LS、MS 下,金银花、绿爬山虎、络石、常春藤的Ci差异没有统计学意义(P>0.05),说明MS 及LS 对Ci影响较小。与CK 相比,在SS 下,扶芳藤、金银花、绿爬山虎、络石、常春油麻藤、常春藤的Ci分别降低了22.02%、43.69%、37.30%、53.21%、49.48%、12.99%,差异达到显著水平(P<0.05)。
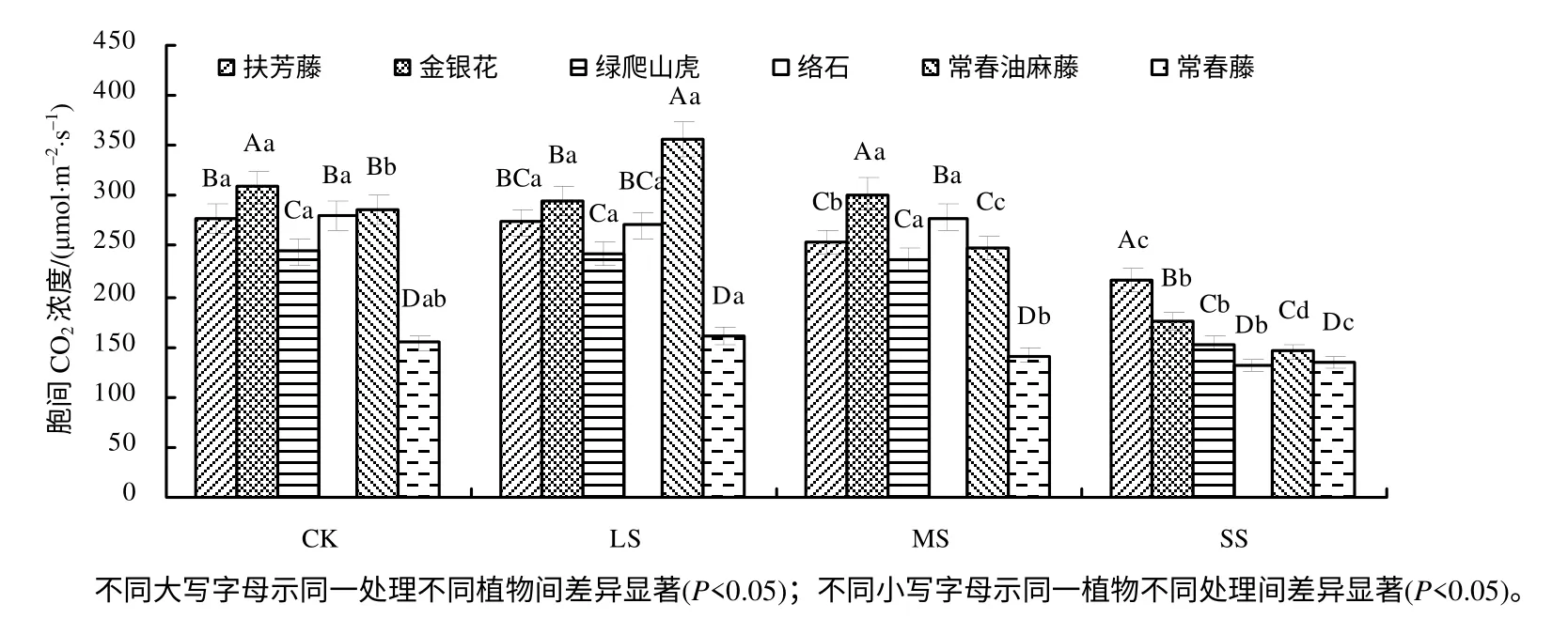
图5 干旱胁迫下6 种藤本植物的胞间CO2 浓度 Fig.5 Intercellular CO2 concentration of six kinds of lianas in drought stress
2.2.5 对水分利用率的影响
由图6 可知,随着干旱胁迫的加重,6 种藤本植物的WUE 呈逐渐升高趋势,说明干旱胁迫有利于提高植物的水分利用效率;在CK、LS、MS 下,常春藤的WUE 最高,分别为4.61、4.65、5.03 μmol/(m2·s),在SS 处理下,WUE 由高到低的6 种藤本植物的顺序为络石、常春藤、常春油麻藤、扶芳藤、金银花、绿爬山虎。

图6 干旱胁迫下6 种藤本植物的水分利用率 Fig.6 Water use efficiency of six kinds of lianas in drought stress
2.3 干旱对6 种藤本植物光合–光响应特征参数的影响
由表1 可知,6 种藤本植物最大净光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)及暗呼吸速率(Rd)的值随着干旱胁迫的加重整体降低。金银花叶片的Pmax均值最高,为12.45 μmol/(m2·s);常春藤叶片的Pmax均值最低,为4.49 μmol/(m2·s);扶芳藤、绿爬山虎、络石、常春油麻藤的Pmax均值分别为9.25、9.21、11.21、7.52 μmol/(m2·s)。在正常供水(CK)下,扶芳藤的LSP 最高,为1 973 μmol/(m2·s);常春藤的最低,为1 509 μmol/(m2·s)。在SS 下,络石的LSP 最高,为703 μmol/(m2·s);常春藤的LSP最低,为497 μmol/(m2·s)。光补偿点(LCP)的高低说明植物对弱光的利用能力。金银花和络石的LCP 均值较高,分别为15.20、18.09 μmol/(m2·s);绿爬山虎的LCP 均值最低,为6.36 μmol/(m2·s)。表观量子效率(AQY)是植物叶片光能利用效率的一个重要指标,它的大小反映叶片对弱光的利用能力[13]。金银花和络石的AQY 较低,而绿爬山虎的AQY 较高,说明金银花和络石适合在阳光较充足的环境下生长,而绿爬山虎较适合在阴湿的环境中生长。6 种藤本植物的暗呼吸速率(Rd)与Pmax呈正相关关系。

表1 干旱胁迫下6 种藤本植物的光合–光响应特征参数 Table 1 Effects of drought stress on photosynthetic-light response characteristics parameters of six kinds of lianas
2.4 干旱对6 种藤本植物叶绿素荧光参数的影响
由表2 可以看出,干旱胁迫使6 种藤本植物的Fo增加,最大荧光(Fm)降低。在正常供水(CK)下,6种藤本植物的最大光化学效率(Fv/Fm)都在0.80 以上;在SS 下,金银花、绿爬山虎、常春藤的Fv/Fm降低到0.75 以下,而络石、常春油麻藤的Fv/Fm都在0.76 以上,说明金银花、常春油麻藤及常春藤受到干旱胁迫的程度高于其他3 种藤本植物。6 种藤本植物的光化学量子产量(Y)、电子传递速率(ETR)及光化学猝灭系数(qp)随着干旱胁迫的加重而降低。在SS下,与CK 相比,扶芳藤、金银花、绿爬山虎、络石、常春油麻藤、常春藤的ETR 分别降低了27.06%、39.52%、41.84%、23.47%、25.66%、40.41%,说明干旱胁迫对绿爬山虎的ETR 影响最大。

表2 干旱胁迫下6 种藤本植物的叶绿素荧光参数 Table 2 Effects of drought stress on chlorophyll fluorescence parameters of six kinds of lianas
3 结论与讨论
土壤水分胁迫对植物生长和代谢的影响是多方面的,其中对光合作用的影响尤为突出[14]。水分胁迫通常抑制叶绿素的生物合成,提高叶绿素酶活性并加速其分解,致使叶绿素含量下降[15],本研究结果与之一致。叶绿体是植物进行光合作用的主要场所,干旱胁迫降低了叶绿素含量,即叶绿素SPAD值,从而使得光合酶活性和光合作用表光量子效率等下降,最终导致Pn降低,这主要是由非气孔因素引起的[16]。影响光合作用的因素除了非气孔因素外,还有气孔因素[17]。通常情况下,植物Pn下降主要是由于植物气孔阻力升高,即由气孔导度降低引起的。 Gs降低是否是光合作用下降的原因,既要看Gs的大小,还要看Ci的变化[18]。如果Gs降低,Ci也降低,则Pn的下降主要是由气孔因素引起的;相反,则是非气孔因素引起的。本研究结果表明,在LS 和MS 下,6 种藤本植物的Pn和Gs稍有降低,而Ci并未降低,说明此时Pn降低主要是由非气孔因素引起的;在SS 下,6 种藤本植物的Pn、气孔导度及Ci均降低,说明此时Pn的降低主要是由气孔因素引起的。
通过对6 种藤本植物光合生理指标的比较发现,在正常供水下,金银花的光合效率要高于其他5 种藤本植物,而常春藤叶片的Pn、Gs及Tr在本试验设置的水分胁迫下均低于其他5 种藤本植物。WUE 是植物抗旱的重要指标之一,虽然金银花具有较高的Pn,但WUE 低于常春藤的,说明常春藤在长期自然条件下已经形成了利用水分的特殊机制,在干旱胁迫下的表现更为明显。随着干旱胁迫的加重,6 种藤本植物的WUE 逐渐升高,植物通过降低Gs,从而减少蒸腾过程中水分的散失,保持较高的节水能力,这与尹丽等[19]的研究结果一致。在重度干旱下,络石具有较高的Pn及WUE,说明络石是干旱环境下抗旱的先锋植物之一。
在光响应曲线拟合过程中,大部分研究人员采用直角双曲线模型与非直角双曲线模型,不同模型拟合出的结果相差较大。笔者在实际应用过程中发现,用这些模型拟合光响应数据时,所给出的最大净光合速率远大于实测值[20],而所给出的光饱和点远小于实测值[21]。由于这些模型是1 条渐进曲线,不存在极点,故无法由这些模型直接给出植物的光饱和点和最大净光合速率的解析解。在6 种植物的光合曲线拟合过程中,采用非直角双曲线模型拟合出的光饱和点较低,最大净光合速率较高,与实测值相差甚远。本研究中采用的是叶子飘等[11]的直角双曲线修正模型,拟合方法简单,效果理想。
对6 种藤本植物的光合生理指标及叶绿素荧光参数的综合评价表明,在正常供水及轻度干旱下,光合效率从高到低的植物依次为金银花、络石、扶芳藤、绿爬山虎、常春油麻藤、常春藤;在重度干旱胁迫下,抗旱性由强到弱的植物依次为络石、常春油麻藤、扶芳藤、常春藤、金银花、绿爬山虎。
[1] 陈柯,王小德.常春油麻藤等3 种藤本植物抗旱性生理指标研究[J].浙江林学院学报,2008,25(3):314–318.
[2] 刘伟玲,谢双喜,喻理飞.几种喀斯特森林树种幼苗对水分胁迫的生理响应[J].贵州科学,2003,1(3):51–55.
[3] 徐利霞.石漠化山区造林树种幼苗的抗旱性研究[D].重庆:西南大学,2007.
[4] 王元军.公路边坡绿化中藤本植物的选择[J].特种经济动植物,2005(7):32.
[5] 韦直,何业祺.浙江植物志[M].第3 卷.杭州:浙江科学技术出版社,1993:371–372.
[6] 张朝阳,许桂芳,向佐湘.干旱胁迫对4 种常绿藤本植物抗性生理生化指标的影响[J].江西农业学报,2008,20(12):42–45.
[7] Kocurek V , Smutny V , Filova J . Chlorophyll fluorescence as an instrument for the assessment of herbicide efficacy[J].Cereal Res Commun,2009,37(1):289–292.
[8] Kopsell D A, Armel G R,Abney K R, et al.Leaf tissue pigments and chlorophyll fluorescence parameters vary among sweet corngenotypes of differential herbicide sensitivity [J].Pest Biochem Physiol,2011,99:194–199.
[9] 王智威,牟思维,闫丽丽,等.水分胁迫对春播玉米苗期生长及其生理生化特性的影响[J].西北植物学报,2013,33(2):343–352.
[10] Mauro Centritto1,Federico Brilli,Roberta Fodale, et al. Different sensitivity of isoprene emission,respiration and photosynthesis to highgrowth temperature coupled with drought stress in black poplar (Populus nigra) saplings [J].Tree Physiology,2011,31:275–286.
[11] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356–1361.
[12] Roháček K . Chlorophyll fluorescenceparameters :Thedefinitions photosynthetic meaning and mutual relationships[J].Photosynthetica,2002,40(1):13–29.
[13] 朱巧玲,冷佳奕,叶庆生.黑毛石斛和长距石斛的光合特性[J].植物学报,2013,48(2):151–159.
[14] 付秋实,李红岭,崔健,等.水分胁迫对辣椒光合作用及相关生理特性的影响[J].中国农业科学,2009,42(5):1859–1866.
[15] 李清芳,马成仓,尚启亮.干旱胁迫下硅对玉米光合作用和保护酶的影响[J].应用生态学报,2007,18(3):531–536.
[16] Yang X H,Chen X Y,Ge Q Y,et al.Tolerance of photosynthesis to photoinhibition,high temperature and drought stress in flag leaves of wheat:A comparison between a hybridization line and its parentsgrown under field conditions[J].Plant Science,2006,171:389–397.
[17] 安玉艳,梁宗锁.植物应对干旱胁迫的阶段性策略[J].应用生态学报,2012,23(10):2907–2915.
[18] Farquhar G D,Sharkey T D,Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317–345.
[19] 尹丽,胡庭兴,刘永安,等.干旱胁迫对不同施氮水平麻疯树幼苗光合特性及生长的影响[J].应用生态学报,2010,21(3):569–576.
[20] Kyei-Boahen S,Lada R,Astatkie T,et al.Photosynthetic response of carrots to varying irradiances[J]. Photo- synthetica,2003,41:1–5.
[21] 高峻,孟平,吴斌,等.杏–丹参林药复合系统中丹参光合和蒸腾特性的研究[J].北京林业大学学报,2006,28(2):64–67.
猜你喜欢
新作文·小学高年级版(2022年2期)2022-03-31
阅读(中年级)(2020年11期)2020-12-28
当代作家(2018年11期)2018-11-27
中成药(2018年8期)2018-08-29
现代园艺(2018年3期)2018-02-10
文艺生活·上旬刊(2017年12期)2018-01-10
作文周刊·小学三年级版(2016年20期)2017-05-26
肝博士(2016年3期)2016-11-17
新高考·高二数学(2016年2期)2016-03-07
西江月(2014年4期)2014-03-13
