
山桃醇腈酶基因PdHNL1的克隆、序列分析与原核表达
2015-08-20 20:58傅江燕徐晟夏冰汪仁
江苏农业科学 2015年7期
傅江燕+徐晟+夏冰+汪仁


摘要:以山桃叶片为材料提取RNA,反转录后得到cDNA,通过PCR扩增得到1个山桃醇腈酶基因全长编码序列,并命名为[WTBX][STBX]PdHNL1[WTBZ][STBZ]。[WTBX][STBX]PdHNL1[WTBZ][STBZ]的开放阅读框为1 737 bp,预测编码蛋白包含539个氨基酸。氨基酸序列比对结果表明,[WTBX][STBX]PdHNL1[WTBZ][STBZ]编码的蛋白具有葡萄糖甲醇胆碱氧化还原酶的序列特征,与黑樱桃醇腈酶PsHNL4蛋白相似度为92%,与桃假定的HNL蛋白相似度为77%。通过将[WTBX][STBX]PdHNL1[WTBZ][STBZ]基因连接到原核表达载体pET28(a)上,并转化大肠杆菌BL21 (DE3)后,成功构建了原核表达重组菌株。SDS-PAGE电泳结果显示,重组蛋白受IPTG诱导表达,分子量大小约为62.4 ku,与已报道的其他蔷薇科植物的醇腈酶蛋白大小基本一致。
关键词:山桃;醇腈酶;基因克隆;原核表达
中图分类号:S662.101;Q786 文献标志码: A 文章编号:1002-1302(2015)07-0020-05
利用酶作为催化剂用于合成单一对映体或直接分离特定手性异构体,被广泛应用于不对称合成领域。醇腈酶作为一种不对称催化手性反应的酶,自20世纪初在扁桃中发现以来,受到愈来愈多的关注[1]。它可逆地催化HCN和醛/酮类化合物反应生成手性氰醇[2],进而转化成为羟基化合物等多种手性中间体,并用于合成多种手性药物,从而克服了为得到纯手性化合物而采用光学拆分所带来的生产工序复杂、成本高等一系列问题[3-4]。此外,在植物体内,醇腈酶可催化氰基糖苷类化合物,反应生成羰基化合物和HCN,而后者在植物防御草食类动物摄食或其他致病菌感染中起到作用,即植物的生氰作用[5-6]。
HNL在自然界中是广泛存在的。据统计,有3 000~12 000 种植物内含有HNL。HNL主要分为两大类:R构型和S构型。目前,已分离纯化并进行酶活分析的R型HNL多来自于蔷薇科植物[7],如扁桃、黑樱桃、梅、枇杷、西番莲等[8-13];而S型HNL多来自于木薯、橡胶树、海檀木与高粱[14-17]。笔者所在研究组根据前期公布的桃的基因组数据[18],通过序列同源比对分析,发现其中有3条序列为编码推测的HNL蛋白。此外,山桃、桃与扁桃同属于蔷薇科桃属,目前有关山桃中HNL的研究并未报道。因此,本研究采用生物信息学结合PCR的方法,克隆得到1个山桃HNL基因,并对其进行了相关的生物信息学分析与原核表达研究。
1 材料与方法
1.1 材料与试剂
山桃[Prunus davidiana (Carr.) C.]叶片取自江苏省中国科学院植物研究所,采集后迅速放入液氮中冷冻,于 -80 ℃ 保存备用。
高纯总RNA快速提取试剂盒购自北京百泰克生物技术有限公司,表达载体pET28(a)购自Novagen公司,rTaq DNA聚合酶、克隆载体pMD19-T、Oligo (dT)18、DNA Markers、限制性内切酶和反转录酶M-MLV (RNase H—)等购自大连宝生物工程有限公司;胰蛋白胨、酵母提取物购自Oxoid公司;其他化学试剂均为国产分析纯。大肠杆菌DH5α、TOP10和BL21 (DE3)菌株为笔者所在实验室保存,引物合成和测序工作分别委托北京鼎国昌盛生物技术有限责任公司与上海美吉生物医药科技有限公司完成。
1.2 方法
1.2.1 总RNA提取及cDNA的合成 按照高纯总RNA快速提取试剂盒的操作说明,提取山桃叶片的RNA,通过琼脂糖凝胶电泳检测RNA纯度与完整性,并以此为模板,根据TaKaRa公司的反转录试剂盒说明书反转录得到山桃cDNA。
1.2.2 [WTBX][STBX]PdHNL1[WTBZ][STBZ]基因的克隆及生物信息学分析 根据桃基因组测序组公布的序列,通过比对搜索出编码假定醇腈酶蛋白的基因序列,并根据这些序列设计特异引物,上游引物为 F0:5′-ATGGTGAAATCAACAATGTC-3′,下游引物为 R0:5′-AGTATCAAACGCAAAGGAT-3′。以反转录得到的 cDNA 为模板进行 PCR扩增,扩增反应程序为:94 ℃预变性 5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸50 s,35个循环;72 ℃ 延伸10 min。PCR产物经 1%琼脂糖凝胶电泳后切胶回收,纯化后的产物连接到 pMD19T载体上,转入大肠杆菌 DH5α感受态细胞,挑取克隆进行PCR与质粒酶切检测,鉴定结果均为阳性的重组子送交公司测序。
测序结果通过NCBI的ORF finder在线软件查找该序列的开放阅读框,得到其编码的氨基酸序列,将该序列使用BlastP进行比对,选择与PdHNL同源的其他植物的醇腈酶蛋白序列,进一步利用DNAman软件进行多重序列比对并使用MEGA5构建系统进化树;使用ExPASy的Compute pI/MW计算蛋白的等电点与分子量;用TMHMM预测跨膜结构;用SignalP 4.1预测信号肽;用PROSITE数据库进行功能结构预测;用SWISS-MODEL预测三级结构并建模。
1.2.3 [WTBX][STBX]PdHNL1[WTBZ][STBZ]基因的载体构建与原核表达 根据获得的[WTBX][STBX]PdHNL1[WTBZ][STBZ]基因与pET28(a)载体图谱,分析并设计带有酶切位点EcoRⅠ和NotⅠ的引物PdHNL1-pETF和PdHNL1-pETR (表1),使用rTaq DNA聚合酶进行PCR扩增。
将pMD19T-[WTBX][STBX]PdHNL1[WTBZ][STBZ]重组质粒与pET28(a)载体质粒同时进行EcoRⅠ和NotⅠ双酶切,回收并纯化目的片段,并使用T4 DNA连接酶16 ℃连接过夜,随后转化大肠杆菌TOP10感受态细胞,重组质粒经卡那霉素筛选,PCR扩增并进行质粒双酶切验证后,最后挑去阳性克隆子进行测序。将测序正确的重组质粒pET28(a)-[WTBX][STBX]PdHNL1[WTBZ][STBZ]和空载体pET28(a)分别转入表达宿主菌BL21(DE3)中,挑取克隆并使用 T7通用引物进行PCR,筛选阳性菌株。
1.2.4 原核表达及SDS-PAGE电泳检测 将PCR验证正确的重组菌株与对照菌株分别接种于50 mL的LB培养基中,37 ℃振荡培养至D600 nm约为0.8,加入终浓度为1 mmol/L的IPTG于16 ℃进行低温诱导;分别在诱导后的0、3、6、18、24 h时取菌液1 mL,10 000 r/min离心1 min收集菌体,制备SDS-PAGE电泳样品;按照《蛋白质技术手册》配制蛋白胶,并进行SDS-PAGE电泳分析。
2 结果与分析
2.1 提取的RNA质量
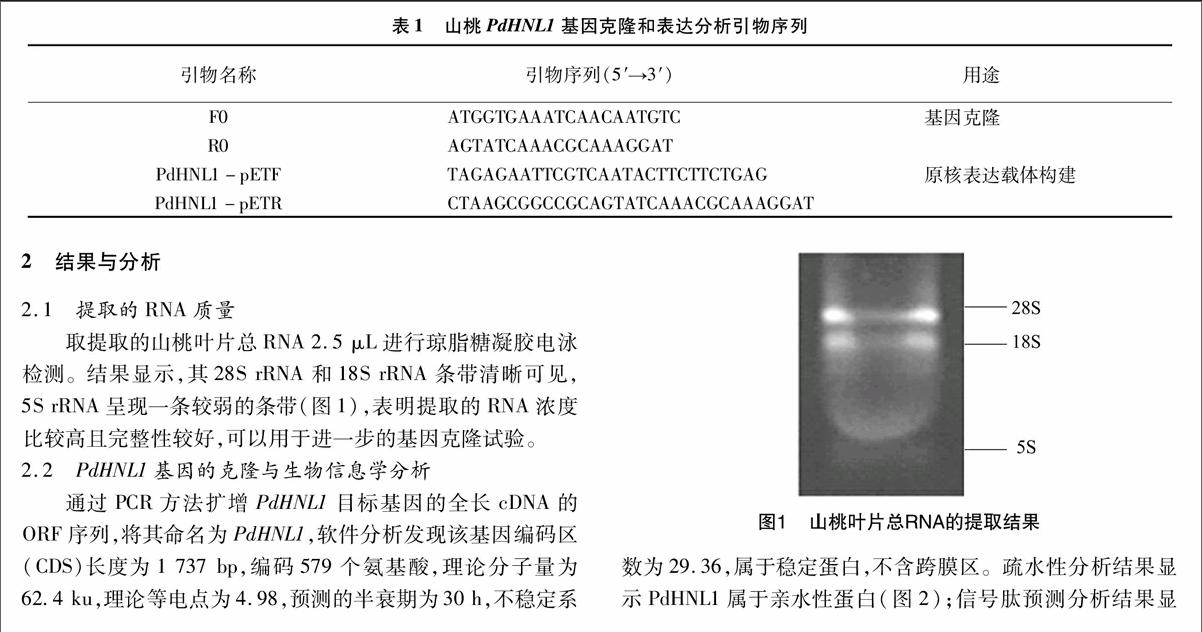
取提取的山桃叶片总RNA 2.5 μL进行琼脂糖凝胶电泳检测。结果显示,其28S rRNA和18S rRNA条带清晰可见,5S rRNA呈现一条较弱的条带(图1),表明提取的RNA浓度比较高且完整性较好,可以用于进一步的基因克隆试验。
2.2 [WTBX][STBX]PdHNL1[WTBZ][STBZ]基因的克隆与生物信息学分析
通过PCR方法扩增[WTBX][STBX]PdHNL1[WTBZ][STBZ]目标基因的全长cDNA的ORF序列,将其命名为[WTBX][STBX]PdHNL1[WTBZ][STBZ],软件分析发现该基因编码区(CDS)长度为1 737 bp,编码579个氨基酸,理论分子量为624 ku,理论等电点为4.98,预测的半衰期为30 h,不稳定系
数为29.36,属于稳定蛋白,不含跨膜区。疏水性分析结果显示PdHNL1属于亲水性蛋白(图2);信号肽预测分析结果显示,该蛋白N端有一段信号肽序列。PdHNL1蛋白序列的二级结构预测结果显示,无规则卷曲是山桃 PdHNL1的主要结构元件;统计结果表明,PdHNL1蛋白的二级结构包含 1672% α-螺旋、17.55%β-折叠和 65.73%无规则卷曲。此外,结构功能分析结果还表明,PdHNL1蛋白有类似葡萄糖甲醇胆碱氧化还原酶的序列特征 (图3)。氨基酸多重序列比对结果显示PdHNL1与黑樱桃PsHNL4同源性较高(图4)。
从GenBank上获得多个植物HNL同源基因编码蛋白的氨基酸序列。利用进化分析MEGA 5.0,采用中邻位相接算法构建系统进化树(图5)。分析结果显示,山桃PdHNL1与同[CM(25]为蔷薇科的黑樱桃PsHNL4进化关系最近;与其他蔷薇科[CM)]植物的HNL蛋白遗传距离也比较接近,同在一个进化分支上。
使用SWISS-MODEL对PdHNL1蛋白的三级结构进行预测,软件自动选择PDB数据库中的梅的 PmHNL isozyme-1蛋白为模板 (同源性为77.65%),对 PdHNL1蛋白进行建模(图6)。结果表明,PdHNL1结构主要为无规则卷曲,这与二级结构预测结果相符。
2.3 [WTBX][STBX]PdHNL1[WTBZ][STBZ]基因的原核表达[JP2]
对PdHNL1进行信号肽预测分析发现,该蛋白N端具有1段信号肽序列。因此,设计去除该信号肽的引物,对PdHNL1的成熟蛋白进行原核表达。将[WTBX][STBX]pMD19T-PdHNL1与pET28(a)[WTBZ][STBZ]质粒使用EcoRⅠ与NotⅠ双酶切(图7-A、图7-B),分别纯化回收后,使用T4 DNA连接酶进行连接,并转化大肠杆菌DH5α感受态细胞。菌液PCR和质粒双酶切试验结果显示,[WTBX][STBX]PdHNL1[WTBZ][STBZ]成功连接到原核表达载体pET28(a) (图7-C、图7-D)。提取重组质粒pET28(a)-[WTBX][STBX]PdHNL1[WTBZ][STBZ]并转入大肠杆菌表达菌株BL21(DE3),菌液PCR检测结果表明获得了用于原核标的重组菌 (图7-E)。
将含有重组质粒pET28(a)-[WTBX][STBX]PdHNL1[WTBZ][STBZ]的阳性菌株和含空载体pET28(a)的对照菌株培养至D600 nm为0.5,随后加入终浓度为0.8 mmol/L IPTG进行诱导表达;分别于IPTG诱导后[CM(25]的0、3、6、18、24[KG*3]h 取样,并制备SDS-PAGE检测样品。[CM)]
SDS-PAGE电泳结果显示,含pET28(a)-[WTBX][STBX]PdHNL1[WTBZ][STBZ]的宿主菌经IPTG诱导后,在62 ku左右有1条清晰的目的条带,且随着时间的延长,该目的条带表达量增加(图8)。这与[WTBX][STBX]PdHNL1[WTBZ][STBZ]预测的分子量相一致,表明山桃 pET28(a)-[WTBX][STBX]PdHNL1[WTBZ][STBZ]重组质粒在大肠杆菌中成功表达。
3 结论与讨论
HNLs是合成纯光学氰醇类化合物的重要的酶催化剂,其在植物体内催化分解氰基糖苷类化合物生成氰化物,从而起到保护植物的作用[19]。自1997年就已经有研究者在植物体内筛选可能的HNL[20];此后,Asano等采用HPLC方法检测[FL)]
了163种产氰植物的HNL活性及其催化产生手性醇氰的能力,发现桃HNL具有R型HNL活性[5]。桃与山桃同属于蔷薇科李亚科桃属,亲缘关系相近。本研究通过筛选桃基因组数据库,PCR扩增得到山桃中克隆得到醇腈酶cDNA序列,生物信息学与分析发现其核苷酸长度、编码蛋白的分子量等与已报道的蔷薇科HNL蛋白相似;进一步的二级结构预测也发现,该蛋白含有大量无规则卷曲和少量 α-螺旋与β-折叠,除N端信号肽外,无明显跨膜螺旋区域,表明同科的HNL具有较高的保守性。
先前的研究表明,R型HNL主要具有氧化还原酶活性,而S型HNL具有α/β水解酶活性。拟南芥HNL则是第1个从非产氰植物中发现的具有α/β水解酶活性的R型HNL[21]。另一方面,HNL蛋白依据是否含有FAD辅基分为FAD-HNLs与非FAD-HNLs。本研究中对包括PdHNL1在内的11种植物的17个HNL蛋白进行聚类分析,发现蔷薇科R型HNL在一个进化分支,且均为FAD-HNLs, 推测蔷薇科
HNL在进化过程中具有高度的保守性。山桃同属蔷薇科且PdHNL1与蔷薇科其他物种的HNL在同一个进化分支上,暗示其可能属于R型HNL。蛋白比对分析结果同样显示,PdHNL1与同属于的蔷薇科的梅、黑樱桃、扁桃的HNL蛋白序列相似度较高,均达75%以上。结构功能分析结果显示,PdHNL1属于葡萄糖甲醇胆碱氧化还原酶家族,且PdHNL1氨基酸序列中具有蔷薇科植物HNL推测的保守氨基酸序列YWHYHGG。此外,拟南芥和亚麻HNL虽属于R型HNL,但与S型醇腈酶亲缘关系较近,归为一个进化分支。
本研究还将[WTBX][STBX]PdHNL1[WTBZ][STBZ]基因连接到原核表达载体pET28(a) 上,通过转化大肠杆菌BL21(DE3)进行融合蛋白的诱导表达,成功获得了重组菌种。上述结果为进一步纯化目标蛋白,并进行PdHNL1催化活性研究奠定了基础。另外,本研究对山桃醇腈酶[WTBX][STBX]PdHNL1[WTBZ][STBZ]基因进行的序列分析与原核表达,还丰富了植物HNL基因的来源,并为推进手性醇腈的酶法制备奠定了基础。
参考文献:
[1]Andexer J N,Langermann J V,Kragl U,et al. How to overcome limitations in biotechnological processes-examples from hydroxynitrile lyase applications[J]. Trends Biotechnol,2009,27 (10):599-607.
[2]Andexer J N,Staunig N,Eggert T,et al. Hydroxynitrile lyases with α/β-hydrolase fold:two enzymes with almost identical 3D structures but opposite enantioselectivities and different reaction mechanisms[J]. Chembiochem,2012,13(13):1932-1939.
[3]邢晓夏,李伟伟,刘均洪. 醇腈酶:生物与化学的结合点[J]. 化工科技,2006,14(2):46-50.
[4]徐军伟,刘均洪. 脂肪酶和醇腈酶在有机合成中的应用[J]. 化工生产与技术,2003,10(4):15-19.
[5]Asano Y,Tamura K,Doi N,et al. Screening for new hydroxynitrilases from plants[J]. Biosci Biotechnol Biochem,2005,69(12):2349-2357.
[6]Krammer B,Rumbold K,Tschemmernegg M,et al. A novel screening assay for hydroxynitrile lyases suitable for high-throughput screening[J]. J Biotechnol,2007,129(1):151-161.
[7]赵冠杰,白锴凯,郑允权,等. 枇杷醇腈酶基因的克隆及结构分析[J]. 福州大学学报:自然科学版,2011,39(6):960-964.
[8]Suelves M,Puigdomènech P. Molecular cloning of the cDNA coding for the (R)-(+)-mandelonitrile lyase of Prunus amygdalus:temporal and spatial expression patterns in flowers and mature seeds[J]. Planta,1998,206 (3):388-393.
[9]Weis R,Poechlauer P,Bona R,et al. Biocatalytic conversion of unnatural substrates by recombinant almond R-HNL isoenzyme 5[J]. Journal of Molecular Catalysis B:Enzymatic,2004,29(1/2/3/4/5/6):211-218.
[10]Hu Z,Poulton J E. Molecular analysis of (R)-(+)-mandelonitrile lyase microheterogeneity in black cherry[J]. Plant Physiol,1999,119(4):1535-1546.
[11]Nanda S,Kato Y,Asano Y. A new (R)-hydroxynitrile lyase from Prunus mume:asymmetric synthesis of cyanohydrins[J]. Tetrahedron,2005,61(46):10908-10916.
[12]Zhao G J,Yang Z Q,Guo Y H. Cloning and expression of hydroxynitrile lyase gene from Eriobotrya japonica in Pichia pastoris[J]. J Biosci Bioeng,2011,112(4):321-325.
[13]Ueatrongchit T,Tamura K,Ohmiya T,et al. Hydroxynitrile lyase from passiflora edulis:Purification,characteristics and application in asymmetric synthesis of (R)-mandelonitrile[J]. Enzyme and Microbial Technology,2010,46(6):456-465.
[14]Narayanan N N,Ihemere U,Ellery C,et al. Overexpression of hydroxynitrile lyase in cassava roots elevates protein and free amino acids while reducing residual cyanogen levels[J]. PLoS One,2011,6(7):e21996.[HJ1.65mm]
[15]Hasslacher M,Schall M,Hayn M,et al. Molecular cloning of the full-length cDNA of (S)-hydroxynitrile lyase from Hevea brasiliensis:functional expression in Escherichia coli and Saccharomyces cerevisiae and identification of an active site residue[J]. J Biol Chem,1996,271(10):5884-5891.
[16]Kuroki G W,Conn E E. Mandelonitrile lyase from Ximenia americana L.:stereospecificity and lack of flavin prosthetic group[J]. Proc Natl Acad Sci USA,1989,86(18):6978-6981.
[17]Wajant H,Mundry K W,Pfizenmaier K. Molecular cloning of hydroxynitrile lyase from Sorghum bicolor (L.) homologies to serine carboxypeptidases[J]. Plant Mol Biol,1994,26(2):735-746.
[18]International Peach Genome Initiative,Verde I,Abbott A G,et al. The high-quality draft genome of peach(Prunus persica) identifies unique patterns of genetic diversity,domestication and genome evolution[J]. Nat Genet,2013,45(5):487-494.
[19]Krammer B,Rumbold K,Tschemmernegg M,et al. A novel screening assay for hydroxynitrile lyases suitable for high-throughput screening[J]. J Biotechnol,2007,129(1):151-161.
[20]Hickel A,Heinrich G,Schwab H,et al. Screening for hydroxynitrile lyasea in plants[J]. Biotechnology Techniques,1997,11(1):55-58.
[21]Andexer J,von Langermann J,Mell A,et al. An R-selective hydroxynitrile lyase from Arabidopsis thaliana with an α/β-hydrolase fold[J]. Angewandte Chemie International Edition,2007,46(45):8679-8681.
猜你喜欢
动漫界·幼教365(小班)(2021年6期)2021-07-21
山东农业科学(2016年11期)2016-12-17
山东农业科学(2014年1期)2015-03-09
中国蜂业(2013年11期)2013-01-23
